
Abstract
Purpose
To study the effect of a countermovement on the lower limb force–velocity (F–v) mechanical profile and to experimentally test the influence of F–v mechanical profile on countermovement jump (CMJ) performance, independently from the effect of maximal power output (P max).
Methods
Fifty-four high-level sprinters and jumpers performed vertical maximal CMJ and squat jump (SJ) against five to eight additional loads ranging from 17 to 87 kg. Vertical ground reaction force data were recorded (1,000 Hz) and used to compute center of mass vertical displacement. For each condition, mean force, velocity, and power output were determined over the entire push-off phase of the best trial, and used to determine individual linear F–v relationships and P max. From a previously validated biomechanical model, the optimal F–v profile maximizing jumping performance was determined for each subject and used to compute the individual mechanical F–v imbalance (Fv IMB) as the difference between actual and optimal F–v profiles.
Results
A multiple regression analysis clearly showed (r 2 = 0.952, P < 0.001, SEE 0.011 m) that P max, Fv IMB and lower limb extension range (h PO) explained a significant part of the interindividual differences in CMJ performance (P < 0.001) with positive regression coefficients for P max and h PO and a negative one for Fv IMB.
Conclusion
Compared to SJ, F–v relationships were shifted to the right in CMJ, with higher P max, maximal theoretical force and velocity (+35.8, 20.6 and 13.3 %, respectively). As in SJ, CMJ performance depends on Fv IMB, independently from the effect of P max, with the existence of an individual optimal F–v profile (Fv IMB having an even larger influence in CMJ).
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Jumping ability is of great interest since it is a key factor in numerous sport activities, but also because it represents the typical example of ballistic movements aiming at accelerating its own body mass the most as possible over one leg extension. Jumping performance is, thus, frequently used to monitor both physical fitness, evaluate the maximal mechanical capabilities of the lower limb neuromuscular system (henceforth referred to as “lower limbs”) and/or set strength and conditioning programs (Cormie et al. 2007). Among the various vertical jumps used as indirect measurements of strength and anaerobic power of the lower limb extensor muscles (Cormie et al. 2011; Cronin and Sleivert 2005), the most frequently used, reliable, valid and contrasted ones have been squat jump (SJ) and countermovement jump (CMJ) (Bobbert et al. 1996; Klavora 2000). CMJ remains the most commonly used in sports training and testing, but also the most appropriate test with regard to specific populations such as athletes, most especially sprinters (Canavan and Vescovi 2004; Markström and Olsson 2013). Indeed, in many sport activities (e.g., sprint running, jumps, changes of direction), dynamic movements involving successive eccentric and concentric muscle actions (closer to CMJ exercise conditions) happen to occur much more frequently than just a sole concentric action (which is the essence of SJ). Furthermore, CMJ height has been regularly used to assess the effectiveness of the stretch–shortening cycle during a given athletic performance (Hoffman et al. 2002, 2005). For instance, a strong and consistent correlation has been observed between CMJ height and sprint ability (Cronin and Hansen 2005).
Performance in CMJ is 5–10 % higher than in SJ (Bobbert and Casius 2005). In ballistic movements such as a jumping, it has been reported that performance depends on the mechanical impulse performed in the movement direction (Winter 2005), and developing a high mechanical impulse during a lower limb push-off was assumed to depend on power output capabilities of lower limbs (Samozino et al. 2008; Vandewalle et al. 1987; Yamauchi and Ishii 2007). Consequently, the countermovement in CMJ likely allows athletes to increase the mechanical impulse or work developed during push-off by enhancing the lower limbs power output capabilities (Bosco et al. 1983). This was first explained by the effects of elastic energy storage and reuse, but also by myoelectrical potentiation (Bosco et al. 1982), and more recently by the fact that muscles’ active state develops during the preparatory downward movement, which induces a higher force at the beginning of the push-off, and thus a higher mechanical work or impulse over the push-off phase (Bobbert and Casius 2005). However, since power output directly depends on both force and velocity capabilities of the lower limbs, it is likely that the countermovement affects lower limbs force and/or velocity capabilities. The latter have been extensively described during dynamic conditions using the inverse linear force–velocity (F–v) and second-degree polynomial power–velocity (P–v) relationships during various types of multi-joint concentric extension movements (Bobbert 2012; Samozino et al. 2012, 2013; McMaster et al. 2013). These relationships describe the changes in external force generation and power output with increasing movement velocity, and may be summarized in two variables: the maximal power output (P max, apex of the P–v relationship) the lower limbs can produce over one extension, and the F–v profile (i.e., slope of the linear F–v relationship) that represents the balance between the external force and the velocity maximal capabilities (Samozino et al. 2012, 2013). These two mechanical properties are independent, since two individuals could present the same P max with different F–v profiles. The F–v characteristic of the lower limb muscles is a key factor in physical performance assessment, and an appropriate determination of F–v relationship appears therefore to be essential to quantify the mechanical properties of the lower limbs neuromuscular system in vivo (Samozino et al. 2010, 2012; Yamauchi and Ishii 2007).
