
Abstract
In this paper a complete energy balance for water locomotion is attempted with the aim of comparing different modes of transport in the aquatic environment (swimming underwater with SCUBA diving equipment, swimming at the surface: leg kicking and front crawl, kayaking and rowing). On the basis of the values of metabolic power (Ė), of the power needed to overcome water resistance (Ẇ d) and of propelling efficiency (η P=Ẇ d/Ẇ tot, where Ẇ tot is the total mechanical power) as reported in the literature for each of these forms of locomotion, the energy cost per unit distance (C=Ė/v, where v is the velocity), the drag (performance) efficiency (η d=Ẇ d/Ė) and the overall efficiency (η o=Ẇ tot/Ė=η d /η P) were calculated. As previously found for human locomotion on land, for a given metabolic power (e.g. 0.5 kW=1.43 l·min−1 V̇O2) the decrease in C (from 0.88 kJ·m−1 in SCUBA diving to 0.22 kJ·m−1 in rowing) is associated with an increase in the speed of locomotion (from 0.6 m·s−1 in SCUBA diving to 2.4 m·s−1 in rowing). At variance with locomotion on land, however, the decrease in C is associated with an increase, rather than a decrease, of the total mechanical work per unit distance (W tot, kJ·m−1). This is made possible by the increase of the overall efficiency of locomotion (η o=Ẇ tot/Ė=W tot /C) from the slow speeds (and loads) of swimming to the high speeds (and loads) attainable with hulls and boats (from 0.10 in SCUBA diving to 0.29 in rowing).
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Water locomotion, although more energy demanding per unit distance than locomotion on land, was used by humans for thousand of years. In addition to swimming freely at the surface (or underwater) humans have learnt to use a variety of passive locomotory devices, from tree trunks and floating objects to human-powered boats and watercrafts in the attempt to improve the economy and/or the speed of progression in water. Even though motorized locomotion has now replaced human-powered locomotion, aquatic activities such as swimming, diving, kayaking and rowing are still largely practicised all over the word mostly, if not exclusively, for recreational purposes.
Because of the complexities of the water environment, aquatic locomotion offers an interesting challenge to the understanding of the physiology of human movement; it is traditionally investigated by means of: (1) a biomechanical approach focused on the study of the mechanical determinants and/or (2) a physiological approach focused on the study of the energy requirements.
Biomechanics
Generally speaking, for a swimmer or a boat to move in still water at constant speed (v), the propulsive force must be equal to the sum of the forces opposing motion, according to the Newtonian principle of action-reaction. These forces are generally grouped together under the term drag (D) whose different components will not be analysed here, the interested reader being referred to Abbott et al. (1995) and Dal Monte and Komor (1989). However, the total power of locomotion (Ẇ tot, W) is greater than that needed to overcome the drag forces only, because a given fraction of it is "wasted" to: (1) accelerate and decelerate the limbs with respect to the body center of mass (Ẇ int, internal power) and (2) accelerate water away from the body (Ẇ k, kinetic power). For aquatic locomotion therefore:
where Ẇ d is the amount of the power dissipated against drag forces (Ẇ d=D·v, where D is in N and v in m·s−1). It should also be noted that it is often convenient to define external power as the sum of Ẇ d and Ẇ k: Ẇ ext=Ẇ d+Ẇ k.
When swimming or paddling, the limb/tool that generates propulsion also creates drag (D). Hence, during aquatic locomotion "active drag" is significantly greater than the passive drag that can be measured by towing the subject (or the boat) motionless through the water. Passive drag is easily measured, whereas active drag can only be estimated indirectly.
For swimming active drag can be calculated from measures of sub-maximal oxygen consumption (V̇O2) while the subjects swim at constant speed and are partially towed by a force acting in the direction of motion. Linear extrapolation of the V̇O2 versus towing force relationship to resting V̇O2 yields the towing force which would allow the subject to swim at the given speed without any (net) energy expenditure; this force is assumed to be equal (and opposite) to the active drag at that speed (e. g. di Prampero et al. 1974). This method can be, and has been, used also to estimate the active drag of other forms of aquatic locomotion (kayaking: Pendergast et al. 1989; SCUBA diving: Pendergast et al. 1996; leg kicking at the surface: Zamparo et al. 2002).
Active drag can also be assessed by means of the MAD (measuring active drag) system (e.g. Toussaint et al. 1988). With this device the subject swims by exerting the propelling action of the arms on fixed pads positioned 1.35 m apart 0.8 m below the water surface. The pads are connected to force transducers so that the force exerted by the swimmer at each "stroke" can be recorded; this force is equal to the drag force since, with this set up, the swimmer does not expend any energy in giving water momentum (Ẇ k=0). It must be pointed out, however, that only arm propulsion can be investigated with this system because the subject's legs are fixed together and supported by a small buoy.
