
Abstract
The time course of muscle oxygen desaturation (StO2 kinetics) following exercise onset reflects the dynamic interaction between muscle blood flow and muscle oxygen consumption. In patients with peripheral arterial disease (PAD), muscle StO2 kinetics are slowed during walking exercise; potentially reflecting altered muscle oxygen consumption relative to blood flow. This study evaluated whether StO2 kinetics measured using near infrared spectroscopy (NIRS) would be slowed in PAD during low work rate calf exercise compared with healthy subjects under conditions in which blood flow did not differ. Eight subjects with PAD and eight controls performed 3 min of calf exercise at 5, 10, 30, and 50% of maximal voluntary contraction (MVC). Calf blood flow responses were measured by plethysmography. Power outputs were similar between groups for all work rates. In PAD, the time constants of StO2 kinetics were significantly slower than controls during 5% MVC (13.5 ± 1.7 vs. 6.9 ± 1.2 s, P < 0.05) and 10% MVC work rates (14.5 ± 2.7 vs. 6.8 ± 1.1 s, P < 0.05). Blood flow assessed when exercise was interrupted after 30 s did not differ between PAD and control subjects at these work rates. In contrast, the StO2 time constants were not different between groups during 30 and 50% MVC work rates, where blood flow responses in PAD subjects were lower as compared with controls. Thus in PAD, the slowed StO2 kinetic responses under conditions of unimpaired calf blood flow reflect slowed muscle oxygen consumption in PAD skeletal muscle during low work rate plantar flexion exercise as compared with healthy skeletal muscle.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Individuals with peripheral arterial disease (PAD) have arterial stenoses that may limit leg blood flow responses during exercise at high, but not low, muscle work rates (Pernow and Zetterquist 1968; Sorlie and Myhre 1978). However, there are abnormalities of oxidative metabolism in PAD-affected skeletal muscle that may additionally impair oxygen consumption (Brass et al. 2001; Pipinos et al. 2003). With exercise, it has been demonstrated that systemic oxygen uptake kinetics are slowed in PAD following the onset of walking exercise (Barker et al. 2004; Bauer et al. 1999, 2004a). These systemic findings are consistent with an impaired ability to increase muscle blood flow, muscle oxygen consumption, or both following the onset of exercise. However, our understanding of the interaction between muscle blood flow and oxygen consumption during exercise at the level of the PAD skeletal muscle remains incomplete.
The balance between muscle oxygen consumption and muscle blood flow is reflected in net muscle oxygen extraction. With muscular contractions, muscle oxygen partial pressure (PO2) and microvascular PO2 decrease, of which changes in hemoglobin oxygen saturation can be measured by the absorbance of oxygen-binding proteins. Across the rest to exercise transition, modeling studies have suggested that the time course (i.e. kinetics) of exercise-induced changes in venous oxygen content may indicate differences in the dynamic balance of muscle oxygen delivery relative to muscle oxygen consumption (Ferreira et al. 2005). Accordingly, evaluation of the kinetics of muscle hemoglobin desaturation following exercise onset in PAD could provide insight into the potential influences of muscle blood flow and oxygen consumption in diseased compared with healthy muscle.
A non-invasive surrogate for muscle oxygen extraction is the assessment of tissue oxygen saturation (StO2) using Near Infrared Spectroscopy (NIRS) (MacDonald et al. 1999; Myers et al. 2005; Quaresima et al. 2001). In perfused muscle, StO2 may be considered conceptually similar to venous hemoglobin oxygen saturation under varied conditions (Esaki et al. 2005; Myers et al. 2005). Thus, it is reasonable that the measured time course of change in StO2 with exercise may parallel changes in the balance of muscle oxygen consumption relative to muscle blood flow. Previously, we observed slowed muscle StO2 kinetics in PAD patients following the onset of treadmill exercise compared with controls (Bauer et al. 2004a). If blood flow was the primary variable limiting PAD muscle oxygen consumption, the expected StO2 response would be more rapid kinetics of oxygen extraction to meet the metabolic demand following exercise onset. In contrast, at very low exercise work rates the initial blood flow responses in PAD may not be impaired, providing conditions that would allow for a comparison of the changes in StO2 and inferred oxygen extraction dynamics between PAD and control subjects. In previous studies, neither the work performed nor the blood flow response could be controlled or measured. Therefore, the aim of the present study was to evaluate StO2 kinetics following the onset of exercise in patients with PAD and healthy controls during isolated, single-leg plantar flexions incorporating conditions in which the anticipated blood flow response would be similar in the two groups. We tested the hypothesis that muscle StO2 kinetics would be slowed at low work rates in PAD compared with healthy controls when blood flow responses to these exercise intensities were preserved.
