
Abstract
The aim of this study was to determine whether muscle stiffness measured in vivo was different between males and females. Distal displacement of the gastrocnemius medialis myotendinous junction was measured directly using ultrasonography during passive dorsiflexion in eight males and eight females (age range 19–28 years). Plantarflexion torque and myotendinous junction displacement were measured at 5° intervals, where 0° was with the foot at right angles to the tibia. Stiffness of the gastrocnemius medialis muscle was calculated between 0° and 25° of dorsiflexion, and defined as passive plantarflexion torque/distal displacement of the myotendinous junction (N m cm−1). Relative muscle stiffness was also calculated as distal displacement relative to resting muscle length, and as passive torque relative to plantarflexion maximal voluntary contraction torque. No significant gender difference was observed in passive dorsiflexion torque, or in passive torque/maximal voluntary torque throughout the range of motion. Distal displacement of the gastrocnemius myotendinous junction was 26% more in females than in males (P < 0.05). Myotendinous junction displacement was 5.0 ± 1.4% of resting gastrocnemius medialis length in females, and 3.9 ± 0.6% in males. Over 25° of passive dorsiflexion, gastrocnemius medialis muscle stiffness was greater in males than in females by 44% (P < 0.05). In conclusion, based on the in vivo assessment of myotendinous junction displacement, passive gastrocnemius medialis muscle stiffness is greater in males than in females.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The passive torque–joint angle relation is a commonly adopted technique for assessing flexibility and is often used as an indicator of muscle-tendon unit (MTU) stiffness about a joint (McHugh et al. 1998; Taylor et al. 1990). However, based on the passive torque generated by movement of the whole limb (here defined as MTU stiffness), it is impossible to state to what extent the muscle or tendon contribute to differences observed in passive torque of the MTU as a whole (Morse et al. 2008). Nonetheless, the passive torque–angle relation is commonly used to assess flexibility and mitigates some of the shortcomings, such as pain tolerance, associated with assessments to volitional end range of motion (ROM) (McHugh et al. 1998). To this end, MTU stiffness, as assessed via the passive torque–angle relation, is reported to be greater in men than in women (Blackburn et al. 2004; Gajdosik et al. 1990). However, based on these measured gender differences in the passive torque-joint angle relation, it may be erroneous to make conclusions regarding muscle stiffness (here defined as the load elongation characteristics of the muscle distal to the myotendinous junction, MTJ), as the muscle and tendon are known to contribute to the length change in MTU to varying degrees under stretch when assessed in vivo using ultrasonography (Morse et al. 2008).
At present, the in vivo determinants of the gender difference in MTU stiffness can only be alluded to from studies on the viscoelastic properties of the tendon. Specifically, Kubo et al. (2003) reported that females have a greater tendon compliance than males during voluntary plantarflexion contractions, which is consistent with the observed relation between oestrogen levels and tendon compliance (Bryant et al. 2008). However, tendon compliance assessed under loading may be of little consequence to passive torque (Kubo et al. 2001), with passive muscle stiffness being of greater significance in the assessment of flexibility (Morse et al. 2008). It seems pertinent therefore that to make conclusions regarding gender differences in muscle stiffness specifically, tracking of the MTJ under stretch is necessary rather than relying on the torque–angle relation. As yet, however, the role that muscle stiffness plays in gender difference of passive torque remains unreported. Therefore, the aim of this investigation was to determine whether there is a gender difference in the passive stiffness of the gastrocnemius muscle. It is hypothesised that based on previous observations on tendon stiffness, and MTU stiffness as a whole, muscle stiffness will be higher in men than in women.
Methods
Participants
Eight males (age 21 ± 1 years, stature 1.76 ± 0.08 m, mass 77.8 ± 15.2 kg) and eight females (age 24 ± 3 years, stature 1.63 ± 0.07 m, mass 59.6 ± 4.7 kg) volunteered to participate in this study. Participant demographics are presented as means ± SD.
