
Abstract
The aim of this study was to determine time-of-day variations of grip force control during a manipulative task. In particular, we repeatedly tested continuous up and down movements of a hand held instrumented object on two consecutive days. The movements caused fluctuations in the load that had to be compensated for by adequate grip forces. The level of grip force, the acceleration of the arm movement, the ratio of grip and load force as well as the temporal relationship between the two force profiles and the subjective sleepiness were examined. The time-of-day protocol lasted 26 h with sessions at different times of day, and sleep at night. Diurnal variations were analyzed with an intersession interval of 2 h; detailed analyses were carried out for the evening sessions until bed time and the sessions in the morning after awake. The present study proves a time-of-day effect for the level of grip force as well as for the acceleration. The effect was most obvious during the morning sessions when both parameters increased. The ratio of grip force to load, i.e., independent of the actual load and acceleration, shows a similar modulation as grip force. This later finding suggests that grip force changed partly independently from changes of arm acceleration. Time of day did not influence the strength of the cross-correlation and the synchronicity between the two force profiles. Thus, while grip force changed with time of day, the predictive coupling of the grip force according to the load was constant.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Human physiology and behaviour is largely influenced by chronobiological rhythms. A well documented circadian rhythm is our body temperature, which shows its maximum in the early evening and its minimum in the early morning hours. Like body temperature, several parameters (e.g., nerve conduction velocity, elbow flexion strength and vigilance) suggest optimal physical performance during the afternoon and early evening (Winget et al. 1985). However, little is known yet about time-of-day influences on fine motor performance during manipulative tasks, although skilled and dexterous manual manipulations constitute an inherent feature of every day life. We probably all have experienced numbness and increased danger of slipping when manipulating objects in the early morning. Time-of-day effects on fine motor control can be highly relevant for work scheduling; for example, how precise is a doctor on duty woken up from sleep in fine manual manoeuvres like using a scalpel?
An established paradigm in research on circadian rhythms in motor control is the production of maximum strength. One of the first diurnal investigations on maximum grip strength disclosed a marked diurnal variation with an increase in the morning and a fall in strength at night, which was in parallel with the rhythms of body temperature (Wright 1959). Ilmarinen et al. (1980) and Atkinson et al. (1993) confirmed these results with a maximum of hand grip strength in the early evening (18:00–19:00 h) and a minimum in the morning (6:00–7:00 h). Strength of grip in the morning immediately after the subjects were awakened (6:30–9:15 h; Jeanneret and Webb 1963) as well as at 8:30–10:00 h (Pearson et al. 1982) was lower, compared with control measurements in the afternoon. Stolz et al. (1988) assessed grip strength during a 24 h cycle and found a minimum at 5:00 h in the morning, followed by improved performance during the day until grip strength was maximum at 20:00 h in the evening. In contrast statistical analysis of the data presented by Lawrie et al. (2000) showed no difference in maximum grip strength between morning (8:00 h) and evening (20:00 h) sessions in healthy controls (Strutton et al. 2003).
Spontaneous motor tempo does not involve maximal motor output, but is self paced at the individual’s preferred and comfortable rate. Dosseville et al. (2002) instructed their subjects to tap as regularly as possible with their index finger at the rate that seemed most natural to them. The spontaneous tapping tempo revealed a significant time-of-day effect with the highest value at 18:00 h and least taps at 6:00 h. This pattern was confirmed by Moussay et al. (2002), who found a significant increase in the spontaneous tapping tempo during the day and a slowdown in the evening after 18:00 h. Similarly in the afternoon the spontaneous pedal rate on a cycle ergometer was higher than in the morning (Moussay et al. 2002, 2003).
In summary, the results of maximum grip strength as well as spontaneous motor tempi exhibit an influence of time-of-day similar to the diurnal rhythm in body temperature.
In every day life we constantly face demands on fast and precise execution of psychomotor processes that do not imply the production of maximum force. When we manipulate an object our grip forces have to counteract the physical loads, in order to prevent the object from slipping out of the hand. In particular grip forces need to compensate the object’s weight (product of mass and gravitation), and additionally have to adapt to the surface friction of the grasped object. Furthermore grip forces should be low to allow fine movements of the fingers with the grasped object and should avoid fatigue. Johansson and Westling (1984) and Johansson (1996) demonstrated high precision and effectiveness in the adaptation of grip force to changes in load force while lifting and holding an object. Grip force was just slightly higher than the minimum to prevent slipping, indicating a small safety margin.
