
Abstract
We tested the hypothesis that menthol application would reduce the magnitude and initiation of sweating via excitation of cold-sensitive afferent pathways and concurrently via a cross-inhibition of heat loss pathways in acclimatized (swimmers, SW) and non acclimatized (control, CON) subjects in cool water. It was expected this effect to be exaggerated in SW subjects. Eight SW and eight CON subjects cycled at 60% of their \( \dot{V} \)O2max, as long as to reach 38°C in rectal temperature (Tre), without or with (4.6 g per 100 ml of water) all-body application of menthol sediment. Heart rate (HR), Tre, sweating rate (SwR), the proximal–distal skin temperature gradient (TSkf–f), and oxygen consumption (\( \dot{V} \)O2) were measured continuously. \( \dot{V} \)O2 and HR were similar between groups and conditions. Menthol increased TSkf–f, Tre threshold for SwR [+0.32 (0.01)°C] and Tre gain, while menthol reduced exercise time by 8.1 (4.1) min. SW group showed higher changes in Tre threshold for SwR [+0.50 (0.01)°C for SW vs. +0.13 (0.03)°C for CON], higher Tre gain, lower time for Tre increase and shorter exercise time [−10.7 (7) min for SW vs. −4.9 (4) min for CON] in menthol condition. Upon exercise initiation, previously applied menthol on the skin seems to induce vasoconstriction, results in a delayed sweating, which in turn affects the rectal temperature. Acclimatized subjects showed higher delay in SwR and earlier rise in Tre, which most probably is due to the inter-group differences in cold receptors activity.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Sweating rate (SwR) is regulated by central mechanisms through integration of mean skin, core temperature and other, non-thermal factors (Eiken and Mekjavic 2004; Mekjavic and Eiken 2006; Shibasaki et al. 2006). During exercise, rectal temperature (Tre) rises and heat dissipation occurs mainly through evaporation of sweat secreted from eccrine glands. On the contrary, stimulation of cold skin sensors induces thermogenesis aiming at body heat conservation and diminution of core temperature drop in a cold environment (Mekjavic and Eiken 2006). According to a long standing theory for reasons of thermoregulatory efficiency, stimulation of skin cold sensors is expected also to deactivate sweating by an input of reciprocal cross-inhibition (RCI) to heat loss center at preoptic anterior hypothalamus (Bazett 1949; Bligh 1998, 2006). Whereas this is a likely occurrence, this hypothesis has not been unequivocally validated in humans.
A novel approach for studying non-invasively the integrative mechanisms of thermal factors affecting SwR is the application of menthol on the skin. At rest, menthol is a stimulant of skin cold sensors (Hensel and Zotterman 1951; Schafer et al. 1986) provoking heat gain autonomic and behavioral responses in animals (Kamo et al. 2008; Tajino et al. 2007) and behavioral responses in humans (Green 1992). In particular, menthol can elevate core temperature by inducing vasoconstriction, non shivering thermogenesis and heat seeking behavior in mice (Tajino et al. 2007) and reduce the perception of warmth in humans (Green 1986, 1992). It is not known, however, whether stimulation of cold sensors through application of menthol, a widespread ingredient of various cosmetic and therapeutic skin creams, could impede sweating and, consequently, affect core temperature in humans during exercise.
Additionally, it has been suggested that repeated exposure to cool water for long periods, as it occurs in competitive pool swimming, could result in a deficiency of sweat gland function caused by de-acclimatization in conjunction with an increased activity of cold sensors in order to prevent heat loss and increase cold tolerance (McMurray and Horvath 1979). To our knowledge, there is no study that examines the effect of chronic cool water exposure on the function of cold sensors and the SwR response during exercise. Imposing menthol in swimmers and control subjects during steady state out of water exercise may provide further insight into local and integrative factors affecting SwR response.