Comparing P max and F–v profile between CMJ and SJ could provide new macroscopic insights into the mechanical advantages of using a countermovement during ballistic tasks, notably in jumping. To our knowledge, the comparisons of the mechanical lower limbs capabilities between exercises—involving a countermovement (CMJ) or not (SJ)—have not been specifically studied yet. Although Bosco and Komi (1979) reported F–v and P–v relationships in SJ and CMJ (Fig. 3 in Bosco and Komi 1979), they did not further analyze this comparison between CMJ and SJ, notably with respect to P max F–v profile. Also, very recently, McMaster et al. (2013) compared different rugby players’ profiles during ballistic and maximum strength upper body exercises, but did not focus on the effect of countermovement on the F–v profile.
The major interest of analyzing the F–v profile in explosive performances has recently been shown using a macroscopic biomechanical model putting forward its effect on SJ performances, independently from the large effect of P max, associated to the existence of an individual optimal F–v profile maximizing jumping performance (Samozino et al. 2008, 2010, 2012). The aforementioned optimal F–v profile represents the best balance between force and velocity qualities to maximize ballistic push-offs, such as jumps. It could therefore be interesting not only for scientists working on muscle function during maximal efforts, but also by coaches for training purposes (Samozino et al. 2012). Very recently, experimental findings have clearly supported that SJ performance depends on both a high P max and a low F–v imbalance (difference between the athlete F–v profile and his/her optimal one to maximize jumping ability) (Samozino et al. 2013). Since it is reasonable to assume that the countermovement induces changes in F–v relationship, experimental data are required to test the influence of F–v mechanical profile on CMJ performance with the associated existence of an individual optimal F–v profile, as theoretically predicted by the above-mentioned biomechanical model (Samozino et al. 2010, 2012).
In this context, this study aimed at (1) analyzing the effect of countermovement on lower limb F–v relationships and power output capabilities by comparing SJ and CMJ modalities, (2) bringing experimental support to the theoretical influence of F–v imbalance on CMJ performance, independently from the largely known effect of P max, and (3) quantifying the respective contribution of P max, F–v imbalance and h PO in CMJ performance in high-level track and field athletes.
Methods
Subjects and study design
Fifty-four trained male athletes (age 23.1 ± 4.4 years, body mass 77.9 ± 6.0 kg, height 1.80 ± 0.06 m) gave their written informed consent to participate in this study, which was approved by the local ethical committee of the University of Pablo de Olavide (Seville, Spain) and performed in agreement with the Declaration of Helsinki. All subjects were Spanish national/international level sprinters or jumpers, and attended two 1.5-h measurement sessions (CMJ and SJ) in a randomized order, at 72-h intervals (no physical activity was allowed on the days preceding the 2 sessions). The two sessions involved a complete and appropriate warm-up, followed by maximal SJs or CMJs in different loading conditions (with and without additional loads) to determine individual F–v relationships in SJ or CMJ, respectively. Individual CMJ and SJ performances were obtained from maximal jumps without loads.