Every time we move a grasped object, e.g., when lifting a cup to drink, our grip force has to adapt to the load, that is composed of gravitational force and acceleration-induced inertial loads. It has been shown that grip force is modulated in parallel with load changes resulting from object movements, for example during discrete or cyclic vertical movements of a hand-held object (Flanagan and Wing 1993, 1995). Grip force regulation is just slightly higher than the minimum to prevent the object from slipping. The coupling between grip force and load force is in close synchrony indicting that grip force precisely anticipated the destabilizing load forces.
Grip force is modulated in parallel with load in a variety of grips (Flanagan and Tresilian 1994), regardless of surface texture or movement frequency (Flanagan and Wing 1995). Nevertheless dexterous hand function is vulnerable to specific disturbances. Hermsdörfer, Nowak and co-workers (2003) examined factors which might deteriorate grip force control during continuous cyclic or discrete vertical arm movements with a grasped object. Nowak and Hermsdörfer (2003) reduced the tactile sensibility by cooling the grasping digits and found no influence on the ability to adjust the grip force to the movement induced fluctuations in load. However, the scaling of grip force level was impaired as obvious from significantly higher grip forces after digit cooling. Patients with cerebral stroke, which caused sensory impairments, applied massively increased grip forces, while the synchronicity between the grip and load forces was preserved in most patients (Hermsdörfer et al. 2003). Furthermore grip force was precisely adjusted to changes in load even if these were due to gross changes in gravity conditions produced by parabolic flight manoeuvres (Hermsdörfer et al. 2000).
Given this evidence, it seems obvious to study whether time-of-day affects fine motor control. For this reason manipulative force control was assessed in a time-of-day protocol, which was defined by Valdez et al. (2005) as one of the basic protocols to measure circadian variations. The time-of-day recording includes sessions at different times of day over the normal working day in subjects living their usual personal and social activities. This protocol does not interfere with the sleep–wake cycle of the subjects as they live at their normal sleep–wake schedule.
We selected continuous (cyclic) vertical movements as the motor task to be tested. This task has proven to be highly sensitive reflecting performance changes under perturbed or pathological conditions (see above). Task evaluation allows a separate characterization of the grip force level and the precision of feedforward processing. Continuous vertical arm movements with a hand-held object have two turning points in a movement cycle that differ clearly in the amount of load which has to be compensated for by an appropriate grip force. At the upper turning point gravitational load is directed downward while inertia is defined by the upward acceleration, i.e., both loads subtract resulting in minimal load force. In contrast at the lower turning point, load is maximal because the inertial load is added to the gravitational force since both now act in the direction of gravity.
On the basis of the results from studies of spontaneous motor tempi we hypothesized a time-of-day effect on the arm movement which should be accompanied by changes in grip force. Faster arm movements in the evening would cause an increase in load that may be counteracted by an increased grip force and vice versa in the morning. Independent of the arm movement, the results of studies on grip force control during manipulative tasks suggest a modulation of grip force at different times of day. Due to the robustness of the grip force load force coupling in healthy subjects during disturbances, we expected the temporal precision between grip and load force to be unaffected by time-of-day effects.
Methods
Subjects
Fourteen healthy subjects (seven females and seven males; aged 23–63 years, mean (SD) age 34.0 (12.2) years) volunteered to participate in this study. Subjects used their dominant hand to grasp and move an instrumented object (two subjects were left-handed, 12 subjects were right-handed). To ensure that all of the subjects were of average chronotypes, they were selected on the basis of their mid-sleep using the Munich CronoType Questionnaire (MCTQ, score 3–5; Roenneberg et al. 2003). None of the subjects participated in shift work or transmeridian travel 1 month prior to the study. Informed consent was obtained from all subjects. The study was conducted in accordance with the Declaration of Helsinki and was approved by an ethical committee.