The purpose of the present study was to investigate the effect of skin surface menthol application on sweating rate response during steady state exercise. It was hypothesized that menthol application, according to RCI model would excite cold sensors activity, trigger heat conservation mechanisms and lessen the gain and the threshold of sweating. The SwR response would be expected to be further suppressed in subjects exposed regularly to cool water, mainly due to a diminished sensitivity of skin cold sensors.
Methods
Subjects
Sixteen male healthy subjects, eight physical education students (CON) and eight swimmers (SW) volunteered to participate in this study. Swimmers were participating in national level events and they were training in the swimming pool four times per week. Physical education students were physically active and never systematically exposed to cool or cold water. All the procedures had the approval of the local Ethical Committee.
Study outline
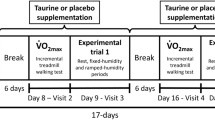
Subjects were initially tested for a possible menthol allergic reaction by applying the substance on a small skin area. On a separate day, a \( \dot{V} \)O2max test was conducted in a cycle ergometer. In a subsequent trial, at least 2 days apart, subjects cycled in a random and counterbalanced order with or without menthol application at ~60% of their maximal oxygen uptake, so long as the rectal temperature of the subject to reach 38°C. All experiments were conducted in the springtime at the same time of the day, in a controlled environmental temperature [24.1 (1.0)°C] and relative humidity [46.2 (4.3)%].
Procedure
\( \dot{V} \)O2max
Each subject performed an incremental protocol on a cycle ergometer (Lode, Groningen, The Netherlands) with increments of 30 W per minute to exhaustion. Resting values were measured for a 5-min period preceding the exercise bout. The initial workload was 60 W and the target pedaling rate 70–80 rpm. The criteria to achieve maximal oxygen uptake were: respiratory exchange ratio >1.1, plateau in \( \dot{V} \)O2 for the last 15 s of the trial and inability to maintain the required cadence despite the strong verbal encouragement provided by the prime researcher.
Steady state exercise
For 2 days prior to each main trial, subjects were asked to keep similar habits before each experiment. Special instructions were given to the subjects for consuming adequate amounts of water three days before the experiment. Additionally, they had to abstain from physical activity, caffeine and alcohol consumption the day before the trial. On the experimental day, after the subjects emptied their bladder, their body mass was measured (Bilance Salus, Milano, Italy) and a thermistor was placed in the rectum, at a depth of 13–15 cm from the sphincter. While subject was seated and relaxed on the bicycle, thermistors on forearm and fingertip were placed and secured with an adhesive tape. Following this preparation, 5-min resting values for HR, \( \dot{V} \)O2 and temperatures were obtained. Afterwards, each subject cycled so long as to reach 38°C in Tre. In menthol condition, 5-min resting period without menthol was followed by application of menthol creamy sediment on the skin with a cotton pellet all over the body except the area (~7 cm2) on the forearm where the sweat capsule was positioned. The menthol creamy sediment contained 4.6 g of the substance (2-Isopropyl-5-methylcyclohexanol) in needlelike crystals being dissolved to 100 ml of warm water. Subsequently, menthol-resting data were collected for five more minutes.
Analytical methods and equipment
Oxygen uptake was measured continuously with a metabolic cart (MedGraphics, CPX-D, USA), which was calibrated with two different gas mixtures before each testing. Minute mass flow of local secreted sweat rate of the forearm was measured incessantly during the experiment at a frequency of 200 Hz, by means of a ventilated capsule positioned on a 5.72-cm2 area of skin of the right forearm, and ventilated at a rate of 2 l min−1 with air at room temperature. The temperature and humidity of air entering and exiting the capsule were measured with thermocouples (TSD 202A, Biopac, USA) and capacitance hygrometers (Delta, UK), respectively. Sweating rate (SwR) was estimated from the difference between the temperature and the humidity of inflowing and out flowing air. Temperature and humidity sensors were appropriately calibrated before their use. Heart rate was measured telemetrically every 5 s (Polar S-810, Finland). Rectal temperature (Tre) and skin (Tsk) temperatures of the forearm (T for) and fingertip (T fing) were recorded every minute with thermistors (Yellow Springs, USA) connected to a tele-thermometer (Yellow Springs, Model 46, USA). The difference in temperature between forearm and fingertip (TSkf–f) was considered as an index of vasomotor tone with the higher temperature difference indicating higher vasoconstriction (Rubinstein and Sessler 1990; House and Tipton 2002). Chest, thigh, triceps, subscapular, suprailiac, abdominal and axillary skinfolds were taken for the calculation of body fat. Body fat percentage was calculated with the equation of Jackson and Pollock (1978).
