
Abstract
This study aimed to characterise both the \( \dot{V}{\text{O}}_{2} \) kinetics within constant heavy-intensity swimming exercise, and to assess the relationships between \( \dot{V}{\text{O}}_{2} \) kinetics and other parameters of aerobic fitness, in well-trained swimmers. On separate days, 21 male swimmers completed: (1) an incremental swimming test to determine their maximal oxygen uptake \( (\dot{V}{\text{O}}_{2\max } ) \), first ventilatory threshold (VT), and the velocity associated with \( \dot{V}{\text{O}}_{2\max } \) \( (v\dot{V}{\text{O}}_{2\max } ) \) and (2) two square-wave transitions from rest to heavy-intensity exercise, to determine their \( \dot{V}{\text{O}}_{2} \) kinetics. All the tests involved breath-by-breath analysis of freestyle swimming using a swimming snorkel. \( \dot{V}{\text{O}}_{2} \) kinetics was modelled with two exponential functions. The mean values for the incremental test were 56.0 ± 6.0 ml min−1 kg−1, 1.45 ± 0.08 m s−1; and 42.1 ± 5.7 ml min−1 kg−1 for \( \dot{V}{\text{O}}_{2\max } \), \( v\dot{V}{\text{O}}_{2\max } \) and VT, respectively. For the square-wave transition, the time constant of the primary phase (τp) averaged 17.3 ± 5.4 s and the relevant slow component (A′sc) averaged 4.8 ± 2.9 ml min−1 kg−1 [representing 8.9% of the end-exercise \( \dot{V}{\text{O}}_{2} \) (%A′sc)]. τp was correlated with \( v\dot{V}{\text{O}}_{2\max } \) (r = −0.55, P = 0.01), but not with either \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) (r = 0.05, ns) or VT (r = 0.14, ns). The %A′sc did not correlate with either \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) (r = −0.14, ns) or \( v\dot{V}{\text{O}}_{2\max } \) (r = 0.06, ns), but was inversely related with VT (r = −0.61, P < 0.01). This study was the first to describe the \( \dot{V}{\text{O}}_{2}\) kinetics in heavy-intensity swimming using specific swimming exercise and appropriate methods. As has been demonstrated in cycling, faster \( \dot{V}{\text{O}}_{2} \) kinetics allow higher aerobic power outputs to be attained. The slow component seems to be reduced in swimmers with higher ventilatory thresholds.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Several studies (Carter et al. 2000; Borrani et al. 2001; Ingham et al. 2007; Whipp and Wasserman 1972; Whipp et al. 1982) have described the kinetics of the oxygen uptake \( (\dot{V}{\text{O}}_{2} ) \) response within constant load cycling, running or rowing exercise. When such exercise is performed at heavy intensities [i.e. above the ventilatory threshold (VT)], the attainment of a steady state \( \dot{V}{\text{O}}_{2} \) is delayed by a supplemental rise in \( \dot{V}{\text{O}}_{2} \) or “slow component” (Whipp and Wasserman 1972; Whipp et al. 2002). It has been suggested that this slow component is primarily linked to the progressive recruitment of fast-twitch fibres in the exercising muscle (Barstow et al. 1996; Poole et al. 1991, 2008; Vanhatalo et al. 2011).
The time constant (τp) of the \( \dot{V}{\text{O}}_{2} \) response to a transition from rest-to-exercise, or between two different exercise intensities, is usually 20–35 s in healthy, young subjects (Whipp and Wasserman 1972). Shorter τp has been associated with improved exercise tolerance and performance in cycling, running and rowing (Whipp et al. 2002; Burnley and Jones 2007; Ingham et al. 2007). Previous studies have produced conflicting results regarding the relationship between τp and maximal oxygen uptake \( (\dot{V}{\text{O}}_{{ 2 {\text{max}}}} ) \). Some authors have shown τp to be shorter in athletes with higher \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) (Powers et al. 1985; Norris and Petersen 1998), whilst others have not (Barstow et al. 2000; Pringle et al. 2003). However, compelling evidence exists that \( \dot{V}{\text{O}}_{2} \) kinetics in young healthy subjects, when they perform rhythmic exercise, is limited by mitochondrial energetics (Poole et al. 2008), whereas \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) for exercise that involves large muscle mass is limited by O2 delivery (Basset and Howley 2000; Calbet et al. 2003; Saltin and Strange 1992). Given that \( \dot{V}{\text{O}}_{2} \) kinetics and \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) seem to differ in the extent to which they are modified by endurance training (Bailey et al. 2009; Demarle et al. 2001; Norris and Petersen 1998) as well as in the limiting factor, it seems logical that they may be dissociated from one other. Similarly, contradictory results have been presented as regards the extent to which the fast component of \( \dot{V}{\text{O}}_{2} \) kinetics is related to the first ventilatory threshold (Chilibeck et al. 1996; Kilding et al. 2007).
