
Abstract
Muscle mass and strength have been shown to be important factors in bone strength. Low muscular force predisposes to falling especially among elderly. Regular exercise helps to prevent falls and resulting bone fractures. Better understanding of muscle function and its importance on bone properties may thus add information to fracture prevention. Therefore the purpose of this study was to examine the relationship between bone strength and muscular force production. Twenty-young men [24 (2) years] and 20 [24 (3) years] women served as subjects. Bone compressive (BSId) and bending strength indices (50 Imax) were measured with peripheral quantitative computed tomography (pQCT) at tibial mid-shaft and at distal tibia. Ankle plantarflexor muscle volume (MV) was estimated from muscle thickness measured with ultrasonography. Neuromuscular performance was evaluated from the measurements of maximal ground reaction force (GRF) in bilateral jumping and of eccentric maximal voluntary ankle plantarflexor torque (MVC). Specific tension (ST) of the plantarflexors was calculated by dividing the MVC with the muscle volume. Activation level (AL) was measured with superimposed twitch method. Distal tibia BSId and tibial mid-shaft 50 Imax correlated positively with GRF, MVC and MV in men (r = 0.45–0.67, P < 0.05). Tibial mid-shaft 50 Imax and neuromuscular performance variables were correlated in women (r = 0.46–0.59, P < 0.05), whereas no correlation was seen in distal tibia. In the regression analysis, MV and ST could explain 64% of the variance in tibial mid-shaft bone strength and 41% of the variation in distal tibia bone strength. The study emphasizes that tibial strength is related to maximal neuromuscular performance. In addition, tibial mid-shaft seems to be more dependent on the neuromuscular performance, than distal tibia. In young adults, the association between bone adaptation and neuromuscular performance seems to be moderate and also site and loading specific.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
It has been suggested that low muscular strength predisposes to falling (Moreland et al. 2004) and ensuing bone fractures (Stevens and Olson 2000). Even though pharmacological interventions in preventing fractures seem effective, the cost of treating all those in need is too high, thus making life style interventions, like exercise, all the more intriguing (Cummings and Melton 2002). In fact, high-impact exercise intervention has been shown to be successful in increasing not only bone mineral density (BMD) but also improving dynamic balance (Heinonen et al. 1996), thus decreasing fall rate (Gass and Dawson-Hughes 2006). Preventing fractures with exercise requires physical activity maintained throughout the elderly years. If muscular loading is not maintained, the age related bone loss cannot be prevented (Schonau 2004). High impact and unusual loading, odd impact such as racket sports, exercises seems beneficial for bone (Nikander et al. 2005, 2006). It seems that impacts (Heinonen et al. 1996) and bending are the most important loading modalities in increasing bone mass (Heinonen et al. 2002).
Muscle mass (Calbet et al. 1999; Pettersson et al. 1999; Ribom et al. 2004; Taaffe and Marcus 2004), strength (Blain et al. 2001; Calmels et al. 1995; Nordstrom et al. 1996; Pettersson et al. 2000; Sandstrom et al. 2000) and high physical activity have been shown to be associated with high areal bone mineral density (aBMD) in the loaded sites (Nordstrom et al. 1995, 1996; Pettersson et al. 2000; Sandstrom et al. 2000). Thus, it has been suggested that the bone strength is dependent on loading caused by muscles (Burr 1997; Ferretti et al. 2003; Frost 2000). Muscle force in turn is related to physiological cross-sectional area of the muscle, the specific tension of the muscle (Fukunaga et al. 2001) and level of neural activation (Kent-Braun and Le Blanc 1996).
The adaptive mechanisms of bone are not likely to affect the bone material properties greatly, so it seems that the most important adaptive mechanism is remodeling, which regulates bone architecture (Ferretti et al. 2003; Heaney 2003). Indeed, the variance of aBMD explained by muscle strength seems to be only 10–20% (Taaffe et al. 1995) although it has been hypothesized that muscular loading should determine half of the postnatal bone strength development (Frost 2000).