Testing procedures
To determine individual F–v relationships, each subject performed vertical maximal jumps without loads and against five to eight extra loads ranging from 17 to 87 kg in a randomized order. The test was performed on a Smith machine (Multipower Fitness Line, Peroga, Spain) that allows a smooth vertical displacement of the bar along a fixed vertical path. Before each jump, participants were instructed to stand up straight and still on the center of the force plate with their hands on the hips for unloaded conditions and on the bar for loaded jumps, this hand position remaining the same during the entire movement. From this position, participants initiated a downward movement until a crouching position with a knee angle of about 90°, followed by a jump for maximal height (immediately for CMJ or after 2 s for SJ). For SJ, countermovement was verbally forbidden and carefully checked after each trial using force–time curves. Two valid trials were performed with each load with 2-min recovery between trials and 4–5 min between loads condition.
Equipment and data acquisition
All CMJ and SJ trials were performed on a standard force plate (Bertec, Type 4060-15, Bertec Corporation, Columbus, OH, USA) that sampled (1,000 Hz) vertical ground reaction force. These devices were interfaced with an analog to digital converter MP100.2.0 (Biopac Systems Inc., Santa Barbara, CA, USA) connected to a PC. A customized software (Isonet, Madrid, Spain) provided real-time collection, synchronization and visualization of data.
Mean force (\( \overline{F} \)), velocity (\( \overline{v} \)), and power (\( \overline{P} \)) output data from the best trial of each condition were determined from instantaneous values recorded over the entire push-off phase. The vertical velocity of the body center of mass was obtained from the integration of the vertical acceleration signal obtained from force plate measurement. The push-off began when the velocity signal increased and ended when the force signal at takeoff fell to zero. As previously suggested (Bosco et al. 1995; Rahmani et al. 2001; Samozino et al. 2012; Yamauchi and Ishii 2007), F–v relationships were determined by least squares linear regressions. F–v curves were extrapolated to obtain F 0 (then normalized to body mass, in N kg−1) and v 0 (in m s−1), which, respectively, correspond to the intercepts of the F–v curve with the force and velocity axis. The F–v profile was computed as the slope of the F–v linear relationship (S Fv, in N s kg−1 m−1) (Samozino et al. 2012). Values of P max (normalized to body mass, in W kg−1) were determined as P max = F 0 · v 0/4 (Samozino et al. 2012; Vandewalle et al. 1987). In addition, the vertical distance covered by the CM during push-off (h PO, in m), that corresponds to the extension range of lower limbs, was determined from integration of the velocity signal (Samozino et al. 2012). The theoretical optimal S Fv (S Fvopt, normalized to body mass, in N s kg−1 m−1) maximizing vertical jumping performance was computed for each subject as proposed by Samozino et al. (2012). The F–v imbalance (Fv IMB, in %) represents the magnitude of the unfavorable balance between force and velocity qualities to maximize jump height, and was individually computed as recently proposed (Samozino et al. 2013):
An individual Fv IMB value at 0 % indicates an optimal profile (perfect balance between force and velocity qualities to maximize jump height), the higher Fv IMB, the higher unfavorable F–v balance to maximize jump height, be it a profile too much oriented towards force or velocity capabilities.
The hypothetical maximal jump height each subject could reach, should he present an optimal F–v profile (h max, in m), was computed from his actual P max, h PO and S Fvopt values, using the following equation derived from previously published equations (Samozino et al. 2010, 2012):
The theoretical loss of performance due to the F–v imbalance was then computed for each subject as the difference between actual unloaded jump height and h max (in percentage of h max).
Statistical analysis
All data are presented as mean ± SD. Normality and homogeneity of variance were checked before analyses. The degree of linear relationship between variables was examined using Pearson’s product moment correlation, and h PO, P max, F 0, S Fv and Fv IMB values were compared between SJ and CMJ conditions using paired t tests. To test the independent effect of P max and Fv IMB on CMJ performance, a stepwise forward multiple regression analysis was performed with maximal unloaded CMJ height as the dependent variable, and Fv IMB, P max and h PO as independent variables. This multiple regression model was based on the previously proposed theoretical approach (Samozino et al. 2012). The F values for the inclusion and exclusion of variables were set at 4.0. Besides, stepwise the forward multiple regression analysis allowed the weighting of the independent variable effects on the CMJ performance. The effect of F–v profile was tested in regression analysis through Fv IMB, since the relationship between F–v profile and performance is curvilinear while the one between performance and Fv IMB is supposed to be purely decreasing (Samozino et al. 2013). For all statistical analyses, a P value of 0.05 was accepted as the level of significance.