The tolerable duration of high-intensity exercise is characterised by a hyperbolic function of external power. Said asymptote is termed critical power (CP). The curvature of the hyperbola (W’) is mathematically equivalent to a constant amount of work that can be performed above CP, such that work capacity remains the same regardless of the work rate (Jones et al. 2010). Therefore, CP is usually defined as the highest sustainable rate of aerobic metabolism and W’ is taken to refer to an energy store comprising muscle ATP, PCr, glycogen and stored O2 (Hill 1993; Jones et al. 2010; Murgatroyd et al. 2011). For exercise intensities above the lactate threshold, the proximity of the power output to the so-called critical power determines the behaviour of the \( \dot{V}{\text{O}}_{2} \) slow component and therefore, time to exhaustion. Moreover, according to Henson et al. (1989), a slow component is always present in constant-load exercise above VT, irrespective of the fitness level of the subject. Murgatroyd et al. (2011) recently showed that the \( \dot{V}{\text{O}}_{2} \) slow component is positively correlated with the tolerable work above CP. Therefore, there is a close relationship between the P–t parameters (CP and W’) and \( \dot{V}{\text{O}}_{2} \) kinetics parameters during high-intensity exercise. This infers that \( \dot{V}{\text{O}}_{2} \) kinetics is a key determinant of exercise tolerance to high-intensity exercise, as proposed by Burnley and Jones (2007).
Since the \( \dot{V}{\text{O}}_{2} \) slow component was proven to be positively related to measures of anaerobic capacity (Berger and Jones 2007; Murgatroyd et al. 2011), we can expect the amplitude of the slow component to be inversely associated with VT.
To date, the investigation of \( \dot{V}{\text{O}}_{2} \) kinetics in swimming has been limited either by the use of only one repetition per exercise transition (Bentley et al. 2005; Fernandes et al. 2008) or by calculation of the \( \dot{V}{\text{O}}_{2} \) slow component as the difference between \( \dot{V}{\text{O}}_{2} \) values within a definite time-frame (such as between the end and the third or second minute of the exercise bout) (Demarie et al. 2001; Fernandes et al. 2003). Both such methods may lead to an impairment in the results so obtained (Bell et al. 2001; Lamarra et al. 1987). To our knowledge, only one study (Bentley et al. 2005) that possessed the methodological limitations mentioned above has described the primary phase \( \dot{V}{\text{O}}_{2} \) response in swimming exercise. This may be due to the technical difficulties that are involved in collection and adequate modelling of the breath-by-breath \( \dot{V}{\text{O}}_{2} \) signal during swimming.
Irrespective of the relative intensity of exercise, \( \dot{V}{\text{O}}_{2} \) kinetics also seems to be influenced by exercise mode. For example, previous studies reported both the τp and the \( \dot{V}{\text{O}}_{2} \) slow component to be smaller in running than in cycling, possibly due to different muscle contraction regimes and profiles of muscle fibre recruitment (Carter et al. 2000; Hill et al. 2003) As \( \dot{V}{\text{O}}_{2} \) kinetics during submaximal exercise is influenced by exercise mode (Carter et al. 2000; Hill et al. 2003), the extent of muscle mass recruited (Schneider et al. 2002) and body position (Koga et al. 1999), all of which factors are likely to differ between swimming and other terrestrial activities, both the \( \dot{V}{\text{O}}_{2} \) kinetics response to high-intensity swimming exercise and its relationship to other indicators of aerobic function (such as VT and \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \)), may differ from that which is observed within upright terrestrial exercise. \( \dot{V}{\text{O}}_{2} \) kinetics may also be differentially influenced by the greater respiratory work (Ogita and Tabata 1992), the specific thermoregulatory response (Koga et al. 1997) and the greater relative contribution of upper body work to the generation of total force (Hollander et al. 1988) that is involved in swimming.
Given that the recent literature (Burnley and Jones 2007; Murgatroyd et al. 2011) acknowledges high-intensity exercise tolerance to be determined by both the traditional aerobic parameters and \( \dot{V}{\text{O}}_{2} \) kinetics, the aim of this study was to determine the relationship between \( \dot{V}{\text{O}}_{2} \) kinetics in heavy-intensity swimming and “traditional” aerobic parameters such as \( \dot{V}{\text{O}}_{2\max } \) and VT. We hypothesised that (1) faster \( \dot{V}{\text{O}}_{2} \) kinetics would be positively correlated with the velocity associated with \( \dot{V}{\text{O}}_{2\max } \) \( (v\dot{V}{\text{O}}_{2\max } ) \) but not with \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) and VT, and that (2) the slow component amplitude would be inversely correlated with VT.
Methods
Twenty-one well trained male swimmers (mean ± SD, age: 20.2 ± 2.9 years; body mass: 73.1 ± 10.6 kg; height: 1.79 ± 0.07 m) gave their written informed consent to participate in the study. The swimmers were familiarised with the equipment and test procedures used in the study before it commenced. All the subjects trained at least eight times per week and had competed in their National Championships for at least the 5 years leading up to the study. The study was approved by the local University Ethical Committee, and conducted in accordance with the 1975 Declaration of Helsinki.