In previous studies bone loading has been characterized indirectly with measurements of muscle volume (Ducher et al. 2005), muscle mass (Calbet et al. 1999), muscle cross-sectional area (Heinonen et al. 2001) or force (Pettersson et al. 1999). However, the muscle–bone interplay is not completely determined either by just muscle size or just by the neuromuscular output revealed by force measurements (Suominen 2006). Force measurements of neuromuscular performance can be further refined with variables such as activation level (Kent-Braun and Le Blanc 1996) and specific tension (Fukunaga et al. 2001).
To avoid the effects of aging on muscle–bone interplay, the purpose of this study was to examine the relationship between neuromuscular system and bone strength in healthy young adults. The hypothesis was that neuromuscular output has explanatory effect on bone independent of body size.
Methods
Subjects
Twenty healthy physically active young men (24.4 (2.4) years) and 20 women (24.0 (3.2) years) served as subjects (Table 1). None of the subjects used medications known to affect bone metabolism. The subjects represented a wide range of physical activity levels ranging from sedentary to amateur athletes. Team sports as well as individual events were represented in the subject group. The subjects were asked to restrain from rigorous physical activity for two days prior to the measurement day. The studies were conducted according to the declaration of Helsinki with the approval of the ethics committee of the University of Jyväskylä. The subjects gave their informed written consent to participate.
Bone structural characteristics
The peripheral quantitative computed tomography (pQCT), (XCT 2000, Stratec Medizintechnik GmbH, Pforzheim, Germany) was performed at the distal tibia (trabecular bone) and at the mid-shaft (cortical bone) of the tibia. The scans of tibia were performed at 5% (distal tibia) and 50% (tibial shaft) of the measured tibial length proximal to the distal end plate. The site of measurement was determined by measuring the length of tibia from anatomical landmarks (from knee joint line to medial malleolus). The bone structural characteristics were assessed with volumetric total bone mineral density (ToD, mg/cm³), cortical maximal moment of inertia (I, mg cm) and total cross-sectional area of bone (ToA, mm²). The bone strength and geometry analysis were conducted with BonAlyse Geanie (BonAlyse Oy, Jyväskylä, Finland) analysis program. The root mean square coefficients of variation (CVRMS) ranged from 0.4 to 1.6% for the bone variables determined.
Overall estimated bone strength was represented by the cortical maximal moment of inertia at the tibial mid-shaft (50 Imax) and by ToD squared multiplied by ToA (compressive bone strength index, BSId = ToD2 × ToA) at distal tibia. Cortical maximal moment of inertia reflects bone’s resistance to bending in the plane of maximal bending strength. Compressive bone strength index represents the compressive strength of bone.
Evaluation of neuromuscular performance
Ground reaction force measurement
Maximal vertical ground reaction force (GRF) was measured during bilateral jumping. Subjects were instructed to perform bilateral jumping on the soles of feet with plantarflexor muscles, keeping the hip and knees extended, on a force platform (University of Jyväskylä, Finland). Subjects were asked to begin jumping with low intensity and build up to maximal jumping height with 10–15 jumps. Knee and hip angle were controlled visually during jumping and the subjects were continuously given verbal instructions to jump with extended knees and hip. Knee flexion was checked on a video after jumping and was seen to be less than 10° on the average. The highest vertical ground reaction force was selected for analysis. CVRMS was 6.8% for maximal vertical GRF measurements.
Torque measurement
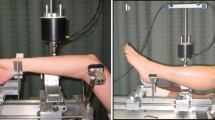
The maximum isometric and eccentric muscle torque was measured with custom-made dynamometer (Nicol et al. 1996) during unilateral ankle plantarflexion with knee extended. The subject was sitting comfortably with the upper body in an angle of 110° compared to the legs. The legs were parallel with the floor. Isometric torque was measured at various angles with 5° increments beginning approximately from 80° between the sole of the foot and tibia and finishing approximately at 105°. Torque angle relationship was determined from the isometric maximal voluntary contractions (MVC). The subjects were exhorted verbally to ensure maximal effort. The angle of the highest isometric torque value was selected as the optimal angle. Eccentric contractions were measured isokinetically at an angular velocity of 20°/s. The angular displacement started from 115°. The eccentric torque was measured at an optimal angle. The CVRMS ranged from 5.6 to 10.2% for the maximal voluntary torque measurements.
