
Abstract
This study was designed to determine the regional differences of magnetic resonance (MR) measurements in the hamstrings [biceps femoris muscle long head (BFl) and short head (BFs), semimembranosus muscle (SM), and semitendinosus muscle (ST)] following eccentric knee-flexion exercise. Twelve male volunteers performed eccentric knee-flexion exercise. Maximum isometric torque, plasma creatine kinase (CK) activity, muscle soreness, and MR images of the hamstrings were measured before and immediately following exercise, and on the first, second, third and seventh days following the exercise. Cross-sectional areas (CSAs) and transverse relaxation times (T2s) of the hamstrings were measured from the T2-weightened MR imaging sequences of 30% (proximal), 50% (middle), and 70% (distal) areas of the thigh length. The CSA of the ST at proximal and middle regions had significantly increased on the third day, but no significant changes were found for the BFl or SM. Immediately following exercise, T2 values had increased significantly in the BFs, BFl, and ST. On the third day, T2 values of only ST increased significantly at proximal, middle and distal regions. Significant differences of T2 values between proximal and distal regions in the ST were found for the second, third and seventh days following the exercise. These results suggest that because of the anatomical characteristics of the muscles, the degrees of response following the exercise differed among the muscles and the regions of ST muscle.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The hamstring muscles include the four muscles [biceps femoris muscle long head (BFl) and short head (BFs), semimembranosus muscle (SM) and semitendinosus muscle (ST)] located in the posterior of the thigh. The architectures and innervation patterns of the respective muscles differ (Friederich and Brand 1990; Wickiewicz et al. 1983; Woodley and Mercer 2005). The BFl has an intermediate fascicle length and a physiological cross-sectional area (CSA) compared with the other hamstring muscles, whereas the BFs have a long fascicular and a small physiological CSA. The SM has a short fascicular length and a large physiological CSA. The ST has unique architectural characteristics among the hamstring muscles. A tendinous intersection (TI) is present within the muscle belly; it divides the muscle into two (proximal and distal) distinct regions (Wickiewicz et al. 1983; Lee et al. 1988; Woodley and Mercer 2005). Because the fascicular lengths of the proximal and distal regions are almost equal and because the fascicles in the two regions are in series, the ST has generally been treated as a single muscle. However, almost all the fascicles in the proximal region are inserted to the TI, those in the distal region originate from the TI (Woodley and Mercer 2005). Moreover, the two regions are innervated via two branches from the tibial part of the sciatic nerve: one proximal and one distal of the TI. In the hamstring muscles, only the ST muscle was partitioned on the basis of both architecture and innervation. Consequently, it is assumed that the degree of involvement and/or response of an exercise differ between the proximal and distal regions of the ST.
The results of numerous studies have shown that the anatomical characteristics of a muscle are the primary determinants of the functional properties (Lieber and Bodine-Fowler 1993; Lieber and Friden 2000). The hamstring muscles generally activate during knee-flexion and/or hip extension, and deal with one skeletal muscle group. However, taking into consideration the different architectural characteristics of each hamstring muscle, it is conceivable that respective muscles have inherent functions and compensate for each other. Many cases of hamstring strain, for example, involve the biceps femoris, although the semitendinosus and semimembranosus muscle are less injured, which might reflect the architectural and functional differences of the muscles (Heiderscheit et al. 2005; Thelen et al. 2005).
Recently, magnetic resonance (MR) imaging has been used to assess skeletal muscle functions. The transverse relaxation time (T2) is a quantitative index of muscle activation. Intensive exercise is known to produce changes in the amount and distribution of water in skeletal muscle. This method can non-invasively monitor the physiological changes of the recruited muscle during exercise. In fact, MR imaging can be used to assess damaged muscles following intensive exercise (Leblanc et al. 1993; Clarkson and Hubal 2002). Many authors have reported that the T2 value increases following eccentric exercise (Jayaraman et al. 2004; Larsen et al. 2007; Prior et al. 2001; Segal and Song 2005; Sesto et al. 2005) and that T2 value is positively correlated with plasma creatine kinase (CK) activity, reflecting exercise-induced muscle damage (Larsen et al. 2007; LeBlanc et al. 1993; Schwane et al. 2000).