Instrumented object
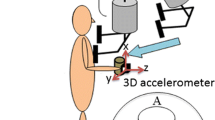
The cylindrical object had a diameter of 9 cm and a depth of 4 cm, the mass of the object was 0.35 kg. During data registration the object was completely autonomous with no connection to external devices. Subjects grasped the object between the thumb and the other fingers in opposition at the circular grip surfaces (see Fig. 1a), which were covered with medium grain sandpaper (no. 240). The object contained a force sensor to measure grip force and three acceleration sensors for registration of acceleration signals in three spatial dimensions (Fig. 1a). By means of internal electronics, the data were analog-to-digital converted with a sampling rate of 100 Hz and stored within the object. Data were transferred to a personal computer for analysis following three experimental sessions.

a Instrumented object containing sensors to measure grip force (GF) and object accelerations in three spatial dimensions (X-, Y-, and Z-axes). The load force (LF) was calculated from the product of the objects mass (m = 0.35 kg) and the vectorial summation of gravity (G) and kinematic accelerations along the object’s Y and Z-axis. b Vertical kinematic acceleration (ACC), load force (LF) and grip force (GF) profiles as well as the force ratio (GF/LF) for continuous vertical arm movements with the hand-held object
Experimental procedure
Subjects were instructed to maintain a regular sleep–wake cycle (within ± 30 min of habitual bed and wake time) during the week of the study, which was verified by sleep diaries. The time-of-day protocol lasted 26 h (Thursday 12:00 h until Friday 14:00 h, see Fig. 2). Subjects tested themselves at home and work. At night subjects slept at home at their habitual bed and wake times. The interval of the sessions was 2 h during the day. In the evening the interval was shortened to 1 h and during the Friday morning sessions were conducted every 30 min, because performance at these hours was expected to show most time-of-day variations (Fig. 2).

Overview of the time-of-day protocol design. Fifteen sessions were carried out over 26 h with an intersession interval of 2 h during the day, 1 h in the evening and 30 min in the morning. During the night, subjects slept at home at their normal bed and waking times
At the beginning of each session subjects rated their current sleepiness from 1 (extremely alert) to 9 (extremely sleepy–fighting sleep) with the Karolinska Sleepiness Scale (KSS; Akerstedt and Gillberg 1990). Subsequently subjects performed cyclic vertical movements with the instrumented object in a sitting position. The grip surfaces were oriented vertically and in parallel to the trunk. Subjects were instructed to continuously move the grasped object in a straight vertical line and to keep this orientation constant during the movement. Movement amplitude was approximately 30 cm. Correct movement execution was trained during practice trials the day before the experiment. Subjects were told to move the object in a spontaneous self paced tempo at a moderately fast and comfortable rate at about 1 Hz. Three trials lasting approximately 10 s were performed in each session. The minimum grip force necessary to prevent the object from slipping (slip force) was assessed by slowly releasing the grip of the hand-held object until the object slipped from the fingers (Johansson and Westling 1984). The slip force was measured directly after the cyclic vertical movements task to control for changes in friction by sweating, washed or dirty hands on the grip force applied. The grip force at the moment of the initial detectable change in acceleration along the object’s Z-axis was taken as slip force. The average value of three attempts was calculated.
Data processing and analysis
The acceleration measured along the object’s Z-axis was a combination of gravity (9.81 m/s2) and movement-induced acceleration. Positive acceleration of the object was directed upwards. Acceleration was maximal at the lower turning point of the vertical cyclic movements. Kinematic acceleration in the direction of movement (ACC, Fig. 1b) was calculated by subtracting the gravitational acceleration from the total acceleration. The net load force (LF Fig. 1a) was calculated as the vector sum of gravitational and inertial load components in the vertical direction along the object’s Z-axis and additionally minor components in the transverse (Y-) direction: LF = SQRT[(G + ACCZ)² + ACCY²]. Loading components acting in the direction of the applied grip forces (X-axis, compare Fig. 1) were not included in the calculation of the objects net load, because these loads did not need to be compensated for by grip force to prevent the object from slipping.