Methods
Subjects
Eight male subjects with PAD and eight healthy men without clinical cardiopulmonary disease were identified prospectively and enrolled in this study. All subjects with PAD were symptomatic, and limited by claudication pain, but were able to perform all testing procedures. Peripheral arterial disease was defined by a resting ankle-brachial index (ABI) < 0.90 (ABI: the ratio of the highest ankle systolic blood pressure in each leg to the highest brachial systolic pressure). The PAD index leg was defined as the leg with the lowest ABI. Subjects were excluded who had a documented history of coronary artery disease, previous myocardial infarction or coronary revascularization, angina, stroke, congestive heart failure, diabetes mellitus, or were taking beta-receptor blocking agents. All subjects provided informed consent, and the study was approved by the University of Colorado Multiple Institutional Review Board in accordance with the Declaration of Helsinki.
Exercise protocol
All subjects performed an incremental treadmill test for the characterization of peak exercise capacity. Subjects performed four exercise trials across two separate visits. The exercise trials consisted of 3-min calf plantar flexion at 5, 10, 30, and 50% of maximal voluntary contraction (MVC) using a high-resolution dynamometer (±1 Newton) (KinCom, Chattanooga, TN, USA). A subset of eight subjects (four PAD; four controls) returned for additional testing to perform exercise trials at 5% MVC and 10% MVC for characterization of early blood flow responses with low work rate exercise. In the follow-up experiments, plantar flexion contractions were performed for 30 s at 5 and 10% MVC work rates.
The dynamometer was adjusted for isotonic plantar flexion contractions from 0 to 15°. Calf exercise was performed in the supine position with the knee at full extension. Contraction frequency was maintained at 0.5 Hz with the aid of an audible metronome. A recovery period of at least 25 min separated each constant work rate bout with subsequent exercise testing performed when resting calf blood flow and StO2 had returned to pre-exercise values.
Tissue oxygen saturation (StO2)
Tissue oxygen saturation was measured using a continuous-wave, near-infrared, four-wavelength spectrometer (InSpectraTM Model 325, Hutchinson Technology, Inc, Hutchinson, MN, USA) as described in detail (Myers et al. 2005). The device measured the absorption of near-infrared light at four wavelengths (680, 720, 760, and 800 nm) as the log (reference intensity/sample intensity). The resulting relative absorption values were used in the calculation of the second derivative of the absorption versus wavelength curves at 720 and 760 nm. The StO2 value (in the units of percent saturation) was derived using the ratio of the second derivatives at 720 and 760 nm, and an empirically scaled relationship to pure blood hemoglobin samples of known oxygen saturation (Myers et al. 2005). Thus, tissue oxygen saturation (StO2) was determined directly from the absorption of NIR light independent of calculated absolute or relative changes in oxy- and deoxyhemoglobin concentrations. The 2nd derivative method has been shown to be experimentally robust at low and high tissue hemoglobin concentrations and has been validated with hemoglobin oxygen saturation in skeletal muscle using co-oximetry (mean error of ±3% vs. co-oximeter) (Myers et al. 2005).
Prior to each visit, the InSpectra device was calibrated using a light scattering standard (closed cell foam prepared by the manufacturer) and the calculated StO2 measurement verified by comparison with optical references of scattering medium with absorbance properties equivalent to 38 and 90% hemoglobin saturation (coefficient of variation of 2.7 and 0.09% for low and high values, respectively). The InSpectra Tissue Spectrometer provided discrete StO2 measurements at a data acquisition rate of 6 Hz to a dedicated computer.
Optical data were acquired with a probe positioned over the lateral gastrocnemius muscle of the index leg. Anatomic localization of the StO2 probe was determined prior to study and was marked for identical placement for all subsequent visits. The probe was firmly attached to the skin by an adhesive patch that eliminated contamination by ambient light. The StO2 probe had a fixed distance of 25 mm from the LED light source to photodetectors. The spacing between the light source and photodetectors provided light attenuation measurements of tissue depth of approximately 12 mm. Because skin and subcutaneous thickness may influence the NIR measurements through the inclusion of oxygenated blood in these regions (van Beekvelt et al. 2001), skin and subcutaneous tissue thickness was assessed by digital calipers over the sampling site using 2D ultrasound (VingMed system 5, GE Milwaukee, WI, USA).