Results
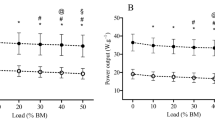
Individual F–v relationships were well fitted by linear regressions for SJ and CMJ (r 2 = 0.9–0.99, P < 0.012, typical subject in Fig. 1). Mean ± SD values of variables associated to SJ and CMJ F–v relationships are presented in Table 1 for all subjects. All these values were significantly different between SJ and CMJ (P < 0.001), except for h PO, S Fv, and Fv IMB (P > 0.05). An important finding was that average differences of 20.6 % in F 0 and 13.3 % in v 0 (both significant) were observed between SJ and CMJ.

Individual F–v relationships for a typical subject for SJ (filled circles) and CMJ (open circles)
Simple correlation analyses showed that CMJ performance was significantly correlated to P max and h PO, but not to S Fv and Fv IMB (Table 2). No simple correlation was found between the three independent variables (P max, Fv IMB and h PO). The stepwise forward multiple regression analysis indicated that, when considered as a whole, the three latter predicting variables accounted for a significant amount of CMJ performance variability (P < 0.001, Table 3), with a high quality of adjustment (r 2 = 0.952, P < 0.001) and minimal error (SEE 0.011 m) (Table 3; Fig. 2). The interindividual variability in CMJ performance was explained for 57.3, 28.5 and 9.5 % by variability in P max, h PO and Fv IMB, respectively. The loss of CMJ performance due to individual F–v imbalance was 10.83 ± 4.93 % (from 1.14 to 21.83 %). Figure 3 presents the actual CMJ jump height reached by each subject (h expressed relatively to h max) according to their F–v profile (S Fv expressed relatively to their personal S Fvopt), as well as the corresponding theoretical changes predicted by the model (Eq. (2) with h PO, h and P max values arbitrarily set to the average values for the group) (Samozino et al. 2012).

Correlation between model-predicted and measured jump heights. Each point represents a subject. The solid line represents the identity line

Actual jump height reached in unloaded CMJ condition (expressed for each individual relatively to the theoretical h max) as a function of the F–v profile (S Fv expressed for each individual relatively to his personal S Fvopt). The solid line represents the theoretical changes predicted by the model [equation (5) in Samozino et al. (2013) with h PO and P max values arbitrarily set to the average values for the group]
Discussion
The novel findings of this study were: (1) F–v relationships in CMJ are linear as previously shown in SJ, with P max, F 0 and v 0 values significantly higher in CMJ compared to SJ, (2) the shift in F 0 between SJ and CMJ is about twice higher than the shift in v 0, and (3) maximal CMJ performance depends not only on P max, but also on the F–v imbalance, i.e., the difference between the actual and optimal F–v profiles, as recently shown for SJ (Samozino et al. 2013).
SJ and CMJ are different from a technical and a biomechanical point of view, which explains that CMJ height is consistently higher than SJ height (Bobbert and Casius 2005). Our results showed a 15.4 % difference between SJ and CMJ performances, which is similar or higher to what has been reported in sprinters (e.g., Markström and Olsson 2013) (7–9 %), well-trained males (Bosco and Komi 1979) (16 %) or trained soccer players (Comfort et al. 2014) (4.5 %). F 0 and v 0 measured here in SJ are also consistent with those previously reported (Bosco et al. 1995; Rahmani et al. 2001, 2004; Samozino et al. 2012, 2013; Yamauchi and Ishii 2007), while S Fv values were slightly lower than those reported by Samozino et al. (2012, 2013) in inclined maximal lower limb extension and SJ.