All were recreationally active but not involved in any structured physical training regime and were free from lower limb injury. None of the females were taking the contraceptive pill. All participants received experimental procedure information sheets; these outlined the experimental protocol in lay terms, and the proposed techniques were clarified verbally by the investigator. Written informed consent was obtained and all procedures were approved by the Local Ethics Committee of Manchester Metropolitan University. The study conformed to the principles set out in the Declaration of Helsinki.
Experimental setup
Participants were secured about the hip and shoulders in a prone position on a dynamometer (Cybex Norm, Cybex International Inc., NY, USA) with their arms against their side. The knee was in full extension and the foot attached securely to the foot plate of the dynamometer so the ankle joint was aligned with the axis of the dynamometer. All the participants were tested on their verbally, self-reported dominant limb. An electrogoniometer (K100, Biometrics Ltd, UK) was attached at the ankle, and the foot was strapped to minimize heel displacement during dorsiflexion: all reported angle measurements refer to the ankle joint assessed with the goniometer, not the angle of the dynamometer foot plate. The passive ROM was determined using an approach similar to that adopted by others (Magnusson et al. 1997; McHugh et al. 1998); this involved passive isokinetic dorsiflexion at 1° s−1, starting from +10° (the foot at right angles to the leg being 0°), until discomfort caused the participant to stop the dynamometer by activating a safety trigger. The angle at which this occurred was taken as the end ROM. Measures of MTJ displacement and passive torque were recorded during passive dorsiflexion at 1° s−1 to end ROM.
Passive torque
Torque was displayed on a computer screen, interfaced with an acquisition system (Acknowledge, Biopac Systems, Santa Barbara, CA, USA) used for analogue-to-digital conversion. The sampling frequency was 2,000 Hz. Each torque signal was filtered with a low-pass fourth-order Butterworth filter with a 30-Hz cut-off frequency. Torque was calibrated as linear up to 180 N m. As the torque experienced with passive loading is much lower than MVC, smaller seven calibration increments of 0.9 N masses were added to the cybex, providing calibration torques up to 3 N m. Over this range typical error of estimate was 0.03 N m with 90% CL of ±0.04 N m, R 2 was 0.999. Signal to noise ratios were confirmed as 0.15 N m at a calibration load of 10.8 N m.
Isometric maximal voluntary contraction torque
Maximal isometric voluntary plantar flexion contractions (MVC) were performed 30 min after the stretch procedures took place. Participants performed a series of three submaximal isokinetic contractions (90° s−1) and two submaximal isometric contractions at an ankle angle of 0° as a warm up. MVC’s were performed at 0° and maintained for 4 s (sufficient to reach a plateau) with 2 min of rest in between each contraction to prevent development of fatigue. To maximize performance, visual feedback of the torque signal and verbal encouragement was given to all participants. The highest torque reached during a contraction was recorded, and the highest value of the two contractions was given as the MVC.
In vivo measurements of muscle and tendon properties
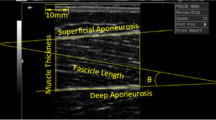
B-Mode ultrasonography (MyLab 50, Esaote, Genoa, Italy) was used to determine the displacement of the MTJ of the gastrocnemius medialis (GM) during passive dorsiflexion. The MTJ was identified as described by Maganaris and Paul (1999), as the distal join between the deep and superficial aponeuroses of the GM muscle. Distal displacement of the GM MTJ was visualized and recorded as a continuous sagittal plane, ultrasound video, using a 10 cm, 7.5 MHz linear-array probe (Fig. 1). The ultrasound image was time locked with the torque and goniometer outputs. Displacement was measured relative to an acoustically reflective marker (a thin strip of micro-pore tape) secured to the skin proximal to the GM myotendinous junction. Images were recorded on digital tape at 30 Hz and analysed offline every 5° from 0° to 25° dorsiflexion using digital tracking software (Dartfish, Friburg, Switzerland).