Muscle volume estimation
The volume of the ankle plantar–flexor muscle group was predicted from muscle thickness and limb length (Miyatani et al. 2004). The limb length was determined as the length between knee joint line at the lateral side and the lateral malleolus of tibia.
Muscle thickness was obtained from a cross-sectional image of the ankle plantar flexor muscle group with ultrasonographic measurement device (Prosound SSD-5500, Aloka, Tokyo, Japan). The thickest part of the muscle was used in the determination of muscle thickness. Muscle thickness was measured online from freezed ultrasound picture. CVRMS was 10% for the volume estimation. The volume of the plantar flexors was estimated from the thickness and limb length as follows (Miyatani et al. 2004):
Specific tension estimation
Relative specific tension was estimated from the ankle plantar flexor muscles. The specific tension was calculated as maximum voluntary torque produced, divided by muscle volume as suggested by Fukunaga et al. (2001):
where TQ is the torque, MV the muscle volume, ST the specific tension, MA the moment arm, FL the fiber length and θ the pennation angle. If the ration of MA to FL is assumed to be constant among subjects and the effect of pennation angle changes on muscle force is assumed to be negligible (Fukunaga et al. 2001), it follows that (Lynch et al. 1999)
Voluntary activation measurement
Voluntary activation was measured during eccentric maximal voluntary contraction from the ankle plantar flexor muscles using superimposed twitch method (Kent-Braun and Le Blanc 1996; Merton 1954). The level of activation was calculated with the activation level (AL) equation (Babault et al. 2001).
The AL was measured at an angle corresponding to optimal angle in isometric contraction during maximal eccentric ankle plantar flexor actions. The CVRMS for the activation level measurement was 4.2%
In the eccentric contraction, the torque level with stimulation was defined as the torque during maximal positive difference between the measured torque curve and post-stimulus torque line estimated with linear extrapolation (Allen et al. 1998). The twitch force was defined as the difference between the measured torque and extrapolated torque (Babault et al. 2001).
Electrical stimulation
Electrical stimulation was conducted to tibial nerve with the stimulation electrode placed over the tibial nerve in the popliteal fossa and the anode (Vtrode neurostimulation electrodes 2 × 4 in. oval electrode, Mettler electronics, Anaheim, CA, USA) placed below the patella. The placement of the stimulation electrode was controlled by visually inspecting the twitch response to single stimuli. The placement of the stimulating electrode was accepted when the twitch response was evenly distributed in plantar flexors and minimal twitch response was observed in the tibialis anterior muscle. Once the stimulation site was established with a reusable stimulating electrode, a disposable electrode (Unilect short-term Ag/AgCl ECG electrode) was placed on the established stimulation site. Sufficient pressure of the stimulation electrode was applied manually.
A 50 ms submaximal stimulus train at 100 Hz frequency (=6 stimuli) was applied with electrical stimulator (Digitimer constant current stimulator model DS7A, Digitimer, Welwyn garden city, England). The intensity of stimulation was adjusted to induce in relaxed muscle a twitch response torque of 30–40% of previously measured isometric MVC torque. The duration of the current pulse was 200 μs and the maximum voltage was 200 V. In the eccentric activity, the timing of the stimulus was adjusted so that maximal evoked twitch torque occurred at the time corresponding to optimal angle. The advance of the stimulus train application was determined from the twitch evoked to active muscle and was the time from the stimulus trigger to the peak of the evoked force. In practice the stimulus train was applied 50 ms prior to ankle angle reaching the optimum angle.