For each trial, an interval containing eight full cycles was selected for further analysis. Performance variables were searched by means of computer algorithms, which first identified maxima and minima of kinematic acceleration (ACC). The algorithm then determined maximum and minimum peaks of load and grip force in windows around the peaks in the acceleration trace (for details see Hermsdörfer et al. 2000). Results are presented for maximum kinematic acceleration (ACC) and the level of grip force, which is the mean grip force in the interval. To evaluate the scaling of the grip force in relation to the load, the ratio of grip force to load force was calculated. The minimum of the ratio (force ratio), which corresponds to the time points when load and grip force are typically maximal at the lower turning point (see Fig. 1b), was determined for each cycle. Maxima of kinematic accelerations and minima of force ratios were averaged across trials. To describe the temporospatial precision of the modulation of the grip force profile with the load force profile, cross correlation analyses were computed between the two forces within each interval containing eight full cycles. The maximum coefficient of cross-correlation is a measure of similarity of both force profiles and the corresponding time lag is a measure of synchronicity (see Hermsdörfer et al. 2000).
The results from the first two sessions were not included in the analyses and were considered as practice sessions. For all statistical comparisons the alpha level was set at 0.05. To examine a time-of-day effect, analyses of variance (ANOVAs) for repeated measures with the factor session were carried out for each performance variable. Diurnal variations were analyzed with eight sessions. Detailed analyses examined performance in the evening until sleep and in the morning after awakening. The violation of the sphericity assumption using repeated measures ANOVA was corrected by the Huynh-Feldt correction. Post hoc contrasts included paired t tests. For each subject data of all performance variables were transformed into the relative performance (in percent) from the individual mean of sessions 3–15 (x relative = 100 × [(x − M x session 3–15)/M x session 3–15]), in order to account for interindividual differences of the absolute measures (cf. Stolz et al. 1988). In an analysis with the absolute values comparable results were found. Effects of the acceleration were partly less clear, probably due to high interindividual variability of movement execution. Missing data during single sessions, resulting from missed sessions and technical failures, led to different sample sizes for the three analyses (diurnal variations N = 8, evening performance N = 13, sleep inertia N = 8). However, all of them were part of the overall sample of 14 subjects. For each performance variable only trial data that lie within 2.5 standard deviations (SD) of the individual daily mean were included in the statistical analyses (0–3.3% data excluded).
Results
Diurnal variations
Diurnal variations were examined with eight sessions during day time assessed in an interval of 2 h, starting at Thursday 16:00 h. Data of eight subjects were analyzed. The arm movement (ACC, Fig. 3a) exhibited a significant effect of session F(7,49) = −3.29, P < 0.05. Post hoc tests revealed a significant increase of acceleration from the evening (18:00 and 20:00 h) till the next morning (8:00 h) and a further significant speeding up of arm movement till 14:00 h. The mean diurnal acceleration of arm movement was 18.1 (2.1) m/s2 and the range of the relative diurnal change was 28.8% (Fig. 3a). The level of grip force (Fig. 3b) showed a trend of session, F(7,49) = 2.05, P = 0.068, with a range of variation of 19.6%. This was due to a relaxation of grip force from the late afternoon (16:00 and 18:00 h) to the evening (20:00 h) followed by an increase till the next morning (8:00 h). The tendency in the diurnal variation in grip force could probably not be attributed to changes in movement acceleration (resulting in fluctuations of load), since in Fig. 3b the course of the ratio between grip force and load varies quite similar to that of grip force. This suggests a diurnal variation of the force ratio, which was however, not supported statistically, F(7,49) < 1 (0.7 ± 0.03), so that the notion of a similar change of grip force and ratio has still to be considered with care. Relative diurnal changes of the force ratio were 9.0%. The mean (SD) diurnal level of grip force was 4.4 (0.3) N and the minimum grip force necessary to hold the object (slip force) was on average 1.6 (0.1) N. Slip force (Fig. 3b) tended to increase over the sessions, F(7,49) = 2.08, P = 0.064, with 9.3% diurnal change. Therefore the modest diurnal decreases and increases in the level of grip force could no be explained by concomitant changes in the risk of slip, indeed the changes of slip force across sessions frequently were in the opposite direction than changes of grip force (see Fig. 3b). The temporospatial relationship between the two force profiles as quantified by the maximum coefficient of the cross-correlation (r corr, Fig. 3c) was very high r = 0.92 (0.01) and did not reveal any diurnal variation F(7,49) = 1.71, P = 0.163. The synchronicity of the two profiles was obvious from a negligible time lag (9 ± 2 m/s) and a high stability over time, F(7,49) < 1 (tlag, Fig. 3c). The subjective sleepiness (KSS) significantly changed during the day F(7,42) = 6.48, P < 0.01 as shown in Fig. 3d. Post hoc tests found a significant increase in sleepiness till bedtime, while the next morning sleepiness decreased significantly and remained constant during lunchtime.