Calf blood flow
Calf blood flow (ml/100 ml tissue/min) was measured in the supine position by venous occlusion strain gauge plethysmography (D.E. Hokanson Inc. Issaquah, WA, USA) using methods previously described (Bauer et al. 2004b). Resting blood flow was calculated as the average of five separate measurements with a coefficient of variation of 7% for repeated measurements. Post-exercise blood flows are presented as individual readings, determined as the first flow immediately following the last contraction.
To estimate the dynamics of the blood flow response, subjects willing to participate returned to the laboratory for another series of plantar exercise sessions at 5 and 10% MVC identical to those above, except that the exercise was stopped after 30 s to allow an estimate of calf blood flow using plethysmography at that time point. This application of plethysmography to assess blood flow during calf exercise has previously been reported (Egana and Green 2005).
Peak oxygen uptake and arterial oxygen saturation
For subject characterization, peak oxygen uptake was measured using pulmonary gas exchange during graded treadmill exercise testing using a metabolic measurement system (CPX/D, Medical Graphics Corporation, St. Paul, MN, USA) (Bauer et al. 1999). Arterial hemoglobin saturation was monitored and recorded during rest, exercise, and recovery of all experiments by an oximeter placed on the index finger of the dominant hand (Ohmeda Corp., Louisville, CO, USA).
Data analysis
The raw StO2 data for each constant work rate test were averaged into 1-s time bins for data smoothing and subsequent data processing using a custom software program (NIRPro, University of Colorado Health Sciences Center, Denver, CO, USA). The StO2 data were curve-fit to a mono-exponential plus delay mathematical model using an iterative least-squares technique by means of a commercial graphing/analysis program (Sigmaplot 2002) as previously described (Bauer et al. 2004a).
For the individual StO2 responses, StO2(b) represents the resting StO2 baseline value. TD1 is the time delay parameter from the onset of exercise to the point of StO2 decrease below baseline as determined by the computer algorithm, τ1 is the time constant, and A 1 is the total decrease in StO2 from baseline to steady state.
Statistical analysis
Group differences were calculated using Student’s t test. The Pearson’s R coefficient was used to evaluate significant correlations. Statistical significance was accepted as P < 0.05 (NCSS 2000-Statistical Software, Kaysville, UT, USA). Data are presented as mean ± SEM.
Results
There were no differences between control and PAD subjects in age or other demographics (Table 1). Four PAD subjects were taking medications indicated for the treatment of hypertension, and two control subjects were treated with statin medications. The resting ABI of the index leg and peak treadmill oxygen uptake in PAD subjects were reduced compared with controls (P < 0.01). Peak treadmill exercise was claudication-limited in all PAD subjects; however, there were no differences between groups for calf plantar flexion MVC. Resting arterial hemoglobin saturation was 94% or higher in all subjects and revealed no changes in any subject during graded or constant work rate exercise testing. Skin and subcutaneous tissue thickness over the region of interest for PAD and control subjects were not different between groups (5.76 ± 0.82 and 5.72 ± 1.16 mm, respectively).
StO2 kinetics
The patterns of StO2 responses during exercise were qualitatively similar between PAD and control subjects (Fig. 1). In both groups the StO2 responses were characterized by a time delay following the onset of exercise, with a subsequent decrease in StO2 which reached a new steady state after 20–40 s. The magnitude of the maximal decrement in StO2 from baseline to steady state increased with increasing work rate.

StO2 responses from a control subject (a) and patient with peripheral arterial disease (b) in the transition from rest to exercise across constant work rates. Exercise time begins at time = 0
The StO2 time constants were slower at 5 and 10% MVC exercise in the PAD subjects compared with controls (P < 0.01 and P < 0.05, respectively Table 2, Fig. 2). The StO2 time constant was independent of work rate in controls, but decreased at work rates above 10% MVC in PAD subjects. As a result, no differences were observed between groups for the StO2 time constants at 30 or 50% MVC exercise. The change in StO2 time constant between low and high work rate exercise (10% MVC–50% MVC) was significantly greater in PAD subjects than controls (−7.7 ± 2.8 vs. 1.6 ± 1.8 s, respectively, P < 0.01). An inverse correlation was observed between the time constant of StO2 during 10% MVC exercise and peak oxygen uptake (ml/min) in PAD subjects (R = −0.88, P < 0.01). The amplitude of StO2 decrease (A1) was similar between groups for all work rates except at 10% MVC exercise where the amplitude parameter was greater in the PAD group compared with controls (P < 0.05).