Moreover, earlier studies have investigated the inter-muscle differences and intra-muscle regional differences of T2 value changes between proximal and distal regions (Akima et al. 2004; Segal and song 2005). However, to our knowledge, no regionally specific differences of the morphology and T2 value have been reported comprehensively for the hamstring muscles. These muscles are long and multiarticular, representing a complex of fusiform and pennate muscles. Therefore, the hamstring muscles would show each muscle’s specific characteristics of architectural change following intensive eccentric exercise. A detailed examination of the changes in MR measurements of the hamstring muscles is applicable to the understanding of the functional difference between the muscles, and of the pathomechanics of the muscles.
In light of these considerations, we hypothesized that the degree of the response following the intensive exercise would be different, as represented by different changes in MR measurements, such as the CSAs and T2 values, among hamstring muscles and between proximal and distal regions of each muscle. This study was designed to investigate the regional specific differences of MR measurements in the hamstring muscles following eccentric knee-flexion exercise.
Methods
Subjects
This study examined 12 healthy young male volunteers with no history of neuromuscular or orthopedic disease (age 23.7 ± 1.8 years; height 171.8 ± 4.8 cm; weight 66.9 ± 8.6 kg). None were participating in any regular training regime. Subjects were instructed to avoid activities and not to use icing or anti-inflammatory medication for the week preceding and the week of the experiment. This study was approved by the Human Research Ethics Committee of the School of Sport Sciences of Waseda University and is consistent with their requirements for human experimentation. This study conforms to the Declaration of Helsinki. Written informed consent statements were obtained after participants had read the volunteer information sheet and questions related to the study had been answered to their satisfaction.
Exercise protocol
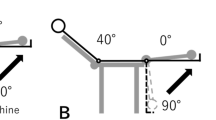
After a few minutes of warming up, subjects performed eccentric exercise of the hamstring muscles with the right leg using a plate-loaded knee-flexion machine (Prone Leg Curl; Nautilus, USA), which was adjusted to 120% of the 1 repetition maximum (1RM).
Subjects were instructed to lower the weight from a knee-flexed position (100°) to a knee-extended position (0°) in 3 s, maintaining the lowering velocity as constant as possible by following the examiner’s counting of “0” for the beginning and “1, 2, and 3” for the movement with planter flexion of the ankle to reduce the contribution of the gastrocnemius muscle. Subjects were verbally encouraged to generate maximal force at the starting position and to resist maximally against the knee-extending action throughout the range of motion. The weight was raised after each eccentric repetition by an examiner; therefore, the overall exercise task was eccentric only for the subject. This exercise was repeated for five sets of 10 repetitions each, with at least a 3-min rest between sets.
Criterion measures
Before and immediately following exercise, and on the first, second, third and seventh days following the exercise, maximum isometric knee-flexion torques were calculated, and plasma creatine kinase (CK) activity and muscle soreness were assessed. In addition, MR imaging of the thigh was performed.
MVC
Maximum voluntary contractions (MVCs) of the knee-flexor muscles were measured using a modified force-measuring machine that included a force gauge (LTZ-200KA; Kyowa Electronic Instruments, Tokyo, Japan) connected to an analog-to-digital converter (LEG-1000; Nihon Kohden, Tokyo, Japan). The metal cord with the force gauge was mounted to the ankle joint. Subjects were prone on a bed with the hip joint at 0° of flexion and abduction, with the knee joint at 15° of flexion. Subjects maintained each MVC for 3 s. They repeated the task two times with at least 3 min rest between tests. Maximum isometric knee-flexion torque was calculated from the MVC value. The highest torque was used for subsequent analyses.
Blood analysis
A 10-ml sample of blood was drawn from a branch of the antecubital vein. The blood was allowed to clot for 30 min at room temperature; it was then centrifuged for 10 min to obtain serum. After separation, all serum samples were stored at −20°C until analysis for CK activity. The CK enzyme activity was measured in the laboratory. Because of the high degree of variability associated with plasma CK, the values were log-transformed to satisfy the analysis of variance (ANOVA) procedure.
Muscle soreness
Muscle soreness was evaluated using a visual analog scale (VAS) consisting of a 100-mm continuous line representing “no pain” at one end (0 mm) and “unbearable pain” at the other (100 mm). Other examiners (Chen et al. 2007; Nosaka and Sakamoto 2001) have used and described this scale previously. Subjects lay prone on a bed and were asked to indicate the muscle soreness level on the line when an investigator pressed the BFl, ST, and SM muscle belly with 4 kg/cm force using a pressure meter (Igarashi Ika Kogyo, Tokyo, Japan). The pressure points for each muscle were marked to facilitate identical pressures at a same position over several days.