Diurnal variations in sensorimotor performance and subjective sleepiness (interval 2 h). Relative performance represents the percentage deviation from the individual mean of sessions 3–15. Bars indicate the standard error of the mean. a Maximum acceleration along the object’s Z-axis (ACC). b The level of grip force, the ratio of grip force to load, minimum grip force necessary to prevent slipping (slip force). c Maximum coefficient of cross-correlation (r corr) and corresponding time lag between grip and load force (tlag). d Subjective sleepiness (KSS, Karolinska Sleepiness Scale)
Evening performance
The detailed analysis of the performance during the evening in 13 subjects contained the last four sessions before sleep, assessed every hour. Given that bed times of seven of the subjects were after 24:00 h, while the remaining six subjects slept earlier, sessions in the analysis were synchronized to the individual “directly before sleep” session (23:00/24:00 h). The mean evening acceleration of arm movement was 16.8 (0.8) m/s2. During the evening until bed time the maximum kinematic acceleration changed significantly (F(3,36) = 4.19, P < 0.05; Fig. 4a), with a range of 23.6%. Post hoc tests revealed that the arm movement significantly slowed down until 2 h before sleep (21:00/22:00 h) and subsequently accelerated significantly until sleep. In contrast, neither level of grip force nor slip force changed in the course of the evening (grip force: F(3,36) < 1, range of relative grip force 3.7%; slip force F(3,36) = 1.24, P = 0.311; Fig. 4b). The mean level of grip force was 6.5 (0.1) N and the slip force was on average 1.8 (0.5) N. The ratio between grip force and load remained uninfluenced by evening time as well, F(3,36) < 1 (Fig. 4b). Likewise the similarity of both force profiles expressed by the maximum coefficient of cross-correlation (r = 0.82 ± 0.02) and the corresponding time lag (1 ± 3 m/s) as a measure of synchronicity were stable in the course of the evening (r corr F(3,36) = 2.03, P = 0.164; tlag F(3,36) < 1; Fig. 4c). As expected subjective sleepiness increased significantly until bed time, F(3,36) = 19.56, P < 0.001 (Fig. 4d). All post hoc comparisons of the sleepiness ratings at the evening sessions significantly differed.

Sensorimotor performance and subjective sleepiness in the evening (1 h interval). For further details see Fig. 3
Sleep inertia
To examine grip force adjustment in the morning after night’s sleep, the last session before sleep and the first three sessions on Friday morning after awakening (30 min interval) were analyzed. Eight subjects were included in this analysis. The reference of the first session in the morning was the individual wake-time of the subjects. The morning acceleration (20.3 ± 1.0 m/s2, range 19.7%) exhibited a trend (F(3,21) = 3.03, P = 0.052; Fig. 5a) which was due to an increase in acceleration being awake for 30 min and 1 h, respectively. Grip force increased significantly, F(3,21) = 7.04, P < .010, with a range of 15.3% relative grip force. Average grip force in the morning was 7.8 (0.6) N. Post hoc test revealed this being due to an increase from the awakening till being awake for 30 min and 1 h, respectively (Fig. 5b). The ratio of grip to load force (0.92 ± 0.05) also showed a significant influence of morning sessions, F(3,21) = 6.21, P < 0.010, suggesting that grip force changed independent from load within the first hour in the morning (Fig. 5b). The range of the relative force ratio in the morning was 11.9%. The minimum grip force necessary to prevent slipping was 1.6 (0.5) N and was uninfluenced by morning time, F(3,21) < 1 (Fig. 5b) confirming that morning variations in grip force were not due varying slip force. The coupling between the two forces was very precise (r = 0.79 ± 0.01, time lag 5 ± 8 m/s) and did not change across sessions (r corr F(3,21) < 1; tlag F(3,21) = 1.62, P = 0.234; Fig. 5c). Subjective sleepiness in the morning decreased significantly F(3,21) = 17.21, P < 0.010, validated by all post hoc comparisons in the morning being significant (Fig. 5d).