a Sample sagittal plane US image during passive dorsiflexion, with the GM MTJ visible as the point of intersect of the deep and superficial aponeurosis. Vertical dashed line denotes the echo absorptive marker placed on the dermal surface. b A sample calibration scan using a digital micrometer to cast two vertical shadows on the US image
The total MTU length was determined at the start of the experiment with an inextensible tape laid over the surface of the GM muscle and tendon and using ultrasound to identify the anatomical landmarks of the GM origin, the insertion at the MTJ and the insertion of the Achilles tendon. Tendon length is reported as the free GM tendon and the Achilles tendon.
Validity of the static and dynamic ultrasound measurements has been discussed previously (Morse et al. 2008). Based on the visual tracking technique adopted in the present study, Mahieu et al. (2004) reported that during gastrocnemius contraction there was a 9% SEM, with a 6.2 mm proximal displacement of the MTJ, and ICC of 0.96. However, in order to ensure validity of MTJ displacement under smaller passive torques, the accuracy of the US was assessed by recording 1-mm increments on a digital micrometer that was accurate to 0.01 mm (Mitutoyo Corp., Tokyo). Briefly, using the analysis procedures described above, 1-mm incremental gaps were measured between the callipers of the micrometer from 1 to 10 mm (Fig. 1). The mean error using the visual tracking US technique was 0.06 mm, with 90% confidence limits (CL) ± 0.02 mm, and R 2 of 0.999. The adopted technique in the present study should be considered sensitive enough to identify the visual tracking of the MTJ during distal displacement under stretch, previously observed at 9.2 mm in males (Morse et al. 2008).
It should be noted that although it is possible to allocate contractile forces based on relative PCSA under loading (Morse et al. 2005), it is as yet not possible to determine accurately the total contribution of the GM to the measured passive plantar flexion (PF) torque (see “Discussion”); therefore with this caveat in mind nominal muscle stiffness is presented as the ratio of passive PF torque (N m)/distal displacement of the GM MTJ (cm). To account for any difference in GM muscle length and MVC, muscle stiffness is also presented as distal displacement relative to GM muscle length at 0°, and as passive torque relative to MVC torque.
Statistics
For variables with between and within measures (passive torque, distal displacement, nominal stiffness) significant difference was determined using a repeated measures ANOVA; within participant variables: angle (6 levels), and between participant: gender. If a significant interaction was found, gender differences were determined with independent t tests. For variables with only a between-group main effect, with no within-group comparison, impendent t tests were performed (e.g. ROM, MVC, static muscle length). Differences were considered significant at an alpha level of P < 0.05. For clarity on figures, data are reported as mean ± SEM unless otherwise indicated.
Results
Passive torque showed a significant angle effect with no gender by angle interaction; therefore, no planned comparisons were performed for passive torque (Fig. 2). PF MVC was significantly higher in males than in females (P < 0.01, Table 1). When expressed relative to MVC torque, passive torque was not significantly different between males and females at any angle. The relative torque followed the same curvilinear relation with ankle angle as passive torque, reaching a maximum value of 27.5 ± 9.9% of MVC in males and 31.1 ± 11.4% of MVC in females throughout the passive stretch.

Passive plantarflexion torque during passive dorsiflexion in males (closed circles), and females (open circles). No significant group effect was observed in passive torque
Throughout the passive stretch distal displacement of the GM MTJ was significantly greater in women than in men between 10° and 25° (Fig. 3, P < 0.05), with ~0.35 cm greater displacement in the females compared to the males from beyond an ankle angle of 10°.

Distal displacement of the GM MTJ in males (closed circles) and females (open circles). Asterisks denote significant difference from males (P < 0.05)
There was no difference in resting GM length or tendon length at 0° prior to passive dorsiflexion (Table 1). MTJ displacement relative to GM length was greater in Females compared to males between 10° and 25° (Fig. 4, P < 0.05), with distal displacement reaching 5.0 ± 1.4% of resting GM length in females, and 3.9 ± 0.6% in males at 25°. Nominal muscle stiffness over the 25° passive stretch was 44% greater in males than in females (Fig. 5; Table 1, P < 0.05).