Data analysis
A two-way analysis of variance for independent (students and swimmers) and dependent (control and menthol) samples was used to define the overall differences in each variable. Where needed time was used as one more repeated measures factor for the analysis of variance. A Tukey test was employed to assign post hoc-specific differences. Deviation from linearity using the least squares linear regression method was used to determine the rectal temperature threshold for the initiation of sweating and the exercise time for the initiation of rise in Tre. The reported values are averages of the individual thresholds and slopes derived from linear regression parameters. Data are presented as mean ± SD unless indicated otherwise. Significance level was set at 0.05.
Results
The subjects’ age, anthropometric characteristics and maximal oxygen uptake are presented in Table 1. Both groups, CON and SW, had similar age, stature and body mass. However, SW tended to have higher body fat (p < 0.07). Moreover, the latter group had higher (p < 0.05) maximal oxygen uptake values (Table 1). All subjects tolerated menthol well on their skin without any adverse reaction.
Steady state exercise
Oxygen uptake and heart rate
Both groups cycled with or without menthol application on their skin at the same relative intensity. Although \( \dot{V} \)O2 was higher by ~220 ml min−1 in SW than in CON, the analysis of variance did not revealed a significant difference (p = 0.11, Table 2). No significant differences were also detected between the two experimental conditions, with and without menthol application, in both groups (Table 2). In addition, \( \dot{V} \)O2 was similar when data were expressed according to Tre changes. HR was similar during cycling with and without menthol, both in CON and SW (Table 2).
Rectal temperature and exercise time
Resting values of Tre were indifferent between or within groups (Table 2). In addition, no significant difference was found when Tre values were compared at the end of the fifth minute of resting with menthol application, with the Tre values in the first 5 min of resting without menthol application in the same condition. More specifically, the mean Tre was 37.4 (0.3) and 37.1 (0.4)°C for the SW and 37.3 (0.4) and 37.1 (0.3)°C for CON without or with menthol application, respectively. The rate of increase in Tre during exercise with menthol application was higher (p < 0.05) compared with the exercise without menthol both in SW and CON group (Table 2). In both groups, core temperature reached at 38°C sooner in the menthol trials (Table 2). Moreover, the initiation of Tre rise exhibited earlier (p < 0.05) in the SW compared with CON, in menthol condition (Table 2). In particular, Tre threshold appeared 2.5 (1.1) min earlier (p < 0.05) in SW menthol condition compared with the CON menthol condition. Menthol application reduced the exercise time in CON group by ~5 min while in the SW group by ~11 min.
Forearm-fingertip temperature difference (Τskf–f) and sweating rate (SwR)
Τskf–f was higher (p < 0.05) for the first 11 min of exercise in SW and CON with menthol skin application (Fig. 1). Afterwards, Τskf–f was similar in M and NM in both groups of subjects. In addition, no significant differences were observed in the resting forearm skin temperature between groups or conditions. Indeed, the mean forearm skin temperature was 32.4 (0.6), 32.9 (0.4), 33.0 (1.0) and 32.7 (0.5) for the SW and CON, in M and NM condition, respectively. The respective mean values during exercise were 34.5 (0.2), 34.9 (0.3), 34.2 (0.3) and 33.9 (0.6) without significant differences between conditions.