Sensorimotor performance and subjeceltive sleepiness as a function of sleep inertia (interval 30 min). For further details see Fig. 3
Discussion
In the present study, time-of-day effects on grip force control during object manipulation were investigated. On the one hand, time of day influenced the acceleration of arm movement and the level of grip force in particular in the late afternoon and in the morning after awakening. On the other hand, time-of-day did not affect the precise anticipatory adjustment of the grip force profile to the movement-induced fluctuations of the load force profile at any session. Subjective sleepiness revealed a characteristic time-of-day rhythm with increasing sleepiness until bed time and increasing alertness during the next morning.
In accordance with the finding for the spontaneous tapping tempo (Moussay et al. 2002), the arm movement slowed down during the evening till 21:00/22:00 h (2 h before sleep). This slowing may be related to a gradual increase of melatonin secretion that starts approximately at the same time, 2–3 h before the decision to go to bed (Wehr et al. 2001).
Arm acceleration during the last sessions on the second day (12:00 and 14:00 h) was of considerable larger amplitude than during the 16:00 h-session on the first day. However, such a fast increase due to time-of-day effects seems rather unlikely. It is rather possible that subjects anticipated the end of the measurements during the last two sessions of the second day and therefore increased the acceleration of arm movement during these sessions. Remarkably grip force did not show such a tendency towards the end of the second days’ sessions.
The finding that grip force increased during the morning, corresponds with an increase in maximum grip strength in the morning (Wright 1959). However, a continuing increase in grip strength till the maximum in the early evening, shown by Stolz et al. (1988), was not reflected in our grip force data. The fact that maximum grip strength is deliberately produced with maximum effort, while grip forces during object manipulation are highly automatized adjustments to changes in load of much lower magnitude, may be responsible for the difference. Diurnal grip force variations were only present as a statistical trend. The evening performance in grip force (the last 3 h before sleep) revealed no statistically significant change, probably because grip force has already decreased in the late afternoon until 20:00 h and subsequently remained stable. In the morning, grip force changed significantly with an amount of 15%.
A behavioural consequence of low grip forces early after awakening may be accidental slips of objects that is a common experience in everyday life. Thus, the changes in grip force may be relevant, in particularly, for activities which need a high degree of fine motor skill and precise control of grip forces.
It has to be noted however, that grip force was always higher than the minimum force required to avoid object slip. In addition, the observed change of the magnitude of grip force was still much smaller compared to the increases of grip forces due to impaired sensibility following digit cooling (Nowak and Hermsdörfer 2003) or in patients with sensory impairments (Hermsdörfer et al. 2003).
The scaling of grip force in relation to load, assessed by the force ratio, changed similarly to grip force. This was statistically confirmed for the morning sessions: grip force and the force ratio changed significantly, suggesting that grip force changes were not a pure consequence of changes of arm acceleration. In the later case the ratio would have been constant. It can therefore be assumed, that the scaling of grip force changed with time-of-day independently from variations of the arm movement. In contrast, scaling of the grip force with varying loads has repeatedly been demonstrated with explicit load changes such variations of movement frequency (Flanagan and Wing 1995) or gravity changes (Hermsdörfer et al. 2000).
In his book about sleep and wakefulness Kleitman (1963) commented “immediately after getting up, irrespective of the hour, one is not at one’s best” (p. 124). This phenomenon of impaired performance upon awakening that improves asymptotically within minutes to hours depending on the task (Ferrara and De Gennaro 2000), is known as sleep inertia. Cognitive performance and subjective alertness rose rapidly in the first hour after awakening under constant routine conditions as well as on normal scheduled days (ambulatory condition; Jewett et al. 1999). The present results correspond with an increase of grip strength and arm movement acceleration already within 30 min after awakening. However, a time-of-day protocol does not allow distinguishing variations in performance due to endogenous circadian factors from those linked to sleep inertia per se. Human circadian performance is influenced by three major processes: the homeostatic process (homeostatic increase of sleepiness depending on time spent awake), the circadian process (driven by the circadian pacemaker) and the process of sleep inertia (for a review see Blatter and Cajochen 2007). Time-of-day recordings provide a first clue of diurnal variations in performance without knowledge of the underlying processes. The appropriate protocol to study sleep inertia would be a multiple nap protocol with naps at different times of a day. There is clear evidence that sleep deprivation impairs clinical performance in medical professionals (for a review see Gaba and Howard 2002). However, fine motor performance might also be affected by sleep in terms of sleep inertia, e.g., this may be highly relevant for a physician on night call.