Representative StO2 kinetics during 10% MVC calf plantar flexion exercise in a control subject (a) and patient with peripheral arterial disease (b). Exercise begins at time = 0. Following a time delay, StO2 falls to steady state with a time constant (τ1). Triangles denote curve fit. The time course of total hemoglobin concentration [tHb] is also shown
Calf blood flow responses to plantar flexion
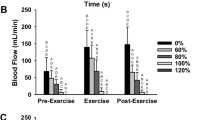
Resting blood flows were not different between PAD and control subjects (Fig. 3). Calf blood flows were also not different after 30 s of 5 and 10% MVC exercise in PAD subjects versus controls (P = 0.64 and P = 0.23, at 5 and 10% MVC, respectively). In contrast, post-exercise blood flow was significantly reduced in the PAD subjects compared with controls at 30% (P < 0.05) and 50% MVC exercise (P < 0.01) but not at lower work rates (Fig. 3).

Comparison of calf blood flow (CBF) in peripheral arterial disease (PAD) patients and control subjects measured at rest and immediately following the last contraction at each work rate. Thirty second CBF data are from four PAD and four controls assessed following independent exercise trials lasting 30 s. All other data represent mean ± SE of eight subjects per group. *P < 0.05, **P < 0.01 PAD vs. control
Discussion
The on-kinetics of muscle oxygen desaturation were slowed in PAD at work rates associated with increases in blood flow not different from responses in controls. During higher work rates and apparent blood flow limitation, muscle oxygen desaturation became more rapid in PAD subjects. Since the changes in muscle oxygen saturation reflect the balance of muscle blood flow relative to oxygen consumption, the slowed time course of muscle oxygen desaturation following the onset of light and moderate exercise indicates an impairment to increase oxygen consumption in PAD-affected muscle early in exercise compared with healthy skeletal muscle.
Blood flow and dynamic calf exercise
Previous studies using NIRS have reported both slowed and accelerated muscle oxygen extraction responses in PAD during various forms and intensities of exercise (Bauer et al. 2004a; Kemp et al. 2001). One possibility for this discrepancy may relate to differences in work rates performed, and thus differential limitations of muscle blood flow relative to muscle oxygen demand. Based on these concepts, the present study evaluated both low and high work rates to elicit two unique hemodynamic conditions in the PAD subjects: (1) where PAD blood flow responses were similar to controls at similar power output, and (2) under conditions where PAD blood flow was reduced relative to the control subjects. This paradigm was based upon empirical observations in PAD which suggested near-normal blood flow increases with low levels of exercise (Bernink et al. 1982; Green 2002; Pernow et al. 1975; Sorlie and Myhre 1978) and impaired leg blood flow responses at higher exercise work rates due to the peripheral arterial occlusive disease (Pernow et al. 1975; Sorlie and Myhre 1978). The present study (using plethysmography to estimate the early and end-exercise blood flow) was able to achieve hemodynamic responses in PAD that were similar to controls at low work rates and reduced at high work rates (Fig. 3).
Differential StO2 kinetics in PAD versus controls
Using computer simulations, Ferreira et al. (2005) demonstrated that in healthy muscle, the kinetics of oxygen extraction following exercise onset is rapid and determined by the dynamic relation between muscle blood flow and oxygen consumption during exercise. In the present study, early calf blood flow responses were similar in PAD and control subjects during low work rate exercise, yet the time course of muscle oxygen desaturation was prolonged in PAD muscle. Given the apparent similarities in blood flow responses during low work rates, the slower rate of desaturation is most consistent with a slower dynamic increase of muscle oxygen consumption in the PAD-affected muscle (Ferreira et al. 2005), a finding that is consistent with the abnormalities of oxidative metabolism previously reported in PAD subjects (Brass et al. 2001; Kemp et al. 2002; Pipinos et al. 2003, 2006). For example, specific enzymes of the electron transport chain exhibit decreased activities on a per mitochondrion basis in muscle from PAD patients (Brass et al. 2001). Moreover, in vivo data indicate the initial rate of oxidative ATP synthesis following the onset of exercise is reduced in PAD skeletal muscle, suggesting a role for abnormal control of oxidative metabolism in the muscle of subjects with PAD (Kemp et al. 2002). Damage to skeletal muscle mitochondria and potential impairment of function is consistent with the finding of mitochondrial ischemia-reperfusion injury observed in other models such as myocardium (Lesnefsky and Hoppel 2003). Thus, claudication-induced ischemia followed by reperfusion injury might also be anticipated in the affected skeletal muscle of patients with PAD.