MRI
All MR images of the thigh were performed using a 1.5-T whole body imager (Magnetom Symphony; Siemens-Asahi Medical Technologies, Tokyo, Japan). For the MR imaging scans, subjects were positioned supine with their knee flexed. Subjects were secured in a leg-holding device that was fitted to the inside of the coil to maximize the repeatability of limb placement in the imager. Then, T2-weightened transverse spin-echo MR axial images [repetition time (TR) = 2,000 ms, echo time (TE) = 30, 45, 60, and 75 ms] were collected beginning with a single scan at the lower end of ischial tuberosity using a 256 × 256 image matrix, with a 270 mm field of view, 10-mm slice thickness, and 12-mm interslice gap using a body coil.
The MRI data were evaluated for anatomical CSA and T2 relaxation time (T2 value) of the hamstring muscles. In the evaluations, the images containing of the areas at 30% (proximal), 50% (middle) and 70% (distal) of thigh length from the upper border of the ischial tuberosity (0%) to the lower border of the tibial plateau (100%) were used. The MR images were transferred to a personal computer in the Digital Imaging and Communications in Medicine (DICOM) file format; image manipulation and analysis software (OSIRIS, University Hospital of Geneva, Switzerland) was used to measure CSA and the signal intensity of each hamstring muscle (BFs, BFl, ST and SM). The region of interest was defined by tracing the outline of the muscles, avoiding visible aponeurosis, vessels, fat, membranes, and the femur. The signal intensity was measured from the same region for all four TEs. A T2 measurement sequence with four TEs was applied to measure the absolute T2 value. Images taken at different TEs were fit to a monoexponential time curve to extract the T2 values based on the formula: SI = M 0 × exp(−TE/T2), where SI represents the signal intensity at a given TE and M 0 is the original MRI signal intensity. The same person performed the MR imaging scan and the T2 calculation.
Statistical analysis
Changes in the maximum isometric knee-flexion torque, plasma CK activity, muscle soreness measured over day were compared using one-way ANOVA with repeated measures. Significant differences of the CSAs and T2 values of hamstring muscles following the eccentric exercise were determined using two-way ANOVA with repeated measures (muscle region × day). Bonferroni’s post hoc analysis was conducted if the ANOVA showed statistically significant main effects or interaction effects. Statistical significance was set at P < 0.05 for the ANOVA and P < 0.003 for the post hoc test. All statistical analyses were conducted using a statistical analysis software program (SPSS ver. 14.0; SPSS, Japan, Tokyo, Japan). Descriptive data are expressed as mean ± SD.
Results
MVC
Maximum isometric knee-flexion strength, as measured before, immediately after, and on the first, second, third, and seventh days following the exercise, is shown in Fig. 1. The maximum torques showed a significant day effect (F 5,55 = 10.9; P < 0.001). The maximum torque was decreased significantly to 25.8% immediately following the exercise (P < 0.003). By the seventh day, the torque had not recovered to its initial values; it also showed a trend toward pre-values.

Change in maximum isometric knee-flexion torque before (pre), immediately after (post), and on the first, second, third, and seventh days following eccentric exercise. Asterisks indicate significant differences from the pre-value (*P < 0.003)
Plasma CK activity
Plasma CK values were transformed to a natural log scale (Fig. 2), revealing a significant day effect (F 5,55 = 19.5; P < 0.001). The CK activity was significantly higher on the second, third, and seventh days than the pre-value (P < 0.003). The value reached its peak on the third day following exercise.

Change in plasma creatine kinase (CK) activity (natural log) before (pre), immediately after (post), and on the first, second, third, and seventh days following eccentric exercise. Asterisks indicate significant differences from the pre-value (*P < 0.003)
Muscle soreness
The changes in muscle soreness of the BFl, ST, and SM are presented in Table 1. They showed a significant day effect for the BFl (F 5,55 = 7.7; P < 0.001), ST (F 5,55 = 8.3; P < 0.001), and SM (F 5,55 = 4.1; P < 0.01). The significant increases in muscle soreness were observed in the BFl and ST; it peaked on the second day following exercise (P < 0.003). Soreness in the SM showed no significant change following the exercise.