Temperature difference between forearm and fingertip (Tskf–f) at rest (R) and during exercise for swimmers (SW, n = 8) and physical education students (CON, n = 8) with (M) and without (NM) menthol application. Values are means; standard deviation was removed for the clarity of the presentation. *Significant differences between M and NM (p < 0.05)
The SwR response in CON and SW group with and without menthol is presented in Fig. 2. In both groups, menthol application postponed SwR initiation; it started later (p < 0.01) at a higher (p < 0.01) Tre (Fig. 2, Table 2). Additionally, SwR threshold, in terms of time and core temperature, was significantly higher (p < 0.01) in SW compared with CON in the menthol condition (Table 2). The SwR increase for a given Tre change was smaller (p < 0.01) in both groups when menthol was applied on the skin compared with the condition without menthol application (Fig. 2, Table 2).

Sweating rate response (SwR) plotted against change in rectal temperature (ΔΤre) for swimmers (SW; lower graph, n = 8) and physical education students (CON; upper graph, n = 8) with (M) and without (NM) menthol application. Tsk values are similar among experimental conditions. The values are means (SE). *Significant differences between M and NM (p < 0.05)
Discussion
In the present study, menthol application on the skin induced vasoconstriction and delayed the initiation of sweating, resulting in a faster rise of Tre. After menthol appliance, swimmers showed similar vasoconstriction as control subjects, but a greater delay in sweating, leading to an earlier rise in rectal temperature.
It is well documented that menthol is a cooling-mimetic chemical that produces in a thermoneutral environment the sensation of cooling, without reducing the skin temperature (Hensel and Zotterman 1951). Furthermore, menthol causes a profound increase in depolarisation of the cold-activated neurons and shifts the threshold temperature of firing significantly towards warmer temperature which appears to be close to normal skin temperature (from 31.5 to 34–37°C; Linte et al. 2007; Reid et al. 2002). Thus, menthol initiates cooling-induced autonomic and behavioral heat gain responses (Tajino et al. 2007). In resting rats, menthol raised \( \dot{V} \)O2, induced vasoconstriction and elevated Tre, 30 min after its application on the skin (Tajino et al. 2007). As we hypothesized, similar responses were observed in the present study, which was conducted in humans exercising at a submaximal intensity. Indeed, application of menthol resulted in severe skin vasoconstriction, and a slight increase in oxygen consumption. Both of these responses led Tre to an earlier [by 3.5 (1) min; p < 0.05] and steeper rise (p < 0.05, Table 2). Therefore, time to reach Tre at 38°C in the menthol compared to the non-menthol condition was reduced by 7.9 (2.1) min. After the first 11 min of exercise, the vasoconstrictive effect of menthol was released, probably due to elevation of Tre by 0.35 (0.08)°C. The \( \dot{V} \)O2 increased slightly, but not significantly, after menthol application. The observed differences with the study of Tajino et al. (2007) in terms of oxygen uptake response could be accredited to the employed methodology, namely due to the utilization of exercise in the present study. The increments of \( \dot{V} \)O2 after menthol application could be masked by the small difference in the intensity of exercise performed in the two experimental conditions. Furthermore, the effect of menthol on oxygen uptake might be small when oxygen requirements due to exercise are high and the sample size probably not adequate to show significant differences between conditions. Unfortunately, in the present study resting values were not collected for a long time with and without menthol application, to verify the results of Tajino et al. (2007).