The present data do not indicate that oxygen extraction at steady state was impaired in PAD, as muscle oxygen desaturation was similar or greater in PAD compared with controls. Rather, the slowed PAD StO2 kinetics suggest that the ability of muscle to rapidly increase muscle oxygen consumption was impaired despite adequate blood flow following exercise onset, resulting in the slowed kinetics of muscle desaturation. Similar responses have been previously described by our group during submaximal treadmill exercise. These studies in PAD demonstrated both slowed kinetics of muscle oxygen desaturation (Bauer et al. 2004a) and slowed systemic oxygen uptake (Bauer et al. 1999) following the onset of exercise. However, these responses eventually achieved similar steady state values to controls by the end of exercise. Of note, true maximal oxygen consumption by skeletal muscle mitochondria cannot be assessed in vivo due to the limitation imposed by blood flow and claudication as clearly evidenced in the reduced peak VO2 of the PAD cohort. Importantly, mitochondrial injury leading to a kinetic defect of oxygen uptake (and resulting in a markedly altered intracellular milieu for any given rate of oxygen consumption) may or may not be associated with a defect in maximal oxygen consumption.
Effect of work rate on StO2 kinetics in PAD
The time course of muscle oxygen desaturation was more rapid in PAD subjects during high work rates as compared with the same subjects at lower work rates. Computer modeling suggests the kinetics of oxygen extraction are exquisitely sensitive to perturbations in the balance of muscle blood flow to oxygen consumption such that even a minor slowing and mis-match of blood flow relative to oxygen consumption results in faster oxygen extraction kinetics (Ferreira et al. 2005). In this manner, any impairment of blood flow relative to oxygen demand following the onset of exercise (as was likely the case in PAD subjects at high, but not low work rates) would have the effect of speeding muscle StO2 kinetics. Moreover, observed differences in StO2 kinetics between two populations or exercise conditions will always be the summation of potential differences caused by blood flow and muscle oxygen consumption. Thus, we speculate this could explain the discordance of StO2 kinetics across low and high work rates of the present study as well as the differences in responses observed in the literature under varying exercise and hemodynamic conditions (Bauer et al. 2004a; Kemp et al. 2001).
Implications of altered StO2 kinetics
Slowed kinetics of muscle oxygen uptake in PAD likely result in a greater depletion of phosphocreatine and reliance upon non-oxidative means of ATP synthesis early in exercise (Kemp et al. 2002). The accumulation of metabolites (e.g. H+, Pi, lactate ion) secondary to the obligate substrate-level phosphorylation required to support exercise contractions may have adverse effects on muscle contractile performance (Greenhaff et al. 2002). Indeed, these higher metabolite concentrations may affect muscle performance even if equivalent net oxygen extraction and rates of oxygen consumption are eventually achieved at steady state, or if maximal muscle oxidative capacity, per se, is preserved. The accumulated oxygen deficit incurred following the onset of low work rate exercise could thus contribute to the premature fatigue observed during ambulation in patients with PAD. It is presently unclear to what extent the abnormalities of oxidative metabolism in PAD skeletal muscle contribute to the exercise intolerance of these patients. However, our findings of slowed muscle oxygen desaturation during isolated low work rate exercise in PAD skeletal muscle appear to support the concept that muscle metabolic abnormalities may play a role in the limited exercise function of this patient population.
Limitations
Plethysmography has significant limitations when used to assess dynamic changes in blood flow, as continuous measures of blood flow cannot be obtained during exercise but only after exercise contractions. However, the intermittent nature of the contraction during calf exercise has facilitated the application of plethysmography to assessing blood flow in these conditions (Egana and Green 2005). Further, the quantitative concordance of the PAD blood flow responses obtained in this study with previous invasive studies (Pernow and Zetterquist 1968; Sorlie and Myhre 1978) strongly supports our inferences as to the relative time course and contributions of blood flow relative to muscle oxygen consumption in the StO2 responses observed.