MRI
Typical T2-weightened MR images of the right thigh before and following exercise are presented in Fig. 3. The CSA of the ST was increased on the third day (Fig. 4, lower left). The brightness of hamstring muscles increased immediately following exercise. On the third day following exercise, the ST showed a conspicuous increase in brightness (Fig. 5, lower left).

Representative T2-weightened magnetic resonance images of the middle region (50% of thigh length) from one subject before, immediately after, and on the first, second, third, and seventh days following eccentric exercise (BFs biceps femoris short head muscle, BFl biceps femoris long head muscle, ST semitendinosus muscle, SM semimembranosus muscle)

Change in the cross-sectional area (CSA) of the 30% (proximal), 50% (middle) and 70% (distal) of thigh length in the biceps femoris short head muscle (BFs) and long head muscle (BFl), semitendinosus muscle (ST), and semimembranosus muscle (SM) before (pre), immediately after (post), and on the first, second, third, and seventh days following eccentric exercise. Asterisks indicate significant differences from the pre-value (*P < 0.003)

Change in the transverse relaxation time (T2 value) of the 30% (proximal), 50% (middle) and 70% (distal) of thigh length in the biceps femoris short head muscle (BFs) and long head muscle (BFl), semitendinosus muscle (ST), and semimembranosus muscle (SM) before (pre), immediately after (post), and on the first, second, third, and seventh days following eccentric exercise. All values are given as a percentage of the pre-value. Asterisks indicate significant differences from the pre-value (*P < 0.003) and alphabets indicate significant differences between regions (a P < 0.017 vs. the middle region, b P < 0.017 vs. the distal region)
CSA
The time course of the changes in CSA of the hamstring muscles is shown in Fig. 4. As shown in this figure, the BFs showed a significant main effect for the muscle region (F 1,11 = 30.0; P < 0.001) and day (F 5,55 = 6.9; P < 0.001), but no muscle region showed a significant by day interaction effect. The CSA of the BFs distal region on the second and seventh day was significantly higher than that before exercise (P < 0.003). On the third day, the CSA showed a trend toward higher values than the pre-values. The ST showed a significant main effect for the muscle region (F 2,22 = 84.2; P < 0.001) and day (F 5,55 = 15.0; P < 0.001), and also muscle region by day interaction effects (F 10,110 = 6.8; P < 0.001) were observed for the ST CSA. The CSA of the proximal region on the third day was significantly higher than that before exercise and remained so until at least the sevnth day (P < 0.003). The CSA of the middle region in ST was higher immediately following exercise, on the second, third and seventh days (P < 0.003) compared with the level before exercise. The BFl and SM showed a significant main effect for the muscle region (F 2,22 = 23.9; P < 0.001, F 2,22 = 100.2; P < 0.001), but no main effect for the muscle region and interaction effect was found between muscle regions and days.
T2
Figure 5 presents changes in the T2 values of the hamstring muscles. A significant main effect for the muscle region (F 1,11 = 37.6; P < 0.001) and day (F 5,55 = 4.4; P < 0.001), and interaction effect between muscle regions and days (F 5,55 = 4.2; P < 0.01) were observed for the T2 values in the BFs. Both the middle and distal regions of the BFs showed elevated T2 values immediately following exercise (P < 0.003). The T2 values immediately after and on the third day following exercise were significantly different between the middle and distal regions (P < 0.01). The ST T2 values showed a significant main effect for muscle region (F 2,22 = 17.4; P < 0.001) and day (F 5,55 = 13.1; P < 0.001), and interaction effect between muscle regions and days (F 10,110 = 3.9; P < 0.001) for the ST T2 values. The T2 values were obtained around 40 ms as pre-values, i.e., in the proximal region (38.7 ± 1.4 ms), middle region (37.5 ± 2.3 ms), and distal region (37.4 ± 1.8 ms) of the ST. The T2 values on the ST proximal region were elevated immediately after (51.1 ± 5.7 ms), and on the second day (65.1 ± 28.1 ms), the third day (71.2 ± 23.7 ms), and seventh day (75.5 ± 28.1 ms) following exercise (P < 0.003). In the middle and distal regions in the ST, the T2 values increased immediately after (52.3 ± 5.9 ms; 50.9 ± 8.5 ms, respectively), and were significantly higher on the third day (67.5 ± 23.8%; 60.8 ± 2.7 ms, respectively), and on the seventh day (71.4 ± 26.2 ms; 68.6 ± 27.0 ms, respectively). The proximal T2 value was lower than that the middle T2 value immediately following exercise (P < 0.01), and was higher than the distal T2 value on the second and third day following exercise (P < 0.01). In the BFl, a significant day effect (F 5,55 = 3.8; P < 0.01) was observed for the T2 values, but no day effect and muscle region by day interaction effect occurred. The T2 values immediately following exercise were significantly elevated on the middle and distal regions (P < 0.01). No T2 value changes were apparent in any regions of the SM following exercise.