The component of the total mechanical power "wasted" in accelerating water away from the body (Ẇ k) is even harder to assess than the active drag itself. Estimates of the term Ẇ k for the arm stroke are reported by Toussaint et al. (1988, 1991) as obtained by means of the MAD system (see above) and by Berger et al. (1997) by means of cinematic analysis.
Studies of animal locomotion (e.g. Lighthill 1975; Alexander 1983, 2003) have shown that, in slender fish, Ẇ k can be computed by measuring the speed of the waves of bending which can be observed to travel along the fish's body in a caudal direction (e.g. Alexander 1983, 2003; Lighthill 1975). Indeed, the Froude efficiency of an undulatory movement (η F) can be calculated from the ratio [(u+v)/2u] (were u is the speed of the bending wave and v is the forward speed). Since η F is also defined as the ratio Ẇ d /(Ẇ d+Ẇ k) (see Eq. 3 below) it is possible to calculate Ẇ k on the basis of values of η F [e.g. calculated as (u+v)/2u] and Ẇ d. This last method was applied to calculate η F in subjects swimming by using the leg kick (with and without fins) since the action of the leg's muscles indeed produces waves of bending similar to those observed in slender fish (Zamparo et al. 2002). These waves were also observed in subjects swimming the butterfly kick (Sanders et al. 1995) and in monofin swimmers (Zamparo, personal communication).
It goes without saying that, in any form of water locomotion, the term Ẇ k can be estimated once the values of η F (calculated/estimated/assumed) and Ẇ d are known.
In most studies regarding aquatic locomotion the contribution of the internal work rate (Ẇ int) to Ẇ tot has not been considered as a source of mechanical work expenditure; it was recently computed by Zamparo (Zamparo et al. 2002; Zamparo 2003) for the leg kick and the arm stroke. Ẇ int was originally defined by Fenn (1930) as the power necessary to accelerate the limbs in respect to the center of mass; as such it is strongly influenced by the frequency of the limbs movement. As shown by Zamparo et al. (2002) Ẇ int is a substantial fraction of Ẇ tot when swimming by kicking the legs because of the high limb frequencies necessary to produce forward motion. The limb frequency is greatly reduced when leg kicking with fins (Zamparo et al. 2002) and further reduced in the arm stroke (Zamparo 2003) thus leading to a substantial reduction of Ẇ int. As a first approximation, in all forms of water locomotion except the leg kick without fins, Ẇ int can be neglected and Ẇ tot can be assumed to be equal to the sum of Ẇ k and Ẇ d only (see also Discussion).
The efficiency with which Ẇ tot produced by the swimmer/rower/kayaker is transformed into useful propulsion (useful mechanical output, Ẇ d) is termed propelling efficiency and is given by:
The efficiency with which the external mechanical power (Ẇ ext=Ẇ d+Ẇ k) produced by the swimmer/rower/kayaker is transformed into useful propulsion (useful mechanical output, Ẇ d) is termed Froude efficiency and is given by:
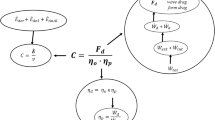
In those forms of locomotion in which the internal work rate can be neglected (see above) Eqs. 2 and 3 are equivalent and η F=η P. From Eq. 2 and 3 is also apparent that, for any form of locomotion in water: (1) the total mechanical power (Ẇ tot=Ẇ d+Ẇ k+Ẇ int) can be calculated rearranging Eq. 2 once the term Ẇ d and the propelling efficiency (η P) are known; and (2) the external mechanical power (Ẇ ext=Ẇ d+Ẇ k) can be calculated rearranging Eq. 3 once the term Ẇ d and the Froude efficiency (η F) are known (see also Scheme 1).

Values of propelling efficiency are reported by Zamparo (2003) for the front crawl, by Toussaint et al. (1988, 1991) and Martin et al. (1980) for the arm stroke, and by Zamparo et al. (2002) for surface kicking (with and without fins). Values of Froude efficiency are reported by Samimy (2002) for underwater kicking with SCUBA and by Zamparo et al. (2002) for surface kicking (with and without fins). To our knowledge, the Froude/propelling efficiency for rowing and kayaking was never measured, essentially because of the difficulties in measuring Ẇ k in these forms of locomotion; however, there is good agreement on the esteems of this parameter, which, according to several authors, ranges from 0.65 to 0.75 (e.g. Celentano et al. 1974; Dal Monte and Komor 1989; Abbott et al. 1995).