Distal displacement of the GM MTJ relative to resting muscle length (Lm) in males (closed circles) and females (open circles). Asterisks denote significant difference from males (P < 0.05)

Passive torque displacement relation in males (closed circles) and females (open circles)
Discussion
The main finding of the present investigation was that muscle stiffness, determined by the lengthening of the GM muscle under stretch, is greater in males than in females. This in vivo data on a gender difference in muscle stiffness under stretch has only ever been eluded to indirectly through the torque–angle relation (Gajdosik et al. 1990) and is consistent with previous studies that have shown a gender difference in the viscoelastic properties of the tendon under loading (Kubo et al. 2003).
Previously, gender differences in flexibility and the passive torque angle relationship of the entire MTU have led to conclusions of lower muscle stiffness in females (Blackburn et al. 2004; Gajdosik et al. 1990). However, as recently demonstrated the contribution of the muscle and tendon to length changes in the MTU under stretch are essentially equal (Morse et al. 2008). Therefore, it may be erroneous to make conclusions regarding the muscle’s contribution to gender differences in passive torque, when the passive torque of the MTU has been measured as a whole. In the present investigation, muscle elongation under stretch was measured directly by recording the distal displacement of the MTJ using ultrasonography. Although there are a number of well-reported limitations to this in vivo technique, ultrasound tracking of the MTJ is essential for making explicit conclusions on muscle stiffness. One such limiting factor to the in vivo tracking of the distal GM MTJ is that it is impossible to state whether the proximal tendon contributed to the distal displacement of the MTJ (Onambele et al. 2007). It should be noted that the proximal GM tendon is almost universally ignored in the assessment of tendon stiffness during plantarflexion (Maganaris and Paul 2002); however, as tendon elongation (strain) is dependent on tendon length (Maganaris and Paul 1999), and in the present investigation MTU length is not significantly different, the contribution of the proximal gastrocnemius tendon to gender differences is likely to be negligible in the present study, although it cannot be entirely discounted. Further to the possible contribution of the proximal tendon, an additional limitation of the in vivo technique that must be acknowledged is that by measuring the distal displacement of the GM MTJ, as in the present study, it is impossible to account for the contribution of the extracellular connective tissue matrix to the passive tension (Purslow 1989). Therefore, with the limitations of the in vivo technique in mind and to ensure there is clarity in the conclusions drawn from the present data, it is pertinent to state that muscle stiffness has been defined within this study as the muscle and connective tissue proximal to the GM MTJ. It remains that a gender difference exists in the distal displacement of the GM MTJ under stretch, and using in vivo tracking of the MTJ, confirms what has been alluded to previously from the passive torque–angle relation, that muscle stiffness is lower in females than in males.
There are a number of possible contributory factors to the observed gender difference in muscle stiffness in the present study. Muscle stiffness is determined by neural activity within the muscle, its cross-sectional area (CSA) and the viscoelastic properties of the muscle under loading (Magnusson et al. 1997). The contribution of muscle activity to gender differences in distal displacement is likely to be negligible. In the present investigation activation was not measured; however, using an identical experimental set-up it was previously established that the EMG output of the GM muscle remained below 1% of that measured during MVC in males (Morse et al. 2008). It is unlikely that this 1% EMG activity previously observed in the GM of males could be measurably lower in the present females and is therefore unlikely to contribute to the observation of a greater GM muscle stiffness in males. At present the only data on gender differences in muscle activity and stiffness are during repeated dynamic loading tasks, such as hopping, or from transient knee extension perturbations (Granata et al. 2002; Padua et al. 2005). Although not directly comparable to the passive loading in the present investigation, it was revealed that in the presence of greater MTU stiffness in males, there was no gender difference in EMG from the quadriceps or gastrocnemius (Granata et al. 2002; Padua et al. 2005, respectively). Therefore, the greater distal displacement and lower muscle stiffness observed in females in the present study likely reflects either a greater CSA in the males, or an intrinsically more compliant muscle, rather than lower EMG activity.