The comparison of lower limbs power, force and velocity capabilities between SJ and CMJ has never been specifically studied, and the present results showed a shift to the right of the F–v relationship between SJ and CMJ (Table 1; Fig. 1). Power output capabilities increased in CMJ compared to SJ, which likely explains the improvement in jump height. Significant differences of 20.6 % in F 0 and 13.3 % in v 0 were observed on average between SJ and CMJ. This difference in F 0 is similar to that observed by McMaster et al. (2013) in countermovement versus concentric-only exercises for the upper body. Investigating lower limb mechanical capabilities in CMJ certainly is of interest, since this jumping modality involves mechanical and neuromuscular mechanisms participating to a more natural jumping movement (i.e., including a countermovement phase) with a higher specificity when assessing athletes and has also been clearly related to the performance in many sports activities such as soccer, rugby or athletics (e.g., Comfort et al. 2014; Wisloff et al. 2004).
Different explanations have been proposed for the higher CMJ height compared to SJ (Bobbert and Casius 2005; Bobbert et al. 1996), and more recently, the fact that muscles’ active state develops during the preparatory downward movement, which involves a higher muscle active state at the beginning of push-off, and in turn a greater force (Bobbert and Casius 2005). The higher CMJ height would, thus, be due to the greater force and work that the lower limbs extensor muscles can produce over the first part of the upward motion in CMJ (Bobbert and Casius 2005). Our results suggest that this greater active state at the beginning of the jump affects more the force capabilities of the neuromuscular system (F 0) than the velocity qualities (v 0), i.e., conditions with high extra loads (high force/low velocity) than conditions without load (high velocity/low force).
In the macroscopic biomechanical model used here (Samozino et al. 2010, 2012), F–v relationships and the associated parameters describe the final result of the different mechanisms involved in limbs extension, which are a complex integration of numerous individual muscle mechanical properties (e.g., intrinsic F–v and length–tension relationship, rate of force development), some morphological factors (e.g., cross-sectional area, fascicle length, pennation angle, tendon properties, anatomical joint configuration), neural mechanisms (e.g., motor unit recruitment, firing frequency, motor unit synchronization, intermuscular coordination) and segmental dynamics (Bobbert 2012; Cormie et al. 2011). The twice higher differences in F 0 compared to v 0 between CMJ and SJ could result from a higher rate of force development (Vanezis and Lees 2005) or an optimized motor unit recruitment (Maffiuletti et al. 2002). These are jumping performance factors that directly affect F 0, and it is likely that these factors play a more important role in CMJ than they do in SJ, notably through the higher active state at the beginning of the push-off phase. Contrastingly, differences in v 0 between CMJ and SJ are of lower magnitude than for F 0 because the neural and/or mechanical mechanisms involved in CMJ are more pronounced in high force/low velocity movement than in low force/high velocity movement.
Assessing individual F–v profiles makes possible to identify the optimal balance between force and velocity capabilities that maximizes, for each individual, jumping performance (Samozino et al. 2013). Therefore, computing each individual’s optimal S Fv, and then determining the F–v imbalance (Fv IMB, in %), i.e., an unfavorable balance between force and velocity could be very interesting to better optimize training in explosive movements, notably in CMJ. Simple correlations (Table 2) showed that jumping performance was highly and significantly correlated to P max, with a similar magnitude as previously reported (e.g., Samozino et al. 2013; Vandewalle et al. 1987; Yamauchi and Ishii 2007, r values ranging from 0.76 in CMJ to 0.87 in SJ), but not directly to S Fv or Fv IMB, which is in line with what Samozino et al. (2013) found in SJ. Our results clearly confirm that CMJ performance mainly depends on P max (Comfort et al. 2014; Markström and Olsson 2013). The absence of an experimental correlation between CMJ performance and S Fv was expected, knowing the theoretical curvilinear relationship between these two variables (Samozino et al. 2012). By testing the effect of each variable (notably P max and Fv IMB) independently from the other, the multiple regression analysis showed that interindividual variability in CMJ performance was partly explained by Fv IMB (Table 3): for a given P max, the higher Fv IMB, the lower the CMJ performance. The present data showed that differences in CMJ performance across the track and field athletes tested were explained at about 10 % by Fv IMB, which is relatively meaningful when high-level performances are considered. As expected, P max accounted for the