Each swimmer performed three testing sessions, each separated by at least 24-h rest, over a 10-day period. All the tests were performed in the same 50-m pool and involved freestyle swimming. In-water starts and open turns without underwater gliding were implemented throughout. The swimmers adjusted their speed within each test to that which was prescribed by the first investigator, on the basis of acoustic feedback given to them at every 25th metre. In the square-wave transitions, if the difference between the prescribed and the attained velocity was greater than 0.01 ms−1, the test was interrupted and repeated after 1 h of rest. Cardiorespiratory analysis of expired air was obtained within all test sessions using a breath-by-breath analyser (K4b2, Cosmed, Italy), connected to the swimmer by a respiratory snorkel and valve system (Aquatrainer, Cosmed, Italy) and calibrated according to the manufacturer’s instructions. The expiratory tube of the respiratory snorkel had a length of 1.83 m and internal diameter of 0.286 m, and the two-way non-re-breathing valve separating the inspiratory and expiratory tube had a volume of 45 ml. The inspiratory and expiratory resistances were minimal (9 cmH2O at 100 L min−1). The temperature of the flow-meter was corrected to ambient air temperature. All tests were conducted under the same environmental conditions (air temperature: 27.4 ± 0.4°C; water temperature: 27.0 ± 0.1°C; humidity: 59 ± 0.9) and between 10 a.m. and 2 p.m.
Incremental test
The swimmers first performed a discontinuous incremental test to voluntary exhaustion, comprising 5 × 200 m with 30-s rest intervals, for determination of \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) and the VT (Roels et al. 2005). The speed of the first 200 m repetition was set as 60% of the subject’s best season competition time for 200 m, and 5–10% speed increments between the first and fourth repetition were imposed. The last repetition was performed at maximal velocity.
\( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) was designated as the highest 30 s average of the \( \dot{V}{\text{O}}_{2} \) values so obtained (Roels et al. 2005). The velocity associated with \( \dot{V}{\text{O}}_{2\max } \) \( (v\dot{V}{\text{O}}_{2\max } ) \) was determined as the minimal velocity at which \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) was elicited (Billat and Koralsztein 1996). Using the aforesaid protocol, \( v\dot{V}{\text{O}}_{2\max } \) was always attained in the last repetition. VT was established as the oxygen uptake at which \( \dot{V}{\text{E}}/\dot{V}{\text{O}}_{2} \) began to increase without a simultaneous increase in \( \dot{V}{\text{E}}/\dot{V}{\text{CO}}_{2} \) (Wasserman et al. 1990). Fingertip blood was sampled immediately and 3, 5 and 7 min post-test, for the determination of maximal lactate concentration (Lamax) using a Lactate Pro portable analyser (Arkray, Kyoto, Japan).
On the second and third test occasion, the swimmers performed two 7-min square-wave transitions from rest to heavy-intensity swimming at Δ25% \( ({\text{VT}} + 0.25 \times (\dot{V}{\text{O}}_{2\max } - {\text{VT}})) \). Heart rate at the end of the exercise (end-exercise HR) was recorded telemetrically (Polar RS 800, Kempele, Finland). Fingertip blood lactate concentration was also determined immediately after the test, as described previously.
Data handling
Breath-by-breath values that were 3 SD from the local mean were removed from the analysis. The data were subsequently interpolated into 1-s values, time-aligned, and then ensemble averaged to provide a single on-transient set of data. \( \dot{V}{\text{O}}_{2} \) kinetics parameters were then calculated by an iterative procedure, whilst minimising the squared error between the modelled and the measured \( \dot{V}{\text{O}}_{2} \) values according to the following equation:
where \( \dot{V}{\text{O}}_{ 2} (t) \) represents the relative \( \dot{V}{\text{O}}_{2} \) at a given time, \( \dot{V}{\text{O}}_{{ 2 {\text{base}}}} \) represents rest \( \dot{V}{\text{O}}_{2} \) (i.e. the averaged \( \dot{V}{\text{O}}_{2} \) for the first 30 s of the last minute before exercise onset); tdp, τp, and A p, respectively, represent the time delay, the time constant and the amplitude of the primary component; and tdsc, τsc, A sc, represent the equivalent time delay, time constant and amplitude parameters for the \( \dot{V}{\text{O}}_{2} \) slow component.
End exercise \( \dot{V}{\text{O}}_{2} \) response was determined as the average for the last 30 s of the exercise bout.
To remove the influence of the cardiodynamic phase on the subsequent \( \dot{V}{\text{O}}_{2} \) response, we also chose to exclude the first 20 s of collected data from analysis (Borrani et al. 2001). Because the asymptotic value of the second function is not necessarily reached at the end of the exercise, the amplitude of the \( \dot{V}{\text{O}}_{2} \) slow component was defined as \( A_{\text{sc}}^{\prime } = A_{\text{sc}} (1 - {\text{e}}^{{ - ({\text{te}} - {\text{td}}_{\text{sc}} )/\tau_{\text{sc}} }} ) \), where te was the time at the end of the exercise bout (Borrani et al. 2001).
Our modelling procedures incorporated a previously reported (Reis et al. 2010) individual “snorkel delay” (ISD), so as to ensure that the VO2 kinetics data were representative of the exercise task. The ISD was calculated for each square-wave transition as the difference between the onset of exercise (t s), and the time (t ISD) at which the following breaths summed a tidal volume (TV) that was superior to volume of the outlet tube (RSV).
Statistical analysis
All statistical analyses were performed using the Statistical Package for the Social Sciences (SPSS Statistics, version 17.0 for Windows, SPSS Inc, Chicago, USA). The relationships between \( \dot{V}{\text{O}}_{2} \) kinetics parameters and \( \dot{V}{\text{O}}_{2\max } \), \( v\dot{V}{\text{O}}_{2\max } \) and VT were examined using Pearson’s product–moment correlation coefficient. Statistical significance was set at P < 0.05.