Neuromuscular measurement protocol
First, the anthropometric measurements were conducted, e.g. limb length, muscle thickness and bone strength measurements. Subjects were asked for self reported height and physical activity in terms of times/week. The dynamometer used for torque measurements was adjusted for the subject. The subjects were allowed to familiarize and warm up with the dynamometer device used for torque measurements for 5–10 submaximal isometric and eccentric actions.
The isometric angle–torque relationship was determined in pseudo-randomized order. Stimulation intensity was determined and stimulation trains were applied to familiarize the subject to the stimuli. The eccentric activation level was measured. All of the measurements were conducted with right leg. Finally the maximal ground reaction force was measured in bilateral jumping with knees and hip extended. Ten to 15 jumps were conducted. The subjects were informed that the rest periods should allow for full metabolic recovery and prevent significant neuromuscular fatigue from developing during the measurements.
Analysis
The torque curve was averaged using 50 ms moving average. In eccentric situation MVC was determined with post-stimulus extrapolation method (Allen et al. 1998). The eccentric MVC was defined as a value estimated with linear extrapolation from the last 25–225 ms of torque data prior to stimulus triggering. The length of the data used for linear extrapolation was determined by taking the linear estimate, which deviated the least from the measured torque value at the time of stimulus triggering. The eccentric MVC was calculated as the value of the linear estimate at the time of occurrence of maximal twitch torque increment.
Statistical analysis
Descriptive data were reported for variables of interest (mean and range). Associations between the independent variables or predictors (maximal ground reaction force, torque, specific tension, activation level) and dependent variables or outcomes (bone geometry variables) were determined by a Pearson product moment correlation. Normal distribution was analyzed with Shapiro-Wilk normality test. For women normality was violated and logarithmic correction was used to normalize the data distribution. When data from men and women was pooled, normality was violated and a logarithmic normalization was applied.
Regression models were developed with muscle volume, specific tension and activation level, as the independent variables. The selected independent variables are associated with neuromuscular output (Fukunaga et al. 2001). Regression analysis was run twice. In the first model height and weight were also included as independent variables. Height and weight were excluded from the model because of the significant co-variation with muscle volume. The dependent variables were distal tibia BSId and tibial mid-shaft 50 Imax. The model was forced model and the independent variables were entered in the order mentioned above. Correlation analyses were run for both sexes independently and with both genders pooled. The results of men and women were pooled for the regression analysis, since specific tension and activation level were not different between men and women. The results are reported as mean (standard deviation). The significance limit was set at P ≤ 0.05 for all statistical analyses. Statistical analyses were conducted with SPSS 13.0.1 (SPSS Inc.) statistical analysis program.
Results
Absolute values of neuromuscular and bone geometry variables are given in Table 2. Men had higher values than women except in specific tension and activation level. In addition, body weight adjusted to the maximal ground reaction force during jumping did not differ between the genders, being 6.2 (1.4) times body weight for men and 5.9 (0.8) for women.
The neuromuscular variables (GRF, maximal eccentric torque and plantar flexor muscle volume) correlated positively in both genders pooled with distal tibia BSId (r² = 0.34–0.37, P < 0.05) and with tibial mid-shaft 50 Imax (r² = 0.50–0.59, P < 0.05). In men GRF, maximal eccentric torque and plantar flexor muscle volume were positively correlated with the distal tibia and tibial mid-shaft bone strength indices (r² = 0.20–0.45, P < 0.05). In women GRF, maximal eccentric torque and plantar flexor muscle volume were positively correlated with tibial mid-shaft 50 Imax (r² = 0.21–0.35, P < 0.05) (Fig. 1). Distal tibia BSId was not associated with any of the neuromuscular variables in women. No significant relationship was found between activation level and specific tension with the measured bone structural variables.