Discussion
This study examined differences of changes in MR measurements, represented as CSA and T2 values, among hamstring muscles. Results showed that almost all the hamstring muscles exhibited a T2 increase immediately following intensive eccentric exercise. Moreover, the ST presented an increase of CSA and T2 values following the exercise, along with increases in the regional differences of the T2 values immediately following exercise, and on the second and third days.
It is generally considered that the hamstring muscles are similarly activated in knee-flexion exercise. However, results of this study showed that the ST muscle especially changed the MR measurements following the intensive eccentric exercise. This result suggested that the respective responses of the hamstring muscles following the exercise differed among hamstring muscles; the ST was particularly damaged. The changes in MR measurements of the ST relate to the architectural characteristics. The ST is a fusiform, thin, and biarticular long muscle, whereas the BFl and SM are pennate and bulky muscles. The results of a previous study indicated that the greater the degree of passive extension to failure in the pennate muscle tended to be greater than that in the fusiform muscle (Garrett et al. 1988). For this reason, the fusiform muscle is potentially more easily injured than the pennate muscle. The results of that study showed the increase of T2 values in the ST. Moreover, the ST has a TI at about the mid-point of the muscle (Lee et al. 1988; Woodley and Mercer 2005). The TI divides the ST muscle into proximal and distal neurovascular compartments, with each region innervated by a different nerve branch. These neuroanatomical characteristics are associated mainly with the notable changes of MR measurements in the ST.
This region-specific difference of changes in MR measurements among the proximal, middle and distal regions in the ST suggests that these regions might have different functional roles. It is interesting that the proximal and middle regions show a CSA increase and greater T2 changes than the distal region in the ST. A possible cause of the regional differences is the presence of the TI, which is contained in the proximal and middle regions, although it is not contained in the distal region. The ending architecture of the muscle fibers near TI resembles the muscle–tendon junction (MTJ) architecture. The MTJ or muscle fibers near the MTJ were more damaged within a muscle when muscle–tendon units were loaded intensively (Tidball et al. 1993). Consequently, the MR measurements in the proximal and middle ST regions were more changed than those in the distal region. Moreover, the TI induced regional differences of the EMG activity, work distribution, and metabolic state. Consequently, as results of previous studies have suggested (Adams et al. 1992; Jenner et al. 1994; Kinugasa et al. 2006; Vandenborne et al. 2000), the regional difference of the T2 value might reflect the muscle-cell metabolism and fluid uptake, represented as the regional difference of neuromuscular and/or metabolic activities.
In this study, the ST especially showed a conspicuous increase of the CSA and T2 values. The delayed increase of muscle-injury markers, such as muscle-swelling, muscle soreness, and plasma CK activity following the exercise that subjects performed in the present study, represented a similar time course to that of the previous studies (Harrison et al. 2001; Jayaraman et al. 2004; Nosaka and Sakamoto 2001). However, compared with those previous studies the degree of the reduction of maximal isometric knee-flexion torque was lower, and the plasma CK activity was higher. In addition, the MR images showed the delayed onset muscle damage signs only in the ST. The eccentric knee-flexion exercise using a prone leg-curl machine with 120%MVC loads can cause damage, especially to the ST in the hamstring muscles. Biomechanical analyses of the intensive eccentric knee-flexion exercise, such as the force contribution of the individual muscles and the three-dimensional knee-joint kinematics, are necessary to resolve the issue of why only the ST showed the T2 and CSA changes following intensive eccentric knee-flexion exercise.