Results
The swimmers’ responses within both the incremental test and the square-wave transitions may be viewed in Tables 1 and 2, respectively. Figures 1 and 2 represent typical \( \dot{V}{\text{O}}_{2} \) versus time and \( \dot{V}{\text{O}}_{2} \) versus \( \dot{V}{\text{CO}}_{2} \) responses in the incremental test.

Example of the typical relationship between the \( \dot{V}{\text{O}}_{2} \) response and time in the incremental test

Example of the typical relationship between the \( \dot{V}{\text{O}}_{2} \) response (grey circles) and \( \mathop V\limits^{ \cdot } CO_{ 2} \) response (black triangles) in the incremental test
In all the swimmers, the best fit of the data was obtained when the model incorporated a slow component as opposed to a single exponential model.
The A p was positively correlated with relative \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) (r = 0.75, P = 0.00), absolute \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) (r = 0.44, P = 0.04), VT (r = 0.62, P = 0.00) and \( v\dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) (r = 0.53, P = 0.01). τp was inversely correlated with \( v\dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) (r = −0.55, P = 0.01) (Fig. 3), but was not related to either \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) (r = 0.05, ns) or VT (r = 0.14, ns).

Negative relationship between the τp and \( v\dot{V}{\text{O}}_{{ 2 {\text{max}}}} \). Linear regression equation and correlation values are indicated
The \( \dot{V}{\text{O}}_{2} \) slow component amplitude (%A′sc) was inversely correlated with VT (r = −0.61, P < 0.01) (Fig. 4), but not with either \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) (r = −0.14, ns) or \( v\dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) (r = 0.06, ns).

Negative relationship between %A′sc and VT. Linear regression equation and correlation values are indicated
Discussion
To our knowledge, this study is the first one in swimming to involve multiple square waves (i.e. more than a single repetition at the same exercise intensity) and bi-exponential modelling of the \( \dot{V}{\text{O}}_{2} \) response to determine \( \dot{V}{\text{O}}_{2} \) kinetics. We collected breath-by-breath data from two transitions for the same swimming intensity so that the signal/noise ratio could be increased, thus allowing for better characterisation of the primary phase of the \( \dot{V}{\text{O}}_{2} \) response.
The respiratory snorkel has been used in several studies in swimming (Bentley et al. 2005; Libicz et al. 2005; Roels et al. 2005), and it does not induce changes in either kinematics or efficiency over that evidenced within free swimming (Barbosa et al. 2010). A previous study from our research group found no significant differences in \( \dot{V}{\text{O}}_{2} \) measurements and \( \dot{V}{\text{O}}_{2} \) kinetics parameters, apart from a small difference in tdp, when the Aquatrainer swimming snorkel and a conventional facemask were used (Reis et al. 2010). Gayda et al. (2010) have stated that the Aquatrainer leads to an underestimation of \( \dot{V}{\text{O}}_{2} \). However, their study neither involved trained athletes nor an investigation of \( \dot{V}{\text{O}}_{2} \) kinetics. Furthermore, the authors did not correct the flow-meter temperature with the temperature of ambient air as is requested by the manufacturers, which may, at least partly, explain the discrepancies that were observed (Brugnoli 2010).
The τp values determined for heavy-intensity exercise were similar to those reported previously for running and rowing in trained athletes (Berger and Jones 2007; Borrani et al. 2001; Carter et al. 2000; Ingham et al. 2007). Although most of the studies in healthy young subjects have obtained τp values that are usually between 20 and 35 s (Poole and Jones 2005), trained athletes generally present faster kinetics (Borrani et al. 2001; Caputo et al. 2003; Ingham et al. 2007). It could be expected that the supine position (Koga et al. 1999), and the predominance of upper arm use, within swimming (Schneider et al. 2002) exercise, would induce slower kinetics than upright leg exercise. However, Cerretelli et al. (1979) have shown that the half-time of the \( \dot{V}{\text{O}}_{2} \) response depends on the training status of the exercising muscle. They also indicated that swimmers present a markedly faster response to supine exercise than do runners. In addition, endurance swim training has been shown to produce specific adaptations to the upper body muscles (Gollnick et al. 1972; Fitts et al. 1989). For example, arm-trained subjects have a higher relative proportion of slow-twitch fibres in their deltoid muscle (Gollnick et al. 1972; Tesch and Karlsson 1985) and a 10-day intensive swim training period has been shown to decrease the diameter of type II fibres in the deltoid muscle (Fitts et al. 1989). Swimmers also present higher citrate synthase activity in their deltoid muscle than untrained subjects (Fitts et al. 1989). Winlove et al. (2010) recently reported that pre-pubertal trained swimmers presented similar τp to untrained subjects in cycling, but 48% faster kinetics within arm-cranking exercise.
Overall, the average values reported in our study for τp (17.3 s) are very close to those (16.9 s) reported by Bentley et al. (2005) for a single 400-m swimming exercise transition at the same relative velocity. The effects of both the specific adaptations of the upper arms to swim training, and the contribution of the legs to total energy expenditure (Hollander et al. 1988; Fitts et al. 1989) could explain the fast kinetics that was detected in well-trained swimmers in both our study and by previous work (Cerretelli et al. 1979; Bentley et al. 2005).