main part of CMJ performance variability (57.3 %), which is an important contribution to performance, yet lower than what could have been expected considering previous results associating directly jumping performance to maximal power capabilities (Canavan and Vescovi 2004; Frost et al. 2010; Cormie et al. 2011). In addition, a relatively large part (28.5 %) of between-subject variability in CMJ height was due to differences in h PO, characterizing the usual or optimal individual lower limb extension range, which directly depends on athlete’s morphological characteristics and on the task specificities, and which cannot be changed by training. This indirectly increases the importance of Fv IMB in the part of CMJ performance that can be improved by training. It is worth noting that this study is focused on sprinters and jumpers, i.e., subjects with mostly F–v imbalance toward velocity qualities since they were used to perform ballistic horizontal movements (e.g., horizontal jumps, sprints) in their sport activities, i.e., movements against low loads at high velocity (faster than a vertical jump). Their training history made them develop more velocity than force qualities, which tends to be the opposite, for instance, of rugby players who rather showed F–v imbalances towards force qualities (Samozino et al. 2013). The relatively high homogeneity in the F–v profiles of present highly trained subjects (few of them presenting F–v profiles too much oriented toward force qualities) inevitably induces an underestimation of the statistical contribution of Fv IMB in CMJ performance.
The negative effect of Fv IMB on CMJ performance was computed through the individual loss of performance due to Fv IMB ranging from 0 % for subjects presenting an optimal profile to ~20 % for the most extreme values (Fig. 3), which echoes both the theoretical simulations previously reported (Samozino et al. 2012, 2013) and the experimental data obtained in SJ for high-level sprinters, soccer and rugby players (Samozino et al. 2013). This result also supports the importance of considering Fv IMB, and in turn the optimal F–v profile when aiming to improve jumping performance, and more largely success in ballistic movements.
The main limit of the biomechanical approach used here might be the macroscopic level from which the multi-segmental neuromuscular system is considered, inducing (1) the description of its mechanical external capabilities by the empirically determined F–v relationships, and (2) the application of principles of dynamics to a whole body considered as a system (these points have been discussed in details in Samozino et al. 2010, 2012) and very recently showed and validated for SJ by Samozino et al. (2013). The bias induced by the simplifications and approximations associated with this approach was shown to be low (< 6 %) and trivial, which supported its validity (Samozino et al. 2012). This validity is also strengthened, as shown in Fig. 3, by the high agreement between measured performances and theoretical changes predicted by the model. The F–v linear model used to characterize the dynamic external capabilities of the lower limb neuromuscular system during multi-joint lower limb extensions has been well supported and discussed (Bobbert 2012; Bosco et al. 1995; Rahmani et al. 2001; Samozino et al. 2012; Yamauchi and Ishii 2007).
Practical applications
Since jumping performance represents the typical example of ballistic movements, and such movements (i.e., jumping, start sprinting, changing of direction, etc.,) are some of the keys of performance in numerous sport activities, an optimization of the F–v profile by reducing the “FV imbalance” through strength and conditioning could be useful. Indeed, athletes would benefit from monitoring each individual F–v profile and set their training regimen accordingly on an individual basis. Furthermore, should they increase their ballistic movement ability, this could play a positive role in their overall performance, whatever the sport considered (soccer, rugby, basketball, etc.,). Thus, the difference between a subject’s theoretically optimal and actual F–v profiles is computed through the Fv IMB, the significant implication of Fv IMB in CMJ performance experimentally supports the existence of an optimal F–v profile, specific to each individual, that maximizes CMJ performance for a given P max. The main practical consequence of this result is that, to improve the CMJ performance (and indirectly partly functional performance in many sports involving dynamic contractions and stretch–shortening cycle actions), coaches and athletes would greatly benefit from going further than only seeking to increase maximal power, and monitoring each individual F–v profile to set their training regimen accordingly. Finally, all the computations and findings presented here are based on anthropometrical and biomechanical variables pretty easy to collect in a typical training context: body and additional load mass, lower limb extension range, and jump height.