As CSA was not measured in the present study, its role can only be approximated using a surrogate measure of muscle mass such as MVC. In the measurement of tendon stiffness, heterogeneity in the strength of participants is often accounted for by presenting torque relative to MVC (Kubo et al. 2003; Onambele et al. 2007). When passive torque is normalised to MVC and presented as stiffness by distal displacement relative to muscle length, men have significantly greater stiffness than women. This would suggest that based on greater displacement of the GM MTJ, there is a difference in the viscoelastic properties of the GM muscle between genders.
Oestrogen is known to play a role in fibroblast proliferation and collagen synthesis in tendon (Kjaer and Hansen 2008). At present the possible role of oestrogen on muscle stiffness is ambiguous, with oestrogen replacement therapies providing evidence of a positive effect of oestrogen on muscle mass (Moran et al. 2007), and one would assume muscle stiffness. In contrast, the suppression of oestrogen through long-term oral contraceptive use has been shown to lead to increased stiffness in the Achilles tendon of humans in vivo (Bryant et al. 2008). It is possible that oestrogen, though mediating collagen content within the muscle and aponeurosis, could account for the findings of lower muscle stiffness in females in the present investigation.
Based on previous investigations on tendon stiffness under loading, it is tempting to calculate tendon force as a means of more accurately determining muscle stiffness. In the gastrocnemius, this has been previously based on the measurement of the Achilles tendon moment arm and the relative contribution of the GM to the calculated tendon force (Fukunaga et al. 1996). However, as the measured passive torque contains an appreciable contribution from frictional forces within the joint capsule, it is impossible to calculate the actual contribution made by the GM in the present study. In addition, although the moment arm was not measured in the present investigation, it is often derived from lower limb length when MRI is not available (Visser et al. 1990). Considering that the male MTU length was within 0.3 cm of the females, and the MTU length is equivalent to lower limb length, the scaling factor of moment arm length if estimated by limb length, as in Visser et al. (1990), is likely to be negligible. Interestingly, the finding that MTU length was not significantly different between males and females in the presence of a 13-cm greater stature in males is consistent with previous reports. For example, Kubo et al. (2003) reported a 3-cm difference in lower limb length between males and females in the presence of a 12-cm difference in stature.
The main findings from the present investigation provide in vivo evidence that GM muscle stiffness is lower in females than in males, which is likely to be due to a gender difference in the viscoelastic properties on the muscle. Secondary to this, the implications from the present investigation would suggest that some caution should be made when making conclusions of muscle stiffness from the passive torque–angle relation of the MTU as a whole. Indeed, in the present population there was no significant gender difference in passive torque of the MTU as a whole (based on the torque angle relation), which if measured in isolation, would have led to the erroneous conclusion that there was no gender difference in muscle stiffness. This is highlighted by previous studies that have based conclusions on the greater muscle stiffness in males, on a higher torque–angle relation on the MTU as a whole (Gajdosik et al. 1990). It is pertinent to reiterate therefore, that in the GM even though there was no difference in the torque–angle relation, in vivo passive stiffness was greater in males than in females.