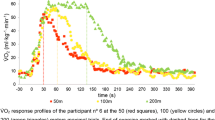
We did not observe a significant correlation between τp and either \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) or VT within submaximal swimming exercise. Whipp et al. (2002), have stated that τp is not a good predictor of individual fitness level. Previous studies showed that individuals with similar τp could have widely varying \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) values and, conversely, that individuals with similar \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) could present wide ranges of τp. In our study, two subjects with \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) values of 56–57 ml kg−1 min−1 presented a τp of 12 and 24 s, respectively (Fig. 5), which is in line with the results of Whipp et al. (2002) where athletes of similar \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) (50 ml kg−1 min−1) had a large difference in τp (20–60 s).

\( \dot{V}{\text{O}}_{2} \) response profiles in two subjects with similar \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) but different τp. Swimmer 1 (grey circles) and swimmer 2 (black circles). Grey and black lines represent the best fit as determined from the exponential modelling procedure for swimmers 1 and 2, respectively
The observed lack of a statistically significant relationship between \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) and the time constant is unsurprising, given the differing extent to which said variables are sensitive to training adaptations (Norris and Petersen 1998; Demarle et al. 2001). Our results seem to confirm that \( \dot{V}{\text{O}}_{2} \) kinetics and \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) are based on different physiological mechanisms: whilst, for exercises involving large muscle mass, maximal oxygen uptake is determined by central mechanisms, such as O2 delivery (Saltin and Strange 1992), and it seems that this is not the case for \( \dot{V}{\text{O}}_{2} \) kinetics within heavy-intensity constant exercise in young, healthy subjects (Poole and Jones 2005). Due to the inherent methodological limitations to the investigation of swimming exercise, we neither determined muscle deoxygenation nor fibre recruitment. However, the fast τp that we observed appears to indicate that although blood flow may be impaired in swimming as a result of the horizontal body posture (Holmer 1972), O2 availability is not sufficiently compromised for \( \dot{V}{\text{O}}_{2} \) kinetics to be slowed down relative to that of trained runners, rowers or cyclists (Carter et al. 2000; Borrani et al. 2001; Ingham et al. 2007).
Interestingly, we observed a significant correlation between τp and \( v\dot{V}{\text{O}}_{2\max } \). \( v\dot{V}{\text{O}}_{2\max } \) is said to explain individual differences in performance more fully than \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) (Billat and Koralsztein 1996). Shorter τp is associated with smaller oxygen deficits and smaller decreases of muscle high-energy phosphates, enhanced time to exhaustion (Demarle et al. 2001), improved exercise tolerance (Whipp et al. 2002) and improved performance (Whipp et al. 2002; Burnley and Jones 2007; Ingham et al. 2007). The relationship between τp and \( v\dot{V}{\text{O}}_{2\max } \) may be due to an increased oxidative contribution to energy transfer, induced by faster kinetics. Faster \( \dot{V}{\text{O}}_{2} \) adaptations to increased metabolic demand, will minimise the O2 deficit, preserving the integrity of the intramyocyte milieu and, consequently, the capacity to sustain high-intensity exercise (Bailey et al. 2009; Burnley and Jones 2007; Hepple 2002). A reduction in the O2 deficit decreases the depletion of muscle creatine phosphate and glycogen, and the accumulation of metabolites that may be associated with fatigue (such as ADP, inorganic phosphate and H+). Thus, for a given magnitude of O2 deficit, a faster \( \dot{V}{\text{O}}_{2} \) kinetics profile corresponds to higher power outputs (Burnley and Jones 2007; Murgatroyd et al. 2011). Our data are in line with those of the latter authors.
The existence of a slow component during swimming bouts performed above the ventilatory threshold was confirmed in this study. The values that we report in this paper (8.9%) are similar to those reported by Fernandes et al. (2008) (8.2%), smaller than those reported by Demarie et al. (2001) (14%), and higher than those reported by Fernandes et al. (2003) (5.0%). However, the aforementioned authors either determined the slow component as the \( \dot{V}{\text{O}}_{2} \) difference between the sixth and second or third minute of exercise, or analysed only one exercise transition, so impairing the comparison of their results with those of other studies (Bell et al. 2001; Lamarra et al. 1987). As has been observed in other studies, great inter-individual variability in the \( \dot{V}{\text{O}}_{2} \) slow component exists among swimmers (Bentley et al. 2005; Fernandes et al. 2008). The increase of drag, the lower mechanical efficiency of swimming and the disruption caused by turns makes it difficult to compare the slow component with different terrestrial activities.