The changing mechanism of the increased T2 values and CSA remains unknown. The exercise-induced T2 increases in skeletal muscle involve not only the osmotic movement of muscle water into the myofibrillar space but also the vascular dilation or increase of blood flow. In this study, although all muscles and regions showed a T2 increase immediately following exercise, the increase of T2 values was shown only in the ST on the second day following exercise. The relationship between the changes of the plasma CK activity and of the T2 value of the ST was not statistically significant only immediately following exercise: it became statistically significant from the second day following exercise (data not shown). The plasma CK activity indicates much skeletal muscle damage, and reflects a delayed increase following unaccustomed eccentric exercise. Therefore, it is conceivable that the T2 increase that occurred immediately following exercise reflects the increased blood flow, and that the increase that occurred after the second day following exercise reflects severe muscle damage.
Recently, Baczkowski et al. (2006) examined thigh-muscle activations using MR imaging method before and after 100 Australian Rules football kicks. The ST muscle showed great increases of the signal intensity in the kicking and stance legs, whereas the BFl, which is the most commonly injured muscle among football players (Askling et al. 2007; Connell et al. 2004; Koulouris and Connel 2006), showed minimal change in both legs. The hamstring muscles of both legs can act eccentrically during and after the ball contact. Compared with the SM and BF, the increases of the ST signal intensity were shown in an actual kicking performance, indicating the important contribution of the ST. However, the workload of the hamstring muscles might be lower than that of the quadriceps femoris muscles during the kicking performance. Consequently, each hamstring muscle might have different sensitivities to eccentric stimulation. The ST might sensitively respond to the eccentric contraction. Further studies are necessary to clarify the pathomechanics and relationships between the injury factors and the observed injury rate in the hamstring muscles.
In summary, the present study has shown that the changes of the CSAs and T2 values following intensive knee-eccentric flexion exercise differed among hamstring muscles. This result suggested that the hamstring muscles that were activated similarly in knee-flexion tasks differed in their respective degrees of response following the exercise. Moreover, the ST showed the region-specific differences of the MR measurements following the exercise, which affected the anatomical characteristics of the ST muscle, especially the presence of the TI.
References
Adams GR, Duvoisin MR, Dudley GA (1992) Magnetic resonance imaging and electromyography as indexes of muscle function. J Appl Physiol 73:1578–1583
Akima H, Takahashi H, Kuno SY, Katsuta S (2004) Coactivation pattern in human quadriceps during isokinetic knee-extension by muscle functional MRI. Eur J Appl Physiol 91:7–14
Askling CM, Tengvar M, Saartok T, Thorstensson A (2007) Acute first-time hamstring strains during high-speed running: a longitudinal study including clinical and magnetic resonance imaging findings. Am J Sports Med 35:197–206
Baczkowski K, Marks P, Silberstein M, Schneider-Kolsky ME (2006) A new look into kicking a football: an investigation of muscle activity using MRI. Australas Radiol 50:324–329
Chen TC, Nosaka K, Sacco P (2007) Intensity of eccentric exercise, shift of optimum angle, and the magnitude of repeated-bout effect. J Appl Physiol 102:992–999
Clarkson PM, Hubal MJ (2002) Exercise-induced muscle damage in humans. Am J Phys Med Rehabil 81:S52–S69
Connell DA, Schneider-Kolsky ME, Hoving JL, Malara F, Buchbinder R, Koulouris G, Burke F, Bass C (2004) Longitudinal study comparing sonographic and MRI assessments of acute and healing hamstring injuries. AJR Am J Roentgenol 183:975–984
Friederich JA, Brand RA (1990) Muscle fiber architecture in the human lower limb. J Biomech 23:91–95