Bioenergetics
The net energy cost of locomotion (C, kJ·m−1) can be calculated from the ratio of metabolic power (Ė kW, above resting values) to the speed of progression (v, m·s−1); it represents the net energy expended to cover a unit distance:
The rate of (net) energy expenditure (Ė) is the sum of the rates of oxidative metabolism, anaerobic glycolysis leading to lactic acid production and creatine phosphate splitting. Whereas the energy cost of locomotion is easy to assess over the range of speeds that can be maintained aerobically (in which case Ė=V̇O2), determining C over the speeds where the anaerobic contribution cannot be neglected is a more challenging task, necessarily implying a certain degree of approximation. At maximal and supra-maximal speeds C was estimated for swimming and kayaking by Capelli et al. (1998), Termin and Pendergast (2001), Zamparo et al. (1999, 2000), and Pendergast et al. (1989).
The values of Ė and C reported in this paper are net values (above resting metabolic rate) and refer to a range of speeds that could be supported completely by oxidative metabolism (Ė=V̇O 2). The "aerobic" speed range for each form of locomotion is given in Table 1.
The "metabolic" efficiency of human swimming was originally calculated by di Prampero et al. (1974) by using a method which did not take into account the contribution of the power "wasted" to impart kinetic energy to the water (Ẇ k) nor the internal work rate (Ẇ int) in the computation of Ẇ tot. With this method the efficiency is calculated on the basis of the slope of the V̇O2 vs. D relationship as obtained during the determination of the active drag: ΔD·v/ΔV̇O2 (were both terms are expressed in the same units). The efficiency calculated in this way can be considered as the "efficiency" of overcoming drag only (drag efficiency, η d) since it is given by:
The drag efficiency, as calculated from the ratio Ẇ d/ Ė (Eq. 5), is a useful parameter in aquatic locomotion because it indicates the efficiency with which the overall energy expenditure is utilized to produce "useful" work (thrust). In studies of animal locomotion this efficiency is referred as performance efficiency (Daniel 1991).
About a decade later, Toussaint and coworkers (1988) proposed an alternative method to determine the efficiency of swimming (by means of the MAD system) in which the component of the external power "wasted" to impart kinetic energy to the water (Ẇ k) was also taken into account. The values of efficiency calculated with this method are based on a more complete understanding of aquatic locomotion; these values, however, refer to the arm stroke only (while swimming on the MAD system the legs are fixed together and supported by a pull buoy). Recently, an even more complete energy balance in aquatic locomotion (while swimming the leg kick) was proposed by Zamparo et al. (2002) based on measures/calculations of all the three components of Ẇ tot:
where Ẇ d was computed according to di Prampero et al. (1974), Ẇ int, according to Minetti (1988) and Ẇ k was estimated from measures of Froude efficiency (calculated as proposed by Lighthill 1975, for the slender fish).
By combining Eqs. 5 and 6 it is apparent that the ratio of drag (performance) efficiency to overall efficiency is the propelling efficiency of locomotion (Eq. 3, see also Scheme 1). Hence, the overall efficiency can also be obtained by:
Therefore, to calculate the overall efficiency of any form of aquatic locomotion, only four parameters need to be known: v, Ė, D and η P.
On these premises, this study was carried out with the aim of comparing different forms of locomotion in water. Of all the data collected by the authors over the last three decades, only a limited number of papers were selected. These papers report data obtained while kicking the legs at the surface (L, Zamparo et al. 2002) or underwater (UWF, with SCUBA diving equipment, Samimy 2002), while swimming the front crawl (AL, Zamparo 2003), kayaking (K, Pendergast et al. 1989) and rowing (R, di Prampero et al. 1971; Celentano et al. 1974) and were selected because they report values of energy cost, active drag and cycle frequency as measured, simultaneously, in experienced athletes. These data, once combined with measures/estimates of propelling efficiency, allowed us to attempt a complete energy balance for different forms of human movement in the aquatic environment.
Results
Metabolic power (Ė̇) and energy cost of locomotion (C)
The overall metabolic power (Ė, kW) is plotted in Fig. 1 as a function of the speed for all the investigated forms of aquatic locomotion. The data reported in this and the following figures were obtained by interpolating the original data, over the aerobic speed range, with power functions the equations of which are indicated in the legend to Fig. 1. Ė increases as a function of the speed in all forms of aquatic locomotion and, at comparable (paired) speeds, it is higher for the leg kick (underwater and at the surface), slightly lower for the front crawl, and largely reduced in kayaking and in rowing, this last being the most economical form of locomotion in water.