Whereas an increase of 0.35 (0.1)°C in the Tre appeared to release the vasoconstrictive effect of menthol on the skin, sweating was not enhanced by this Tre increase, despite the fact that sweating is primarily initiated by a core temperature rise and central warm reception (Mekjavic and Eiken 2006). On the contrary, menthol application on the skin appeared to delay the initiation and reduce the gain of sweating dramatically both in swimmers and nonswimmers. This effect could not be attributed to a local effect of menthol since the skin area under investigation in both experimental conditions was free of menthol and no significant differences were observed in the forearm skin temperature between groups or conditions. It is likely that menthol modified the firing characteristics of skin cold sensors and exerted its effects on heat dissipation effector, namely sweating, centrally. It has been suggested originally by Bazett (1949) that nerve fibers carrying thermo-afferent information from the skin sensors to heat production locus of the preoptic posterior hypothalamus might have collaterals innervating the heat dissipation locus of the preoptic anterior hypothalamus. Thus, an excitatory stimulus on skin cold sensors might lead to an inhibitory input in sweating (Bligh 2006). Neuroanatomical and physiological evidence certainly supports the existence of such RCI concept; the significance of this interaction at the systemic level is not obvious (Mekjavic and Eiken 2006). The present study shows that RCI of skin cold afferent information, as in the case of cold ambient exposure, suppresses sweating which on one hand contributes to core temperature increase and on the other to conservation of body fluids and maintain of cardiovascular stability. Sweating is initiated by enhanced activity of central warm sensitive elements evoked in the present experiments by the thermogenic effect of exercise. Therefore, the widening of the sweating threshold zone in the menthol condition would imply that the applied substance opposes the exercise-induced sweating. This would take place until the excitatory thermogenic effect of exercise on sweating would prevail over the menthol inhibitory effect. This opposition effect of menthol on sweating appears to persist in a constant way even after its initiation since sweating gain was similarly reduced in both the swimmers and the control subjects.
Whereas sweating gain was not different between swimmers and controls, the former group of subjects when compared with the latter initiated sweating at a smaller rectal temperature increase. Habituation responses of the thermoregulatory system to cold have been reported by other studies (Golden and Tipton 1988; Leppaluoto et al. 2001) but to our knowledge, this is the first time that the control of SwR after prolonged periods of skin exposure in cool water is investigated. According to Reid et al. (2002), adaptation of cold sensors to cooling shifts its threshold of depolarization to a lower temperature. So a stronger cooling stimulus is required to activate the sensor. Regarding the present study, we could assume that the repeated exposure of swimmers to 24°C water desensitized skin cold sensors at 33°C, which was the recorded forearm skin temperature, during exercise. A putative mechanism to explain this finding is that RCI in swimmers when compared with controls was weaker and initiation of sweating occurred at lower rectal temperature. Noteworthy, one study (McMurray and Horvath 1979) reported lower total sweat losses in swimmers compared with runners when exercised in warm water (35°C).
Menthol application induced wider shift to the right in the sweating threshold zone in SW compared with CON. This prolonged lateness in sweating initiation occurred without any difference, between groups, in the gain of SwR and the vasoconstriction response, as expressed by Τskf–f. This suggests that the delay of sweating initiation is almost the exclusive cause of the higher gain in Tre being recorded in menthol condition between SW and CON. The initiation of sweating in SW is exhibited later compared to CON probably due to a synergistic effect of cold sensors adaptation and menthol application. Adaptation to cold shifts the threshold across the temperature axis to the left, whereas menthol application shifts the threshold to the right, namely to warmer temperatures. Adaptation shifts the temperature sensitivity but the sensitivity of the sensor channel to increment of stimulus is preserved (Reid et al. 2002). Therefore, application of menthol on the skin of swimmers was perceived as a stronger stimulus compared to controls since cold sensors were silent due to cooling acclimatization in the former group of subjects. Thus, in swimmers compared to non-swimmers, skin cold sensors are more receptive to menthol and the forthcoming RCI to sweating stronger. Once initiated, sweating gains are the same indicating that this response is dependent on peripheral factors. With the data available, lower central warm sensitivity, in SW than in CON, could not be excluded as a factor explaining the SwR threshold differences, but further research is needed.