The relative contribution of the slow component to end-exercise \( \dot{V}{\text{O}}_{2} \) (%A′sc) was not related to either \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) or \( v\dot{V}{\text{O}}_{2\max } \), but was inversely correlated with VT. The ventilatory threshold reflects or is concomitant with the increase in lactate concentration above baseline values, signalling altered phosphorylation and redox potentials in the muscle fibres that are involved (Jones and Carter 2000). Previous studies have associated smaller slow components with higher percentages of type I fibres in the exercising muscle(s) (Barstow et al. 1996; Pringle et al. 2003; Russel et al. 2002). Although we did not study muscle fibre recruitment, as type I fibres have greater oxidative capacity (Crow and Kushmerick 1982), it seems logical that the enhanced recruitment of this muscle fibre type—resulting either from training (Andersen and Henriksson 1977; Spina et al. 1996) or a genetic predisposition (Sale et al. 1990)—would induce both a diminished slow component and a higher ventilatory threshold. Our results agree with those of Murgatroyd et al. (2011), who demonstrated a positive correlation between the quantity of tolerable work above the critical power and the \( \dot{V}{\text{O}}_{2} \) slow component. The \( \dot{V}{\text{O}}_{2} \) slow component reflects an increase in the O2 cost of supra-VT exercise, and thus a reduction in work efficiency, that may be attributed to the recruitment of additional (mostly type II, i.e. less efficient) muscle fibres or to a reduction in the efficiency of the working fibres (Barstow et al. 1996; Cannon et al. 2011; Pringle et al. 2003). A reduction in the amplitude of the \( \dot{V}{\text{O}}_{2} \) slow component would both signify an improved exercise economy/efficiency, and be influenced by the elevation of the VT. Therefore, an increase of VT towards \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) reduces the room for the slow component to develop. The negative relationship between the \( \dot{V}{\text{O}}_{2} \) slow component and VT in the present study is in line with previous findings of a negative correlation between the \( \dot{V}{\text{O}}_{2} \) slow component, anaerobic work capacity (Murgatroyd et al. 2011) and performance in a Wingate test (Berger and Jones 2007).
The amplitude of the slow component was not associated with \( v\dot{V}{\text{O}}_{2\max } \) of the present swimmers. This is in line with previous results obtained in rowing, where the slow component was unable to differentiate between elite and club-level athletes (Ingham et al. 2007).
Conclusion
This study was the first to describe the time constant in heavy-intensity swimming using specific swimming exercise and appropriate methods (e.g. multiple exercise transitions and bi-exponential fitting). Faster kinetics in heavy-intensity swimming is associated with the maximal velocity attained in an incremental swimming test designed to elicit \( \dot{V}{\text{O}}_{{ 2 {\text{max}}}} \) in accordance with what has been previously described in other exercise modes. In addition, the slow component seems to be reduced in swimmers with higher ventilatory thresholds.
References
Andersen P, Henriksson J (1977) Training induced changes in the subgroups of human type II skeletal muscle fibres. Acta Physiol Scan 99(1):123–125
Bailey SJ, Wilkerson DP, Dimenna FJ, Jones AM (2009) Influence of sprint training on pulmonary O2 uptake and muscle deoxygenation kinetics in humans. J Appl Physiol 106(6):1875–1887
Barbosa T, Silva AJ, Reis AM, Costa M, Garrido N, Policarpo F, Reis VM (2010) Kinematical changes in swimming front crawl and breastroke with the Aquatrainer snorkel. Eur J Appl Physiol 109(6):155–162
Barstow TJ, Jones AM, Nguyen PH, Casaburi R (1996) Influence of muscle fiber type and pedal frequency on oxygen uptake kinetics in heavy exercise. J Appl Physiol 81(4):1642–1650
Barstow TJ, Jones AM, Nguyen PH, Casaburi R (2000) Influence of muscle fibre type and fitness on the oxygen uptake/power output slope during incremental exercise in humans. Exp Physiol 85(1):109–116
Basset DR, Howley ET (2000) Limiting factors for maximum oxygen uptake and determinants of endurance performance. Med Sci Sports Exerc 32(1):70–84
Bell C, Paterson DH, Kowalchuck JM, Padilla J, Cunningham DA (2001) A comparison of modelling techniques used to characterise oxygen uptake kinetics during the on-transient of exercise. Exp Physiol 86(5):667–676
Bentley DJ, Roels B, Hellard P, Fauquet C, Libicz S, Millet GP (2005) Physiological responses during submaximal interval swimming training: effects of interval duration. J Sci Med Sport 8:392–402
Berger NJ, Jones AM (2007) Pulmonary uptake on-kinetics in sprint- and endurance-trained athletes. Appl Physiol Nutr Metab 32(3):383–393
Billat LV, Koralsztein JP (1996) Significance of the velocity at VO2max and time to exhaustion at this velocity. Sports Med 22(2):90–108