Net energy expenditure (Ė, kW) is plotted as a function of the speed (v, m·s−1) for the different forms of aquatic locomotion investigated in this study. The lines interpolate the original data over the aerobic speed range. The fitted equations are: L (swimming by using the leg kick at the surface): Ė=0.91·v 1.75; UWF (underwater swimming with SCUBA diving equipment): Ė=1.08·v 1.39; AL (swimming the front crawl): Ė=0.60·v 1.87; K (kayaking): Ė=0.098·v 2.76; R (rowing): Ė=0.088·v 2.05
The energy cost per unit distance (C, kJ·m−1) can be calculated from data reported in Fig. 1 by dividing the energy expenditure by the speed of progression (C=Ė/v, where Ė is in kW and v in m·s−1). C is plotted in Fig. 2 as a function of speed, the continuous lines representing iso-metabolic power hyperbolae of 0.5, 1.0, 1.5 and 2 kW (from bottom to top). As shown in this figure, at the same metabolic power (e.g. 0.5 kW), the decrease in C allows for a fourfold increase in the speed of locomotion: from about 0.6 m·s−1 in SCUBA diving to 2.4 m·s−1 in rowing (see also Table 2).

The energy cost to cover a given distance (C, kJ·m−1) is plotted as a function of the speed (v, m·s−1) for the different forms of aquatic locomotion investigated in this study. (AL Swimming the front crawl, K kayaking, L swimming by using the leg kick at the surface, R rowing, UWF underwater swimming with SCUBA diving equipment.) The continuous lines represent iso-metabolic power hyperbolae of 0.5, 1.0, 1.5 and 2 kW (from bottom to top)
Froude (η F) and propelling (η P) efficiency
Table 1 reports the values of Froude and propelling efficiency as reported in the literature for different forms of aquatic locomotion. In the indicated speed range η P and η F are essentially unaffected by the speed. The following values of η P were utilized in the calculations:
-
1.
η P=0.36 for the leg kick without fins (as measured by Zamparo et al. 2002).
-
2.
η P=0.58 for fin swimming underwater with SCUBA diving equipment (as measured by Samimy 2002. In the study of Samimy only the Froude efficiency was calculated; however, since the η F of swimming underwater is the same as that calculated when swimming at the surface with the same kind of fins (0.72 vs. 0.70), it is fair to assume that also the propelling efficiency is similar.
-
3.
η P=0.45 for the arm stroke; this value is an average of the values of η P as measured by means of the MAD system (e.g. by Toussaint et al. 1988, 1991) or calculated by mathematical modelling (Martin et al. 1980) for the arm stroke and of the values of η P as calculated by Zamparo (2003) for the front crawl with and without fins.
-
4.
η P=0.7 for rowing and kayaking; this value is an average of the (estimated) values of η P reported in the literature (e.g. Dal Monte and Komor 1989; Celentano et al. 1974; Abbott et al. 1995).
The power to overcome drag (Ẇ d), the drag efficiency (η d) and the overall efficiency (η o)
The values of active drag (D, N) are shown in Fig. 3, as a function of the speed, for all forms of locomotion but rowing, in which case the data reported here refer to passive drag. D increases with speed in all forms of locomotion: at comparable (paired) speeds, water resistance is higher when swimming underwater with SCUBA diving equipment, slightly lower for the leg kick and the front crawl and largely reduced when boats are used, the rowing shell offering the lowest resistance to motion in water. Ẇ d (W) can be calculated from the product D·v, were D is in N and v is in m·s−1; drag (performance) efficiency can be calculated from the ratio Ẇ d/ Ė.

Active/passive drag (D, N) is plotted as a function of the speed (v, m·s−1) for the different forms of aquatic locomotion investigated in this study. The lines interpolate the original data over the aerobic speed range. The fitted equations are: L (swimming by using the leg kick at the surface): D=42.9·v 1.55; UWF (underwater swimming with SCUBA diving equipment): D=115.7·v 1.57; AL (swimming the front crawl): D=49.8·v 1.03; K (kayaking): D=14.5·v 2.01; R (rowing): D=12.8·v 1.33
The values of η d are reported in Fig. 4 for all forms of water locomotion considered here. From these values the overall efficiency was calculated by assuming a value of propelling efficiency (η ο=η d/ η P) specific for each form of locomotion, as indicated above.