Skin and adipose thickness have been shown to influence measures of muscle oxygenation using NIRS (van Beekvelt et al. 2001). None of our subjects had a skin and subcutaneous adipose thickness equal to or greater than 15 mm, minimizing this effect. Nevertheless, because near infrared light must pass through these superficial, non-active tissue layers, it may be impossible to quantitatively evaluate purely muscle oxygen saturation, per se. Despite this limitation, it was possible to evaluate the relative change and kinetics of estimated muscle oxygen extraction with exercise since the change in tissue oxygen saturation during exercise was fairly large.
Muscle StO2 measures may also be influenced by the absorption spectra of myoglobin and hemoglobin, which are nearly identical yet may have differing kinetics of desaturation (Marcinek et al. 2003; Tran et al. 1999). However, given the sizable difference in myoglobin versus hemoglobin concentrations in perfused skeletal muscle, its contribution is estimated to be relatively small (Mancini et al. 1994; Seiyama et al. 1988). Additionally, our estimates of tissue hemoglobin concentration (tHb) indicated a small decrease in tHb with contractions in all subjects prior to muscle StO2 desaturation. Thus, although the StO2 algorithm has been shown to be robust to changes in [tHb] (Myers et al. 2005), we cannot exclude the possibility that dynamic changes in [tHb] following exercise onset may have affected the StO2 measurements.
Conclusions
Muscle StO2 kinetics were slowed in PAD compared with control subjects during low calf work rates eliciting similar increases in calf blood flow. During higher work rates and reduced blood flow responses, PAD StO2 kinetics were accelerated as compared with low work rates, while the kinetics were work-rate independent in healthy subjects. The results in PAD during low work rate exercise are consistent with an impaired ability to increase muscle oxygen consumption in PAD-affected skeletal muscle following the onset of exercise.
References
Barker GA, Green S, Green AA, Walker PJ (2004) Walking performance, oxygen uptake kinetics and resting muscle pyruvate dehydrogenase complex activity in peripheral arterial disease Clin. Sci (Lond) 106:241–249
Bauer TA, Regensteiner JG, Brass EP, Hiatt WR (1999) Oxygen uptake kinetics during exercise are slowed in patients with peripheral arterial disease. J Appl Physiol 87:809–816
Bauer TA, Brass EP, Hiatt WR (2004a) Impaired muscle oxygen use at onset of exercise in peripheral arterial disease. J Vasc Surg 40:488–493
Bauer TA, Brass EP, Nehler M, Barstow TJ, Hiatt WR (2004b) Pulmonary VO2 dynamics during treadmill and arm exercise in peripheral arterial disease. J Appl Physiol 97:627–634
Bernink PJ, Lubbers J, Barendsen GJ, van den Berg J (1982) Blood flow in the calf during and after exercise: measurements with Doppler ultrasound and venous occlusion plethysmography in healthy subjects and in patients with arterial occlusive disease. Angiology 33:146–160
Brass EP, Hiatt WR, Gardner AW, Hoppel CL (2001) Decreased NADH dehydrogenase and ubiquinol-cytochrome c oxidoreductase in peripheral arterial disease. Am J Physiol 280:H603–H609
Egana M, Green S (2005) Effect of body tilt on calf muscle performance and blood flow in humans. J Appl Physiol 98:2249–2258
Esaki K, Hamaoka T, Radegran G, Boushel R, Hansen J, Katsumura T, Haga S, Mizuno M (2005) Association between regional quadriceps oxygenation and blood oxygen saturation during normoxic one-legged dynamic knee extension. Eur J Appl Physiol 95:361–370
Ferreira LF, Poole DC, Barstow TJ (2005) Muscle blood flow–O(2) uptake interaction and their relation to on-exercise dynamics of O(2) exchange. Respir Physiol Neurobiol 147:91–103
Green S (2002) Haemodynamic limitations and exercise performance in peripheral arterial disease. Clin Physiol Funct Imaging 22:81–91