Apart from thermal, other non-thermal factors can influence SwR during exercise (Eiken and Mekjavic 2004; Kacin et al. 2007; Mekjavic and Eiken 2006). Body fat percentage and fitness level could influence SwR in the present study. One of the characteristics of the acclimatized subjects exposed for prolonged periods in cool water is the increased percentage of body fat for insulation reasons (McMurray and Horvath 1979; Yoshida et al. 1998). In the present study, the differences in body fat between groups although small (about 3%), tended to be significant (p = 0.07), a fact that could potentially affect the regulation of Tre course in swimmers and potentially enhance sweating. In addition, SW group in comparison with control subjects had significantly higher maximal oxygen uptake. The differences in \( \dot{V} \)O2max have also been suggested that could enhance SwR (Buono and Sjoholm 1988; Yamazaki et al. 1994). In the present study, there was a faster initiation of SwR in the SW compared with CON group in the condition without menthol application, which is in line with the above studies. However, the magnitude of the SwR in the SW was not higher as could be expected. This dissimilarity could be attributed to the fact that \( \dot{V} \)O2max difference in the present study was ~5 mL kg−1 min−1, while in the previous studies was 15–20 mL kg−1 min−1. Menthol application compromises sweating and induces vasoconstriction in the skin, which is presumably accompanied by central blood pooling. Thoracic blood pooling has been also proven to reduce sweating (Geladas 1990). The influence of these non-thermal cardiovascular factors on our results cannot be excluded and further research is apparently needed. No placebo group was included in the present experimental design since it was not possible to duplicate the menthol effect on the skin without affecting thermoregulation. In addition to that, a second control group should have been used and had 100 ml of water, without menthol, applied to the skin. This group would have nullified the likelihood that evaporation of the water contained in the menthol solution would have per se caused skin cooling. Most possibly however this is not the case, due to similar initial skin and core temperatures and similar skin temperatures and different SwR responses observed during exercise among conditions. It is likely that the rate of evaporation of the menthol sediment was limited due to its high density and inter-molecular forces developed between the water and the menthol matter dissolved in it. Furthermore, part of the probable heat loss due to the evaporation of water contained in the menthol sediment would be counteracted by the heat transfer of the warm water to the skin.
Additionally, the maximal oxygen uptake was tested in the cycle ergometer, a fact that could underestimate the real maximal values for the SW, as this group has reduced efficiency in out water testing. This is unlikely that could affect our results because subjects cycled at similar relative workload according to their maximal values obtained from the same ergometer. Lastly, the menthol effect seems to be dose-dependent (Tajino et al. 2007). Consequently the effects of different doses of menthol application to the human’s skin still need to be further investigated.
Taken collectively, our findings suggest that during steady state exercise, previously applied menthol on the skin induces vasoconstriction, delays initiation of sweating and reduces the sweating gain, enhancing the rectal temperature rise. Acclimatized subjects to cool water showed a further shift of sweating threshold to higher rectal temperature and an earlier rise in Tre, which is most probably due to the inter-group differences in cold sensors activity.