Borrani F, Candau R, Millet GY, Perrey S, Fuchslocher J, Rouillon JD (2001) Is the VO2 slow component dependent on progressive recruitment of fast-twitch fibers in trained runners? J Appl Physiol 90(6):2212–2220
Brugnoli P (2010) Letter to the editor of EJAP. Re: comparison of gas exchange data using the Aquatrainer® system and the facemask with Cosmed K4b2 during exercise in healthy subjects. Eur J Appl Physiol 110(4):875–876
Burnley M, Jones AM (2007) Oxygen uptake kinetics as a determinant of sports performance. Eur J Sport Sci 7(2):63–69
Calbet JA, Boushel R, Rådegran C, Søndergaard H, Wagner PD, Saltin B (2003) Determinants of maximal oxygen uptake in severe acute hypoxia. Am J Physiol Regul Integr Comp Physiol 284(2):R291–R303
Cannon DT, White AC, Andriano MF, Kolkhorst FW, Rossiter HB (2011) Skeletal muscle fatigue precedes the slow component of oxygen uptake kinetics during exercise in humans. J Physiol 589(Pt 3):727–739
Caputo F, Mello MT, Denadai BS (2003) Oxygen uptake kinetics and time to exhaustion in cycling and running: a comparision between trained and untrained subjects. Arch Physiol Biochem 111(5):461–466
Carter H, Jones AM, Barstow TJ, Burnley M, Williams CA, Doust JH (2000) Oxygen uptake kinetics in treadmill running and cycle ergometry: a comparison. J Appl Physiol 89:899–907
Cerretelli P, Pendergast D, Paganelli WC, Rennie DW (1979) Effects of specific muscle training on VO2 on-response and early blood lactate. J Appl Physiol 47(4):761–769
Chilibeck PD, Paterson DH, Petrella RJ, Cunningham DA (1996) The influence of age and cardiorespiratory fitness on kinetics of oxygen uptake. Can J Appl Physiol 21(3):1331–1338
Crow MT, Kushmerick MJ (1982) Chemical energetics of slow- and fast-twitch muscles of the mouse. J Gen Physiol 79:147–166
Demarie S, Sardella F, Billat V, Magini W, Faina M (2001) The VO2 slow component in swimming. Eur J Appl Physiol 84:95–99
Demarle AP, Slawinski JJ, Laffite LP, Bocquet VG, Koralsztein JP, Billat V (2001) Decrease of O2 deficit is a potential factor in increased time to exhaustion after specific endurance training. J Appl Physiol 90(3):947–953
Fernandes RJ, Cardoso CS, Soares SM, Ascensão A, Colaço PJ, Vilas-Boas JP (2003) Time limit and VO2 slow component at intensities corresponding to VO2max in swimmers. Int J Sports Med 24(8):576–581
Fernandes RJ, Keskinen KL, Colaço P, Querido AJ, Machado LJ, Morais PA, Novais DQ, Marinho DA, Vilas Boas JP (2008) Time limit at VO2max velocity in elite crawl swimmers. Int J Sports Med 29:145–150
Fitts RH, Costill DL, Gardetto PR (1989) Effect of swim exercise training on human muscle fiber function. J Appl Physiol 66(1):465–475
Gayda M, Bosquet L, Juneau M, Guiraud T, Lambert J, Nigam A (2010) Comparison of gas exchange data using the Aquatrainer® sytem and the facemask with Cosmed K4b2 during exercise in healthy subjects. Eur J Appl Physiol 109(2):191–199
Gollnick PD, Armstrong RB, Saubert CW, Piehl K, Saltin B (1972) Enzyme activity and fiber composition in skeletal muscle of untrained and trained men. J Appl Physiol 33:312–319
Henson LC, Poole DC, Whipp BJ (1989) Fitness as a determinant of oxygen uptake response to constant-load exercise. J Appl Physiol 59:21–28
Hepple RT (2002) The role of O2 supply in muscle fatigue. Can J Appl Physiol 27(1):56–69
Hill DW (1993) The critical power concept. A review. Sports Med 16(4):237–254
Hill DW, Halcomb JN, Stevens EC (2003) Oxygen uptake kinetics during severe intensity running and cycling. Eur J Appl Physiol 89:612–618
Hollander AP, Groot GJ, Schneau VI, Kahman R, Toussaint HM (1988) Contributions of the legs to propulsion in front crawl swimming. In: Undregerechts BE, Wilkie K, Reischle K (eds) Swimming science V. Human Kinetics, Champaign, pp 39–43
Holmer I (1972) Oxygen uptake during swimming in man. J Appl Physiol 33:502–509
Ingham SA, Carter H, Whyte G, Doust JH (2007) Comparison of the oxygen uptake kinetics of club and olympic champion rowers. Med Sci Sports Exerc 39(5):865–871
Jones AM, Carter H (2000) The effect of endurance training on parameters of aerobic fitness. Sports Med 29(6):373–386
Jones AM, Vanhatalo A, Burnley M, Morton RH, Poole DC (2010) Critical power: implications for determination of VO2max and exercise tolerance. Med Sci Sports Exerc 42(10):1876–1890
Kilding AE, Fysh M, Winter EM (2007) Relationships between pulmonary oxygen uptake kinetics and other measures of aerobic fitness in middle- and long-distance runners. Eur J Appl Physiol 100:105–114
Koga S, Tomoyuki S, Kondo N, Barstow T (1997) Effect of increased muscle temperature on oxygen uptake kinetics during exercise. J Appl Physiol 83(4):1333–1338
Koga S, Tomoyuki S, Shibasaki M, Kondo N, Fukuba Y, Barstow T (1999) Kinetics of oxygen uptake during supine and upright heavy exercise. J Appl Physiol 87(1):253–260