Correlations between neuromuscular and bone strength variables. Results from men are in grey. Results from women are in black. BSI d distal tibia compressive bone strength index, 50 Imax tibial mid-shaft cortical maximal moment of inertia. GRF maximal ground reaction force in bilateral jumping, ECC T maximal voluntary eccentric torque, Muscle V plantarflexor muscle group volume
In the first regression model with height and weight included, significant explanatory power was reached for distal tibia BSId (46% of the variation explained) and tibial mid-shaft 50 Imax (70% of the variation explained). Specific tension had significant independent explanatory power in tibial mid-shaft 50 Imax (4% of the variation explained). When height and weight were removed from the independent variables the second regression model reached significant explanatory power for distal tibia BSId (44% of the variation explained) and for tibial mid-shaft 50 Imax (65% of the variation explained). In addition to specific tension having significant independent explanatory power in tibial mid-shaft 50 Imax (7% of the variation explained), muscle volume was observed to have significant independent explanatory power in BSId (36% of the variation explained) and in 50 Imax (57% of the variation explained). The total percentages of variation explained by the regression models are given in Table 3.
Discussion
The primary observations in the present study were that muscle volume, maximal voluntary eccentric torque, maximal ground reaction force in jumping and specific tension significantly related to bone structural characteristics. From these parameters the specific tension came out as a new predictor variable in addition to the other variables, which have been previously shown to be importantly involved in muscle-bone interplay (Blain et al. 2001; Calbet et al. 1999; Calmels et al. 1995; Ducher et al. 2005; Macdonald et al. 2006; Nordstrom et al. 1996; Pettersson et al. 1999; Pettersson et al. 2000; Ribom et al. 2004; Sandstrom et al. 2000; Specker et al. 2004; Taaffe and Marcus 2004). The results thus further support the suggestion that appropriate neuromuscular loading is related to bone strength (Burr 1997; Ferretti et al. 2003; Frost 2000).
Specific tension, in addition to muscle volume, had a significant contribution in explaining the variation in tibial mid-shaft 50 Imax. These findings are in line with previous studies by Specker et al. (2004) who found strength as an explaining factor in bone structural differences between rural and non-rural people with comparable body size and with MacDonald et al. (2006) who saw a significant effect of muscle cross-sectional area on bone bending and compressive strength indexes in pre-pubertal boys and girls when body size was accounted for by bone length. In our study variables of muscular force production, muscle volume, specific tension and activation level (Fukunaga et al. 2001) were used in the regression model to explain bone strength. Ducher et al. (2005) have shown previously that muscle volume plays a role in muscle-bone interplay. Concomitant differences in bone and muscle volume in tennis players between dominant and non-dominant hand were observed in the study. Specific tension can vary between individuals depending on muscle architecture (Fukunaga et al. 2001), muscle fiber distribution (Fitts et al. 1991) and activation level (Narici et al. 2004). It was also assumed that activation level could vary inter-individually. Our results indicated that activation level played no role in explaining the bone strength. There seemed to be some variation in activation level among individuals, but evidently activation level is not a major determinative factor in force production in young adults (Merton 1954; Oskouei et al. 2003; Stackhouse et al. 2000). As has been shown previously, our results suggest that, in addition to variation in muscle volume, there is significant inter-individual variation in specific tension (Ikai and Fukunaga 1968; Maughan and Nimmo 1984), which also plays a role in determining the bone strength. Obviously, bone strength is to a great extent dependent on body size. However, manipulating body size to gain bone is non-feasible and thus body-size independent bone strength predictors were sought in the regression. Although specific tension, being size independent variable, was not seen to have any dependency with bone variables in correlation analyses it was selected as a predictor variable for the developed regression models because of its role in determining neuromuscular output.
Altogether our results indicate that the structure of tibial mid-shaft is more strongly dependent on the muscles moving the tibia than the structure of distal tibia. It has been suggested that one of the functions of the trabecular meshwork is to work as a shock absorber underneath joints whereas the purpose of the shaft of the bone is to provide stiff levers for locomotory actions (Currey 2002). Our results seem to support the aforementioned suggestion. Highest compressive loadings on distal part of lower limb in daily activities are likely caused by impacts, which are not necessarily closely related to muscular force production (Komi et al. 1992). In contrast, bending loads on legs are most likely caused by the muscles moving the bone during normal daily activities (Van Buskirk 1989). In addition, distal tibia has functional task in forming the ankle joint. Adult joints and epiphyseal regions are not likely to grow substantially in size, which may explain the lower bone-muscle dependency in the distal tibia observed in our study (Frost 1997). Also, since the functional task plays a role in determining the size and robustness of distal tibia (Carter et al. 1996; Currey 2002), it seems plausible that the adaptations caused by daily activities could not be efficiently measured with the variables used in our study. In addition, the relationship between neuromuscular output and distal bone strength was weaker in females than in males. This discrepancy could be related to the role of female trabecular bone as calcium storage. This could cause the female distal tibia to be even less responsive to loading than the distal tibia of males (Jarvinen et al. 2003).