Garrett WE Jr, Nikolaou PK, Ribbeck BM, Glisson RR, Seaber AV (1988) The effect of muscle architecture on the biomechanical failure properties of skeletal muscle under passive extension. Am J Sports Med 16:7–12
Harrison BC, Robinson D, Davison BJ, Foley B, Seda E, Byrnes WC (2001) Treatment of exercise-induced muscle injury via hyperbaric oxygen therapy. Med Sci Sports Exerc 33:36–42
Heiderscheit BC, Hoerth DM, Chumanov ES, Swanson SC, Thelen BJ, Thelen DG (2005) Identifying the time of occurrence of a hamstring strain injury during treadmill running: a case study. Clin Biomech (Bristol, Avon) 20:1072–1078
Jayaraman RC, Reid RW, Foley JM, Prior BM, Dudley GA, Weingand KW, Meyer RA (2004) MRI evaluation of topical heat and static stretching as therapeutic modalities for the treatment of eccentric exercise-induced muscle damage. Eur J Appl Physiol 93:30–38
Jenner G, Foley JM, Cooper TG, Potchen EJ, Meyer RA (1994) Changes in magnetic resonance images of muscle depend on exercise intensity and duration, not work. J Appl Physiol 76:2119–2124
Kinugasa R, Kawakami Y, Fukunaga T (2006) Quantitative assessment of skeletal muscle activation using muscle functional MRI. Magn Reson Imaging 24:639–644
Koulouris G, Connell D (2006) Imaging of hamstring injuries: therapeutic implications. Eur Radiol 16:1478–1487
Larsen RG, Ringgaard S, Overgaard K (2007) Localization and quantification of muscle damage by magnetic resonance imaging following step exercise in young women. Scand J Med Sci Sports 17:76–83
LeBlanc AD, Jaweed M, Evans H (1993) Evaluation of muscle injury using magnetic resonance imaging. Clin J Sport Med 3:26–30
Lee TC, O’Driscoll KJ, McGettigan P, Moraes D, Ramphall S, O’Brien M (1988) The site of the tendinous interruption in semitendinosus in man. J Anat 157:229–231
Lieber RL, Bodine-Fowler SC (1993) Skeletal muscle mechanics: implications for rehabilitation. Phys Ther 73:844–856
Lieber RL, Friden J (2000) Functional and clinical significance of skeletal muscle architecture. Muscle Nerve 23:1647–1666
Nosaka K, Sakamoto K (2001) Effect of elbow joint angle on the magnitude of muscle damage to the elbow flexors. Med Sci Sports Exerc 33:22–29
Prior BM, Jayaraman RC, Reid RW, Cooper TG, Foley JM, Dudley GA, Meyer RA (2001) Biarticular and monoarticular muscle activation and injury in human quadriceps muscle. Eur J Appl Physiol 85:185–190
Schwane JA, Buckley RT, Dipaolo DP, Atkinson MA, Shepherd JR (2000) Plasma creatine kinase responses of 18-to 30-yr-old African-American men to eccentric exercise. Med Sci Sports Exerc 32:370–378
Segal RL, Song AW (2005) Nonuniform activity of human calf muscles during an exercise task. Arch Phys Med Rehabil 86:2013–2017
Sesto ME, Radwin RG, Block WF, Best TM (2005) Anatomical and mechanical changes following repetitive eccentric exertions. Clin Biomech (Bristol, Avon) 20:41–49
Thelen DG, Chumanov ES, Hoerth DM, Best TM, Swanson SC, Li L, Young M, Heiderscheit BC (2005) Hamstring muscle kinematics during treadmill sprinting. Med Sci Sports Exerc 37:108–114
Tidball JG, Salem G, Zernicke R (1993) Site and mechanical conditions for failure of skeletal muscle in experimental strain injuries. J Appl Physiol 74:1280–1286
Vandenborne K, Walter G, Ploutz-Snyder L, Dudley G, Elliott MA, De Meirleir K (2000) Relationship between muscle T2* relaxation properties and metabolic state: a combined localized 31P-spectroscopy and 1H-imaging study. Eur J Appl Physiol 82:76–82
Wickiewicz TL, Roy RR, Powell PL, Edgerton VR (1983) Muscle architecture of the human lower limb. Clin Orthop Relat Res 179:275–283
Woodley SJ, Mercer SR (2005) Hamstring muscles: architecture and innervation. Cells Tissues Organs 179:125–141
Acknowledgments
The authors gratefully acknowledge N. Tawara and Y. Iwahara for their technical assistance to this research. We also thank the Japan Institute of Sports Sciences. This work was supported by Basic Research (B) (19650179) in 2007 and 2008.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Kubota, J., Ono, T., Araki, M. et al. Non-uniform changes in magnetic resonance measurements of the semitendinosus muscle following intensive eccentric exercise. Eur J Appl Physiol 101, 713–720 (2007). https://doi.org/10.1007/s00421-007-0549-x
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-007-0549-x