References
Blackburn JT, Riemann BL, Padua DA, Guskiewicz KM (2004) Sex comparison of extensibility, passive, and active stiffness of the knee flexors. Clin Biomech 19:36–43
Bryant AL, Clark RA, Bartold S, Murphy A, Bennell KL, Hohmann E, Marshall-Gradisnik S, Payne C, Crossley KM (2008) Effects of estrogen on the mechanical behavior of the human Achilles tendon in vivo. J Appl Physiol 105:1035–1043
Fukunaga T, Roy RR, Shellock FG, Hodgson JA, Edgerton VR (1996) Specific tension of human plantar flexors and dorsiflexors. J Appl Physiol 80:158–165
Gajdosik RL, Giuliani CA, Bohannon RW (1990) Passive compliance and length of the hamstring muscles of healthy men and women. Clin Biomech 5:23–29
Granata KP, Wilson SE, Padua DA (2002) Gender differences in active musculoskeletal stiffness. Part I. Quantification in controlled measurements of knee joint dynamics. J Electromyogr Kinesiol 12:119–126
Kjaer M, Hansen M (2008) The mystery of female connective tissue. J Appl Physiol 105:1026–1027
Kubo K, Kanehisa H, Kawakami Y, Fukunaga T (2001) Influence of static stretching on viscoelastic properties of human tendon structures in vivo. J Appl Physiol 90:520–527
Kubo K, Kanehisa H, Fukunaga T (2003) Gender differences in the viscoelastic properties of tendon structures. Euro J Appl Physiol 88:520–526
Maganaris CN, Paul JP (1999) In vivo human tendon mechanical properties. J Physiol 521(Pt 1):307–313
Maganaris CN, Paul JP (2002) Tensile properties of the in vivo human gastrocnemius tendon. J Biomech 35:1639–1646
Magnusson SP, Simonsen EB, Aagaard P, Boesen J, Johannsen F, Kjaer M (1997) Determinants of musculoskeletal flexibility: viscoelastic properties, cross-sectional area, EMG and stretch tolerance. Scand J Med Sci Sport 7:195–202
Mahieu N, Witvrouw E, Stevens V, Willems T, Vandertraeten G, Cambier D (2004) Test-retest reliability of measuring the passive stiffness of the Achilles tendon using ultrasonography. Isok Exerc Sci 12:185–191
McHugh MP, Kremenic IJ, Fox MB, Gleim GW (1998) The role of mechanical and neural restraints to joint range of motion during passive stretch. Med Sci Sport Exerc 30:928–932
Moran AL, Nelson SA, Landisch RM, Warren GL, Lowe DA (2007) Estradiol replacement reverses ovariectomy-induced muscle contractile and myosin dysfunction in mature female mice. J Appl Physiol 102:1387–1393
Morse CI, Thom JM, Reeves ND, Birch KM, Narici MV (2005) In vivo physiological cross-sectional area and specific force are reduced in the gastrocnemius of elderly men. J Appl Physiol 99:1050–1055
Morse CI, Degens H, Seynnes OR, Maganaris CN, Jones DA (2008) The acute effect of stretching on the passive stiffness of the human gastrocnemius muscle tendon unit. J Physiol 586:97–106
Onambele GN, Burgess K, Pearson SJ (2007) Gender-specific in vivo measurement of the structural and mechanical properties of the human patellar tendon. J Orthop Res 25:1635–1642
Padua DA, Carcia CR, Arnold BL, Granata KP (2005) Gender differences in leg stiffness and stiffness recruitment strategy during two-legged hopping. J Motor Behav 37:111–125
Purslow PP (1989) Strain-induced reorientation of an intramuscular connective tissue network: implications for passive muscle elasticity. J Biomech 22:21–31
Taylor DC, Dalton JD Jr, Seaber AV, Garrett WE Jr (1990) Viscoelastic properties of muscle-tendon units. The biomechanical effects of stretching. Am J Sport Med 18:300–309
Visser JJ, Hoogkamer JE, Bobbert MF, Huijing PA (1990) Length and moment arm of human leg muscles as a function of knee and hip-joint angles. Euro J Appl Physiol 61:453–460
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Toshio Moritani.
Rights and permissions
About this article
Cite this article
Morse, C.I. Gender differences in the passive stiffness of the human gastrocnemius muscle during stretch. Eur J Appl Physiol 111, 2149–2154 (2011). https://doi.org/10.1007/s00421-011-1845-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-011-1845-z