The parameters used to characterize performance were thought to give a comprehensive measure of neuromuscular function. It needs to be stressed, however, that muscle volume, which was used in the estimation of specific tension, was obtained using indirect methods. Specific tension estimation was further simplified by assuming a constant pennation angle and a constant moment arm to fiber length ratio. These assumptions may cause the specific tension estimate to be more a measure of torque production relative to body size rather than a measure of force production relative to physiological cross-sectional area of the muscle. It was seen that in young adults no relationship exists between activation level and bone. It remains questionable whether the results can be extrapolated to elderly adults. It has been shown that in elderly both specific tension and activation level can be increased with resistance training (Narici et al. 2004) and that resistance training is effective in increasing the skeletal integrity (Martyn-St James and Carroll 2006) and fall prevention (Carter et al. 2001). Thus the current results highlight the possibilities for non-pharmacological interventions, namely physical exercise. Individual determinants of neuromuscular performance, such as specific tension, have contribution in increasing skeletal integrity and can be positively manipulated with exercises, which are also effective in bone strengthening and fall prevention.
Considering the inaccuracies of the estimations made and the nature of indirect measurements conducted, the following conclusions can be drawn: the study emphasizes that tibial strength is related to maximal neuromuscular performance in young men and women. In addition, tibial mid-shaft seems to be more dependent on the neuromuscular performance than distal tibia. The dependency of bone adaptation to neuromuscular performance seems to be moderate, but site, and loading specific.
References
Allen GM, McKenzie DK, Gandevia SC (1998) Twitch interpolation of the elbow flexor muscles at high forces. Muscle Nerve 21:318–328
Babault N, Pousson M, Ballay Y, Van Hoecke J (2001) Activation of human quadriceps femoris during isometric, concentric, and eccentric contractions. J Appl Physiol 91:2628–2634
Blain H, Vuillemin A, Teissier A, Hanesse B, Guillemin F, Jeandel C (2001) Influence of muscle strength and body weight and composition on regional bone mineral density in healthy women aged 60 years and over. Gerontology 47:207–212
Burr DB (1997) Muscle strength, bone mass, and age-related bone loss. J Bone Miner Res 12:1547–1551
Calbet JA, Diaz Herrera P, Rodriguez LP (1999) High bone mineral density in male elite professional volleyball players. Osteoporos Int 10:468–474
Calmels P, Vico L, Alexandre C, Minaire P (1995) Cross-sectional study of muscle strength and bone mineral density in a population of 106 women between the ages of 44 and 87 years: relationship with age and menopause. Eur J Appl Physiol Occup Physiol 70:180–186
Carter DR, Van Der Meulen MC, Beaupre GS (1996) Mechanical factors in bone growth and development. Bone 18:5S–10S
Carter ND, Kannus P, Khan KM (2001) Exercise in the prevention of falls in older people: a systematic literature review examining the rationale and the evidence. Sports Med 31:427–438
Cummings SR, Melton LJ (2002) Epidemiology and outcomes of osteoporotic fractures. Lancet 359:1761–1767
Currey JD (2002) Bones: structure and mechanics. Princeton University Press, Princeton, pp 194–244