Conclusions
The linear F–v relationship in CMJ was shifted to the right in comparison to SJ with a larger shift in F 0 than in v 0. The corroborated increase in CMJ maximal height compared to SJ is due to a higher P max and more especially to a shift in the F–v profile, which is more marked on the force than on the velocity axis. Furthermore, even if P max remains the main determinant of CMJ performance, the latter depends also on the F–v profile of lower limbs, which characterizes the ratio between their maximal force and their maximal velocity capabilities. Ceteris paribus, a F–v imbalance is associated to a lower CMJ performance, which extends recent experimental conclusions drawn for pure concentric SJ (Samozino et al. 2013) to an exercise (CMJ) that is more frequently used in sports training and testing.
Abbreviations
- SJ:
-
Squat jump
- CMJ:
-
Countermovement jump
- F–v :
-
Force–velocity
- P–v :
-
Power–velocity
- P max :
-
Maximal power output
- \( \overline{F} \) :
-
Mean force
- \( \overline{v} \) :
-
Mean velocity
- \( \overline{P} \) :
-
Mean power
- h PO :
-
Vertical distance covered by the CM during push-off
- Fv IMB :
-
F–v imbalance
- S Fv :
-
F–v profile
- S Fvopt:
-
Optimal F–v profile
References
Bobbert MF (2012) Why is the force–velocity relationship in leg press tasks quasi-linear rather than hyperbolic? J Appl Physiol. doi:10.1152/japplphysiol.00787.2011
Bobbert MF, Casius LJ (2005) Is the effect of a countermovement on jump height due to active state development? Med Sci Sports Exerc 37(3):440–446
Bobbert MF, Gerritsen KG, Litjens MC, van Soest AJ (1996) Why is countermovement jump height greater than squat jump height? Med Sci Sports Exerc 28(11):1402–1412
Bosco C, Komi PV (1979) Potentiation of the mechanical behavior of the human skeletal muscle through prestretching. Acta Physiol Scand 106(4):467–472
Bosco C, Viitasalo JT, Komi PV, Luhtanen P (1982) Combined effect of elastic energy and myoelectrical potentiation during stretch-shortening cycle exercise. Acta Physiol Scand 114(4):557–565
Bosco C, Luhtanen P, Komi PV (1983) A simple method for measurement of mechanical power in jumping. Eur J Appl Physiol Occup Physiol 50(2):273–282
Bosco C, Belli A, Astrua M, Tihanyi J, Pozzo R, Kellis S, Tsarpela O, Foti C, Manno R, Tranquilli C (1995) A dynamometer for evaluation of dynamic muscle work. Eur J Appl Physiol Occup Physiol 70(5):379–386
Canavan PK, Vescovi JD (2004) Evaluation of power prediction equations: peak vertical jumping power in women. Med Sci Sports Exerc 36(9):1589–1593
Comfort P, Stewart A, Bloom L, Clarkson B (2014) Relationships between strength, sprint and jump performance in well trained youth soccer players. J Strength Cond Res 28(1):173–177. doi:10.1519/JSC.0b013e318291b8c7
Cormie P, McCaulley GO, Triplett NT, McBride JM (2007) Optimal loading for maximal power output during lower-body resistance exercises. Med Sci Sports Exerc 39(2):340–349. doi:10.1249/01.mss.0000246993.71599.bf
Cormie P, McGuigan MR, Newton RU (2011) Developing maximal neuromuscular power: part 1—biological basis of maximal power production. Sports Med 41(1):17–38
Cronin J, Hansen KT (2005) Strength and power predictors of sports speed. J Strength Cond Res 19(2):349–357
Cronin J, Sleivert G (2005) Challenges in understanding the influence of maximal power training on improving athletic performance. Sports Med 35(3):213–234
Frost DM, Cronin J, Newton RU (2010) A biomechanical evaluation of resistance: fundamental concepts for training and sports performance. Sports Med 40(4):303–326