Greenhaff PL, Campbell-O’Sullivan SP, Constantin-Teodosiu D, Poucher SM, Roberts PA, Timmons JA (2002) An acetyl group deficit limits mitochondrial ATP production at the onset of exercise. Biochem Soc Trans 30:275–280
Kemp GJ, Roberts N, Bimson WE, Bakran A, Harris PL, Gilling-Smith GL, Brennan J, Rankin A, Frostick SP (2001) Mitochondrial function and oxygen supply in normal and in chronically ischemic muscle: a combined 31P magnetic resonance spectroscopy and near infrared spectroscopy study in vivo. J Vasc Surg 34:1103–1110
Kemp GJ, Roberts N, Bimson WE, Bakran A, Frostick SP (2002) Muscle oxygenation and ATP turnover when blood flow is impaired by vascular disease. Mol Biol Rep 29:187–191
Lesnefsky EJ, Hoppel CL (2003) Ischemia-reperfusion injury in the aged heart: role of mitochondria. Arch Biochem Biophys 420:287–297
MacDonald MJ, Tarnopolsky MA, Green HJ, Hughson RL (1999) Comparison of femoral blood gases and muscle near-infrared spectroscopy at exercise onset in humans. J Appl Physiol 86:687–693
Mancini DM, Bolinger L, Li H, Kendrick K, Chance B, Wilson JR (1994) Validation of near-infrared spectroscopy in humans. J Appl Physiol 77:2740–2747
Marcinek DJ, Ciesielski WA, Conley KE, Schenkman KA (2003) Oxygen regulation and limitation to cellular respiration in mouse skeletal muscle in vivo. Am J Physiol Heart Circ Physiol 285:H1900–H1908
Myers DE, Andersen LD, Seifert RP, Ortner JP, Cooper CE, Beilman GJ, Mowlem JD (2005) Noninvasive method for measuring local hemoglobin oxygen saturation in tissue using wide gap second derivative near-infrared spectroscopy. J Biomed Opt 10:034017-1–034017-18
Pernow B, Zetterquist S (1968) Metabolic evaluation of the leg blood flow in claudicating patients with arterial obstructions at different levels. Scand J Clin Lab Invest 21:277–287
Pernow B, Saltin B, Wahren J, Cronestrand R, Ekestroom S (1975) Leg blood flow and muscle metabolism in occlusive arterial disease of the leg before and after reconstructive surgery. Clin Sci Mol Med 49:265–275
Pipinos II, Sharov VG, Shepard AD, Anagnostopoulos PV, Katsamouris A, Todor A, Filis KA, Sabbah HN (2003) Abnormal mitochondrial respiration in skeletal muscle in patients with peripheral arterial disease. J Vasc Surg 38:827–832
Pipinos II, Judge AR, Zhu Z, Selsby JT, Swanson SA, Johanning JM, Baxter BT, Lynch TG, Dodd SL (2006) Mitochondrial defects and oxidative damage in patients with peripheral arterial disease. Free Radic Biol Med 41:262–269
Quaresima V, Homma S, Azuma K, Shimizu S, Chiarotti F, Ferrari M, Kagaya A (2001) Calf and shin muscle oxygenation patterns and femoral artery blood flow during dynamic plantar flexion exercise in humans. Eur J Appl Physiol 84:387–394
Seiyama A, Hazeki O, Tamura M (1988) Noninvasive quantitative analysis of blood oxygenation in rat skeletal muscle. J Biochem (Tokyo) 103:419–424
Sorlie D, Myhre K (1978) Lower leg blood flow in intermittent claudication. Scand J Clin Lab Invest 38:171–179
Tran TK, Sailasuta N, Kreutzer U, Hurd R, Chung Y, Mole P, Kuno S, Jue T (1999) Comparative analysis of NMR and NIRS measurements of intracellular PO2 in human skeletal muscle. Am J Physiol 276:R1682–R1690
van Beekvelt MC, Borghuis MS, van Engelen BG, Wevers RA, Colier WN (2001) Adipose tissue thickness affects in vivo quantitative near-IR spectroscopy in human skeletal muscle. Clin Sci (Lond) 101:21–28
Acknowledgments
This study was supported by the General Clinical Research Center of University of Colorado Hospital (NIH MO1 #RR000051) and Hutchinson Technology, Inc.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Bauer, T.A., Brass, E.P., Barstow, T.J. et al. Skeletal muscle StO2 kinetics are slowed during low work rate calf exercise in peripheral arterial disease. Eur J Appl Physiol 100, 143–151 (2007). https://doi.org/10.1007/s00421-007-0412-0
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-007-0412-0