References
Bazett HC (1949) The regulation of body temperature. In: Newburgh LH (ed) Physiology of heat regulation and the science of clothing. Saunders, Philadelphia, pp 109–192
Bligh J (1998) Mammalian homeothermy: an integrative thesis. J Therm Biol 23:143–258
Bligh J (2006) A theoretical consideration of the means whereby the mammalian core temperature is defended at a null zone. J Appl Physiol 100:1332–1337
Buono MJ, Sjoholm NT (1988) Effect of physical training on peripheral sweat production. J Appl Physiol 65:811–814
Eiken O, Mekjavic IB (2004) Ischaemia in working muscles potentiates the exercise-induced sweating response in man. Acta Physiol Scand 181:305–311
Geladas ND (1990) Effects of posture and static exercise on heat dissipating mechanisms in humans. PhD dissertation, Simon Fraser University, Canada. http://ir.lib.sfu.ca/dspace/bitstrean/1892/6395/1/b14677453.pdf
Golden FS, Tipton MJ (1988) Human adaptation to repeated cold immersions. J Physiol 396:349–363
Green BG (1986) Menthol inhibits the perception of warmth. Physiol Behav 38:833–838
Green BG (1992) The sensory effects of l-menthol on human skin. Somatosens Mot Res 9:235–244
Hensel H, Zotterman Y (1951) The effect of menthol on thermoreceptors. Acta Physiol Scand 24:27–34
House JR, Tipton MJ (2002) Using skin temperature gradients or skin heat flux measurements to determine thresholds of vasoconstriction and vasodilatation. Eur J Appl Physiol 88:141–145
Jackson AS, Pollock ML (1978) Generalized equations for predicting body density of men. Br J Nutr 40:497–504
Kacin A, Golja P, Eiken O, Tipton MJ, Mekjavic IB (2007) The influence of acute and 23 days of intermittent hypoxic exposures on the exercise-induced forehead sweating response. Eur J Appl Physiol 99:557–566
Kamo H, Honda K, Kitagawa J, Tsuboi Y, Kondo M, Taira M, Yamashita A, Katsuyama N, Masuda Y, Kato T, Iwata K (2008) Topical capsaicin application causes cold hypersensitivity in awake monkeys. J Oral Sci 50:175–179
Leppaluoto J, Korhonen I, Hassi J (2001) Habituation of thermal sensations, skin temperatures, and norepinephrine in men exposed to cold air. J Appl Physiol 90:1211–1218
Linte RM, Ciobanu C, Reid G, Babes A (2007) Desensitization of cold- and menthol-sensitive rat dorsal root ganglion neurones by inflammatory mediators. Exp Brain Res 178:89–98
McMurray RG, Horvath SM (1979) Thermoregulation in swimmers and runners. J Appl Physiol 46:1086–1092
Mekjavic IB, Eiken O (2006) Contribution of thermal and nonthermal factors to the regulation of body temperature in humans. J Appl Physiol 100:2065–2072
Reid G, Babes A, Pluteanu F (2002) A cold- and menthol-activated current in rat dorsal root ganglion neurones: properties and role in cold transduction. J Physiol 545:595–614
Rubinstein EH, Sessler DI (1990) Skin-surface temperature gradients correlate with fingertip blood flow in humans. Anesthesiol 73:541–545
Schafer K, Braun HA, Isenberg C (1986) Effect of menthol on cold receptor activity. Analysis of receptor processes. J Gen Physiol 88:757–776
Shibasaki M, Wilson TE, Crandall CG (2006) Neural control and mechanisms of eccrine sweating during heat stress and exercise. J Appl Physiol 100:1692–1701
Tajino K, Matsumura K, Kosada K, Shibakusa T, Inoue K, Fushiki T, Hosokawa H, Kobayashi S (2007) Application of menthol to the skin of whole trunk in mice induces autonomic and behavioral heat-gain responses. Am J Physiol Regul Integr Comp Physiol 293:R2128–R2135
Yamazaki F, Fujii N, Sone R, Ikegami H (1994) Mechanisms of potentiation in sweating induced by long-term physical training. Eur J Appl Physiol 69:228–232
Yoshida T, Nagashima K, Nakai S, Yorimoto A, Kawabata T, Morimoto T (1998) Nonshivering thermoregulatory responses in trained athletes: effects of physical fitness and body fat. Jpn J Physiol 48:143–148
Acknowledgments
We are indebted to the subjects for their enthusiastic and consistent participation for the completion of the present study. The authors also express their appreciation to Prof Igor B. Mekjavic for his helpful comments in the preparation of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by George Havenith.
Rights and permissions
About this article
Cite this article
Kounalakis, S.N., Botonis, P.G., Koskolou, M.D. et al. The effect of menthol application to the skin on sweating rate response during exercise in swimmers and controls. Eur J Appl Physiol 109, 183–189 (2010). https://doi.org/10.1007/s00421-009-1345-6
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-009-1345-6