The observed pattern of a simultaneously increase and decrease of grip force with load during continuous arm movements with a hand-held object is consistent with previous studies (Flanagan et al. 1993; Flanagan and Tresilian 1994; Flanagan and Wing 1995). This close temporal coupling between grip and load force profiles implies an anticipatory modulation of grip force with changes in load. Flanagan and Wing (1997) suggested that feedforward control is based on an internal forward model, which predicts the physical consequences of our actions enabling a synchronous anticipation of self-produced loads. The coordination of grip and load force involves both a feedforward control and a feedback mechanism, whereas the latter is important to adapt the predictions to the actual requirements (Flanagan and Johansson 2002; Kawato et al. 2003). The present study suggested feedforward predictions to be uninfluenced by time-of-day effects, which is in line with previous findings of a robust anticipatory grip force control regardless of changes in environmental conditions (Hermsdörfer et al. 2000) or sensory impairments (Hermsdörfer et al. 2003; Nowak and Hermsdörfer 2003). However, since the null results are based on relatively low number of subjects in two of the analyses, the notion of a maintained coupling between grip and load force has to be considered with care.
In conclusion, during spontaneous movements with a hand-held object, the level of grip force and the acceleration of arm movement changed with time-of-day. In particular in the morning, while the body re-activates after the night’s sleep, grip force changes particularly obvious and independent of physical demands given by acceleration. However, time-of-day did not influence the predictive temporal coupling between grip and load force profiles evidenced by a high similarity and synchronicity of the two force profiles over time.
References
Akerstedt T, Gillberg M (1990) Subjective and objective sleepiness in the active individual. Int J Neurosci 52:29–37
Atkinson G, Coldwells A, Reilly T, Waterhouse J (1993) A comparison of circadian rhythms in work performance between physically active and inactive subjects. Ergonomics 36:273–281
Blatter K, Cajochen C (2007) Circadian rhythms in cognitive performance: methodological constraints, protocols, theoretical underpinnings. Physiol Behav 90:196–208
Dosseville F, Moussay S, Larue J, Gauthier A, Davenne D (2002) Physical exercise and time of day: influences on spontaneous motor tempo. Percept Mot Skills 95:965–972
Ferrara M, De Gennaro L (2000) The sleep inertia phenomenon during the sleep–wake transition: theoretical and operational issues. Aviat Space Environ Med 71:843–848
Flanagan JR, Johansson RS (2002) Hand movements. In: Ramshandran VS (ed) Encyclopedia of the human brain. Academic, San Diego, pp 399–414
Flanagan JR, Tresilian JR (1994) Grip-load force coupling: a general control strategy for transporting objects. J Exp Psychol Hum Percept Perform 20:944–957
Flanagan JR, Wing AM (1993) Modulation of grip force with load force during point-to-point arm movements. Exp Brain Res 95:131–143
Flanagan JR, Wing AM (1995) The stability of precision grip forces during cyclic arm movements with a hand-held load. Exp Brain Res 105:455–464
Flanagan JR, Wing AM (1997) The role of internal models in motion planning and control: evidence from grip force adjustments during movements of hand-held loads. J Neurosci 17:1519–1528
Flanagan JR, Tresilian J, Wing AM (1993) Coupling of grip force and load force during arm movements with grasped objects. Neurosci Lett 152:53–56
Gaba DM, Howard SK (2002) Patient safety: fatigue among clinicians and the safety of patients. N Engl J Med 347:1249–1255
Hermsdörfer J, Marquardt C, Philipp J, Zierdt A, Nowak D, Glasauer S, Mai N (2000) Moving weightless objects. Grip force control during microgravity. Exp Brain Res 132:52–64