Hoffman JR, Maresh CM, Newton RU, Rubin MR, French DN, Volek JS, Sutherland J, Robertson M, Gomez AL, Ratamess NA, Kang J, Kraemer WJ (2002) Performance, biochemical, and endocrine changes during a competitive football game. Med Sci Sports Exerc 34(11):1845–1853. doi:10.1249/01.MSS.0000035373.26840.F8
Hoffman JR, Ratamess NA, Cooper JJ, Kang J, Chilakos A, Faigenbaum AD (2005) Comparison of loaded and unloaded jump squat training on strength/power performance in college football players. J Strength Cond Res 19(4):810–815
Klavora P (2000) Vertical-jump tests: a critical review. Strength Cond 22:70–75
Maffiuletti NA, Dugnani S, Folz M, Di Pierno E, Mauro F (2002) Effect of combined electrostimulation and plyometric training on vertical jump height. Med Sci Sports Exerc 34(10):1638–1644
Markström JL, Olsson CJ (2013) Countermovement jump peak force relative to body weight and jump height as predictors for sprint running performances: (in)homogeneity of track and field athletes? J Strength Cond Res 27(4):944–953
McMaster DT, Gill N, Cronin J, McGuigan M (2013) Force–velocity–power assessment in semi-professional rugby union players. J Strength Cond Res. doi:10.1519/JSC.0b013e3182a1da46
Rahmani A, Viale F, Dalleau G, Lacour JR (2001) Force/velocity and power/velocity relationships in squat exercise. Eur J Appl Physiol 84(3):227–232
Rahmani A, Locatelli E, Lacour JR (2004) Differences in morphology and force/velocity relationship between Senegalese and Italian sprinters. Eur J Appl Physiol 91(4):399–405
Samozino P, Morin JB, Hintzy F, Belli A (2008) A simple method for measuring force, velocity and power output during squat jump. J Biomech 41(14):2940–2945
Samozino P, Morin JB, Hintzy F, Belli A (2010) Jumping ability: a theoretical integrative approach. J Theor Biol 264(1):11–18. doi:10.1016/j.jtbi.2010.01.021
Samozino P, Rejc E, Di Prampero PE, Belli A, Morin JB (2012) Optimal force–velocity profile in ballistic movements—altius: citius or fortius? Med Sci Sports Exerc 44(2):313–322. doi:10.1249/MSS.0b013e31822d757a
Samozino P, Edouard P, Sangnier S, Brughelli M, Gimenez P, Morin JB (2013) Force–velocity profile: imbalance determination and effect on lower limb ballistic performance. Int J Sports Med. doi:10.1055/s-0033-1354382
Vandewalle H, Peres G, Heller J, Panel J, Monod H (1987) Force–velocity relationship and maximal power on a cycle ergometer. Correlation with the height of a vertical jump. Eur J Appl Physiol Occup Physiol 56(6):650–656
Vanezis A, Lees A (2005) A biomechanical analysis of good and poor performers of the vertical jump. Ergonomics 48(11–14):1594–1603
Winter EM (2005) Jumping: power or impulse. Med Sci Sports Exerc 37(3):523–524
Wisloff U, Castagna C, Helgerud J, Jones R, Hoff J (2004) Strong correlation of maximal squat strength with sprint performance and vertical jump height in elite soccer players. Br J Sports Med 38(3):285–288
Yamauchi J, Ishii N (2007) Relations between force–velocity characteristics of the knee-hip extension movement and vertical jump performance. J Strength Cond Res 21(3):703–709
Acknowledgments
The authors thank coaches and athletes for their “explosive” and “powerful” implication in the protocol. The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Michael Lindinger.
Rights and permissions
About this article

Cite this article
Jiménez-Reyes, P., Samozino, P., Cuadrado-Peñafiel, V. et al. Effect of countermovement on power–force–velocity profile. Eur J Appl Physiol 114, 2281–2288 (2014). https://doi.org/10.1007/s00421-014-2947-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-014-2947-1