Ducher G, Courteix D, Meme S, Magni C, Viala JF, Benhamou CL (2005) Bone geometry in response to long-term tennis playing and its relationship with muscle volume: a quantitative magnetic resonance imaging study in tennis players. Bone 37:457–466
Ferretti JL, Cointry GR, Capozza RF, Frost HM (2003) Bone mass, bone strength, muscle−bone interactions, osteopenias and osteoporoses. Mech Ageing Dev 124:269–279
Fitts RH, McDonald KS, Schluter JM (1991) The determinants of skeletal muscle force and power: their adaptability with changes in activity pattern. J Biomech 24(Suppl 1):111–122
Frost HM (1997) Indirect way to estimate peak joint loads in life and in skeletal remains (insights from a new paradigm). Anat Rec 248:475–483
Frost HM (2000) Muscle, bone, and the Utah paradigm: a 1999 overview. Med Sci Sports Exerc 32:911–917
Fukunaga T, Miyatani M, Tachi M, Kouzaki M, Kawakami Y, Kanehisa H (2001) Muscle volume is a major determinant of joint torque in humans. Acta Physiol Scand 172:249–255
Gass M, Dawson-Hughes B (2006) Preventing osteoporosis-related fractures: an overview. Am J Med 119:S3–S11
Heaney RP (2003) Is the paradigm shifting? Bone 33:457–465
Heinonen A, Kannus P, Sievanen H, Oja P, Pasanen M, Rinne M, Uusi-Rasi K, Vuori I (1996) Randomised controlled trial of effect of high-impact exercise on selected risk factors for osteoporotic fractures. Lancet 348:1343–1347
Heinonen A, McKay HA, Whittall KP, Forster BB, Khan KM (2001) Muscle cross-sectional area is associated with specific site of bone in prepubertal girls: a quantitative magnetic resonance imaging study. Bone 29:388–392
Heinonen A, Sievanen H, Kannus P, Oja P, Vuori I (2002) Site-specific skeletal response to long-term weight training seems to be attributable to principal loading modality: a pQCT study of female weightlifters. Calcif Tissue Int 70:469–474
Ikai M, Fukunaga T (1968) Calculation of muscle strength per unit cross-sectional area of human muscle by means of ultrasonic measurement. Int Z Angew Physiol 26:26–32
Jarvinen TL, Kannus P, Sievanen H (2003) Estrogen and bone—a reproductive and locomotive perspective. J Bone Miner Res 18:1921–1931
Kent-Braun JA, Le Blanc R (1996) Quantitation of central activation failure during maximal voluntary contractions in humans. Muscle Nerve 19:861–869
Komi PV, Fukashiro S, Jarvinen M (1992) Biomechanical loading of Achilles tendon during normal locomotion. Clin Sports Med 11:521–531
Lynch NA, Metter EJ, Lindle RS, Fozard JL, Tobin JD, Roy TA, Fleg JL, Hurley BF (1999) Muscle quality. I. Age-associated differences between arm and leg muscle groups. J Appl Physiol 86:188–194
Macdonald H, Kontulainen S, Petit M, Janssen P, McKay H (2006) Bone strength and its determinants in pre- and early pubertal boys and girls. Bone 39:598–608
Martyn-St James M, Carroll S (2006) High-intensity resistance training and postmenopausal bone loss: a meta-analysis. Osteoporos Int 17:1225–1240
Maughan RJ, Nimmo MA (1984) The influence of variations in muscle fibre composition on muscle strength and cross-sectional area in untrained males. J Physiol 351:299–311
Merton PA (1954) Voluntary strength and fatigue. J Physiol 123:553–564
Miyatani M, Kanehisa H, Ito M, Kawakami Y, Fukunaga T (2004) The accuracy of volume estimates using ultrasound muscle thickness measurements in different muscle groups. Eur J Appl Physiol 91:264–272
Moreland JD, Richardson JA, Goldsmith CH, Clase CM (2004) Muscle weakness and falls in older adults: a systematic review and meta-analysis. J Am Geriatr Soc 52:1121–1129
Narici MV, Reeves ND, Morse CI, Maganaris CN (2004) Muscular adaptations to resistance exercise in the elderly. J Musculoskelet Neuronal Interact 4:161–164