Lamarra N, Whipp BJ, Ward SA, Wasserman K (1987) Effect of interbreath fluctuations on characterizing exercise gas exchange kinetics. J Appl Physiol 62:2003–2012
Libicz S, Roels B, Millet GP (2005) VO2 responses to intermittent swimming sets at velocity associated with VO2max. Can J Appl Physiol 30(5):543–553
Murgatroyd SR, Fergunson C, Ward SA, Whipp BJ, Rossiter HB (2011) Pulmonary O2 uptake kinetics as a determinant of high-intensity exercise tolerance in humans. J Appl Physiol. doi:10.1152/japplphysiol.01092.2010 (in press)
Norris SR, Petersen SR (1998) Effects of endurance training on transient oxygen uptake responses in cyclists. J Sports Sci 16:733–738
Ogita F, Tabata I (1992) Oxygen uptake kinetics during swimming in a hypobaric hypoxic environment. Eur J Appl Physiol 65:192–196
Poole D, Jones AM (2005) Understanding the mechanistic bases of VO2 kinetics. In: Jones AM, Poole DC (eds) Oxygen uptake kinetics in sport. Exercise and Medicine, Routledge, pp 294–328
Poole DC, Schaffartzik W, Knight DR, Derion T, Kennedy B, Guy HJ, Prediletto R, Wagner PD (1991) Contribution of exercising legs to the slow component of oxygen uptake kinetics in humans. J Appl Physiol 71(4):1245–1260
Poole DC, Barstow TJ, Mcdonough P, Jones AM (2008) Control of oxygen uptake during exercise. Med Sci Sport Exerc 40(3):462–474
Powers SK, Dodd S, Beadle RE (1985) Oxygen uptake kinetics in trained athletes differing in VO2 max. Eur J Appl Physiol 54:306–308
Pringle JS, Doust JH, Carter H, Tolfrey K, Campbell IT, Sakkas GK, Jones AM (2003) Oxygen uptake kinetics during moderate, heavy and severe intensity submaximal exercise in humans: the influence of muscle type and capillarisation. Eur J Appl Physiol 89:289–300
Reis JF, Millet GP, Malatesta D, Roels B, Borrani F, Vleck VE, Alves FB (2010) Are oxygen uptake kinetics modified when using a respiratory snorkel? Int J Sports Physiol Perform 5(3):292–300
Roels B, Schmitt L, Libicz S, Bentley D, Richalet JP, Millet GP (2005) Specificity of VO2 max and the ventilatory threshold in free swimming and cycle ergometry: comparison between triathletes and swimmers. Br J Sports Med 39:965–968
Russel A, Wadley G, Snow R, Giacobino JP, Muzzin P, Garnham A, Cameron-Smith D (2002) Slow component of VO2 kinetics: the effect of training status, fibre type, UCP3 mRNA and citrate synthase activity. Int J Obes Relat Metab Disord 26(2):157–164
Sale DG, MacDougall JD, Jacobs I, Garner S (1990) Interaction between concurrent strength and endurance training. J Appl Physiol 68(1):260–270
Saltin B, Strange S (1992) Maximal oxygen uptake: “old” and “new” arguments for a cardiovascular limitation. Med Sci Sports Exerc 24:30–37
Schneider DA, Wing A, Morris NR (2002) Oxygen uptake and heart rate during heavy exercise: a comparision between arm cranking and leg cycling. Eur J Appl Physiol 88:100–106
Spina RJ, Chi MM, Hopkins MG, Nemeth PM, Lowry OH, Holloszy JO (1996) Mitochondrial enzymes increase in muscle response to 7–10 days of cycle exercise. J Appl Physiol 80(6):2250–2254
Tesch PA, Karlsson J (1985) Muscle fiber types and size in trained and untrained muscles of elite athletes. J Appl Physiol 59(6):1716–1720
Vanhatalo A, Poole DC, DiMenna FJ, Bailey SJ, Jones AM (2011) Muscle fiber recruitment and the slow component of O2 uptake: constant work rate vs. all-out sprint exercise. Am J Physiol Regul Integr Comp Physiol 300(3):R700–R707
Wasserman K, Beaver WL, Whipp BJ (1990) Gas exchange theory and the lactate acidosis (anaerobic) threshold. Circulation 81(1 Suppl II):14–30
Whipp BJ, Wasserman K (1972) Oxygen uptake kinetics for various intensities of constant-load work. J Appl Physiol 33:351–356
Whipp BJ, Ward SA, Lamarra N, Davis JA, Wasserman K (1982) Parameters of ventilatory and gas exchange dynamics during exercise. J Appl Physiol 52(6):1506–1513
Whipp BJ, Rossiter HB, Ward SA (2002) Exertional oxygen uptake kinetics: a stamen of stamina? Biochem Soc Trans 30(2):237–247
Winlove MA, Jones AM, Welsman JR (2010) Influence of training status and exercise modality on pulmonary O2 uptake kinetics in pre-pubertal girls. Eur J Appl Physiol 108(6):1169–1179
Acknowledgments
The first author gratefully acknowledges the “Fundação para a Ciência e Tecnologia, Portugal” (“The Foundation for Science and Technology, Portugal”) for their doctoral fellowship award (reference number SFRH/BD/23351/2005). VV also gratefully acknowledges the “Fundação para a Ciência e Tecnologia, Portugal” for their award of a post-doctoral research fellowship under the “Ciência 2008” programme.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by David C. Poole.
Rights and permissions
About this article
Cite this article
Reis, J.F., Alves, F.B., Bruno, P.M. et al. Effects of aerobic fitness on oxygen uptake kinetics in heavy intensity swimming. Eur J Appl Physiol 112, 1689–1697 (2012). https://doi.org/10.1007/s00421-011-2126-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-011-2126-6