Hermsdörfer J, Hagl E, Nowak DA, Marquardt C (2003) Grip force control during object manipulation in cerebral stroke. Clin Neurophysiol 114:915–929
Ilmarinen J, Ilmarinen R, Korhonen O, Nurminen M (1980) Circadian variation of physiological functions related to physical work capacity. Scand J Work Environ Health 6:112–122
Jeanneret P, Webb W (1963) Strenght of grip on arousal from full night’s sleep. Percept Mot Skills 17:759–761
Jewett ME, Wyatt JK, Ritz-De Cecco A, Khalsa SB, Dijk DJ, Czeisler CA (1999) Time course of sleep inertia dissipation in human performance and alertness. J Sleep Res 8:1–8
Johansson RS (1996) Sensory control of dexterous manipulation in humans. In: Wing A, Haggard P, Flanagan JR (eds) Hand and brain. Academic, San Diego, pp 381–414
Johansson RS, Westling G (1984) Roles of glabrous skin receptors and sensorimotor memory in automatic control of precision grip when lifting rougher or more slippery objects. Exp Brain Res 56:550–564
Kawato M, Kuroda T, Imamizu H, Nakano E, Miyauchi S, Yoshioka T (2003) Internal forward models in the cerebellum: fMRI study on grip force and load force coupling. Prog Brain Res 142:171–188
Kleitman N (1963) Sleep and wakefulness. University of Chicago Press, Chicago
Lawrie SM, MacHale SM, Cavanagh JT, O’Carroll RE, Goodwin GM (2000) The difference in patterns of motor and cognitive function in chronic fatigue syndrome and severe depressive illness. Psychol Med 30:433–442
Moussay S, Dosseville F, Gauthier A, Larue J, Sesboue B, Davenne D (2002) Circadian rhythms during cycling exercise and finger-tapping task. Chronobiol Int 19:1137–1149
Moussay S, Bessot N, Gauthier A, Larue J, Sesboue B, Davenne D (2003) Diurnal variations in cycling kinematics. Chronobiol Int 20:879–892
Nowak DA, Hermsdörfer J (2003) Digit cooling influences grasp efficiency during manipulative tasks. Eur J Appl Physiol 89:127–133
Pearson R, MacKinnon MJ, Meek AP, Myers DB, Palmer DG (1982) Diurnal and sequential grip functions in normal subjects and effects of temperature change and exercise of the forearm on grip function in patients with rheumatoid arthritis and in normal controls. Scand J Rheumatol 11:113–118
Roenneberg T, Wirz-Justice A, Merrow M (2003) Life between clocks: daily temporal patterns of human chronotypes. J Biol Rhythms 18:80–90
Stolz G, Aschoff JC, Born J, Aschoff J (1988) VEP, physiological and psychological circadian variations in humans. J Neurol 235:308–313
Strutton PH, Catley M, Davey NJ (2003) Stability of corticospinal excitability and grip force in intrinsic hand muscles in man over a 24 h period. Physiol Behav 79:679–682
Valdez P, Ramírez C, García A, Talamantes J, Armijo P, Borrani J (2005) Circadian rhythms in components of attention. Biol Rhythm Res 36:57–65
Wehr TA, Aeschbach D, Duncan WC Jr (2001) Evidence for a biological dawn and dusk in the human circadian timing system. J Physiol 535:937–951
Winget CM, DeRoshia CW, Holley DC (1985) Circadian rhythms and athletic performance. Med Sci Sports Exerc 17:498–516
Wright V (1959) Some observations on diurnal variation of grip. Clin Sci (Lond) 18:17–23
Acknowledgments
We wish to thank all subjects sincerely for their participation in the present study which included not only testing at 15 sessions but also maintaining a fixed sleep–wake schedule for 1 week. We also wish to express our sincere thanks to Prof. Anna Wirz-Justice, Prof. Christian Cajochen and Kurt Kräuchi for their support in preparing the protocol. Many thanks to Dr. Yong Li and Andreas Zierdt for their patient help to draw up the figures. This research is part of the Ladenburg Collegium “ClockWork” that is funded by the Gottlieb Daimler- and Karl Benz-Foundation.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Jasper, I., Hermsdörfer, J. Time-of-day effects on force control during object manipulation. Eur J Appl Physiol 101, 437–444 (2007). https://doi.org/10.1007/s00421-007-0514-8
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-007-0514-8