Nicol C, Komi PV, Horita T, Kyrolainen H, Takala TE (1996) Reduced stretch-reflex sensitivity after exhausting stretch-shortening cycle exercise. Eur J Appl Physiol Occup Physiol 72:401–409
Nikander R, Sievanen H, Heinonen A, Kannus P (2005) Femoral neck structure in adult female athletes subjected to different loading modalities. J Bone Miner Res 20:520–528
Nikander R, Sievanen H, Uusi-Rasi K, Heinonen A,Kannus P (2006) Loading modalities and bone structures at nonweight-bearing upper extremity and weight-bearing lower extremity: a pQCT study of adult female athletes. Bone 39:886–894
Nordstrom P, Thorsen K, Nordstrom G, Bergstrom E, Lorentzon R (1995) Bone mass, muscle strength, and different body constitutional parameters in adolescent boys with a low or moderate exercise level. Bone 17:351–356
Nordstrom P, Thorsen K, Bergstrom E, Lorentzon R (1996) High bone mass and altered relationships between bone mass, muscle strength, and body constitution in adolescent boys on a high level of physical activity. Bone 19:189–195
Oskouei MA, Van Mazijk BC, Schuiling MH, Herzog W (2003) Variability in the interpolated twitch torque for maximal and submaximal voluntary contractions. J Appl Physiol 95:1648–1655
Pettersson U, Nordstrom P, Lorentzon R (1999) A comparison of bone mineral density and muscle strength in young male adults with different exercise level. Calcif Tissue Int 64:490–498
Pettersson U, Alfredson H, Nordstrom P, Henriksson-Larsen K, Lorentzon R (2000) Bone mass in female cross-country skiers: relationship between muscle strength and different BMD sites. Calcif Tissue Int 67:199–206
Ribom E, Ljunggren O, Piehl-Aulin K, Ljunghall S, Bratteby LE, Samuelson G, Mallmin H (2004) Muscle strength correlates with total body bone mineral density in young women but not in men. Scand J Med Sci Sports 14:24–29
Sandstrom P, Jonsson P, Lorentzon R, Thorsen K (2000) Bone mineral density and muscle strength in female ice hockey players. Int J Sports Med 21:524–528
Schonau E (2004) The peak bone mass concept: is it still relevant? Pediatr Nephrol 19:825–831
Specker B, Binkley T, Fahrenwald N (2004) Rural versus nonrural differences in BMC, volumetric BMD, and bone size: a population-based cross-sectional study. Bone 35:1389–1398
Stackhouse SK, Dean JC, Lee SC, Binder-MacLeod SA (2000) Measurement of central activation failure of the quadriceps femoris in healthy adults. Muscle Nerve 23:1706–1712
Stevens JA, Olson S (2000) Reducing falls and resulting hip fractures among older women. MMWR Recomm Rep 49:3–12
Suominen H (2006) Muscle training for bone strength. Aging Clin Exp Res 18:85–93
Taaffe DR, Marcus R (2004) The muscle strength and bone density relationship in young women: dependence on exercise status. J Sports Med Phys Fitness 44:98–103
Taaffe DR, Pruitt L, Lewis B, Marcus R (1995) Dynamic muscle strength as a predictor of bone mineral density in elderly women. J Sports Med Phys Fitness 35:136–142
Van Buskirk WC (1989) Elementary stress analysis of the femur and tibia. In: Cowin SC (ed) Bone mechanics. CRC, Florida, pp 43–51
Acknowledgments
The study was funded by the Academy of Finland. The authors would like to thank Ph. D. Vincent Martin, Ph. D. Masaki Ishikawa, MSc. Merja Hoffren and Mr. Matthew Holmes for their valuable help in the measurements.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Rantalainen, T., Heinonen, A., Komi, P.V. et al. Neuromuscular performance and bone structural characteristics in young healthy men and women. Eur J Appl Physiol 102, 215–222 (2008). https://doi.org/10.1007/s00421-007-0575-8
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-007-0575-8