Drag (performance) efficiency (η d ) is plotted as a function of the speed (v, m·s−1) for the different forms of aquatic locomotion investigated in this study. The lines interpolate the original data over the aerobic speed range. The fitted equations are: L (swimming by using the leg kick at the surface): η d=0.057·v 0.51; UWF (underwater swimming with SCUBA diving equipment): η d=0.108·v 1.17; AL (swimming the front crawl): η d=0.083·v 0.16; K (kayaking) and R (rowing): η d=0.146·v 0.28
The values of η ο are reported in Fig. 5. Both η d and η ο (Figs. 4 and 5) were found to increase with the speed for each form of water locomotion considered in this study. In addition, overall efficiency, as measured at the maximal (aerobic) speeds investigated in this study, was lower for swimming underwater with SCUBA and for swimming at the surface by kicking the legs (about 0.13 on average), corresponding to 0.19 in the front crawl, to 0.27 in kayaking and 0.32 in rowing.

The overall efficiency (η 0=η d /η P) is plotted as a function of the speed (v, m·s−1) for the different forms of aquatic locomotion investigated in this study. (AL Swimming the front crawl, K kayaking, L swimming by using the leg kick at the surface, R rowing, UWF underwater swimming with SCUBA diving equipment.) Data for "boat locomotion" (K and R) could be fitted by the following equation: η 0=0.21·v 0.28, n=30, r 2=0.994; whereas the data for the leg kick (L and UWF) could be fitted by: η 0=0.14·v 0.80, n=15, r 2=0.824
Cycle frequency, distance per cyle (d c) and internal work rate (Ẇ int)
The cycle frequency (cycles·min−1) as a function of average forward speed (m·s−1) is shown in Fig. 6 for all the investigated forms of aquatic locomotion. In this figure the continuous lines irradiating from the origin correspond to the average distances covered per cycle (d c, m·cycle−1). This figure shows that, in the range of aerobic speeds, the increase in speed is essentially obtained by an increase in the cycle frequency while the distance per cycle is maintained essentially constant (with the exception of kayaking). Hence, at least in the range of speeds considered here, each form of locomotion in water is almost univocally defined by the distance covered per cycle (d c).

The cycle frequency (f, cycles·min−1) is plotted as a function of the average speed (v, m·s−1) for the different forms of aquatic locomotion investigated in this study. The lines interpolate the original data over the aerobic speed range. The fitted equations are: L (swimming by using the leg kick at the surface): f=112.75·v 0.74; UWF (underwater swimming with SCUBA diving equipment): f=57.24·v 0.92;AL (swimming the front crawl): f=22.97·v 1.06; K (kayaking): f=8.19·v 1.86;R (rowing): f=4.04·v 1.44. The average distance that the body/shell/hull travels per cycle (d c , m·cycle−1) is indicated by the continuous lines irradiating from the origin. In the investigated speed range the following data apply: L=0.5 m·cycle−1; UWF=1.0 m·cycle−1; AL=2.5 m·cycle−1; K=4 m·cycle−1; R=8 m·cycle−1
As shown in Fig. 7, d c is related to the efficiency of locomotion: the higher d c the higher the propelling and overall efficiencies: η P=0.46, d c 0.21 (r=0.78) and η o=0.14, d c 0.34 (r=0.87).

Values of Froude (η F ) and propelling efficiency (η P ) are plotted as a function of the distance per cycle (d c , m) for the forms of locomotion investigated here
The internal work in aquatic locomotion was measured only for the leg kick and the arm stroke (Zamparo et al. 2002; Zamparo 2003). Ẇ int is a substantial fraction of Ẇ tot when swimming by kicking the legs (from 0.25 to 0.65 W·kg−1 at speeds between 0.6 and 1.0 m·s−1) because of the high limb frequencies necessary to produce forward motion (from 1.3 to 1.9 Hz in the same speed range). The limb frequency is greatly reduced when leg kicking with fins (from 0.7 to 1.3 Hz in the same speed range) and when swimming by using the arm stroke (again, from 0.3 to 0.6 Hz at speeds between 1 and 1.4 m·s−1) thus leading to a substantial reduction of Ẇ int: in the arm stroke and in the leg kick with fins Ẇ int ranges from 0.05 to 0.25 W·kg−1 whereas in the leg kick without fins it is twice that value, ranging from 0.25 to 0.65 W·kg−1 (see above). As shown in Fig. 6 the "average" cycle frequency is about half that of the leg kick without fins in all (the other) forms of water locomotion. Moreover, as indicated below, the external mechanical power of locomotion increases steadily as a function of the speed of movement in water; as such the contribution of Ẇ int to Ẇ tot, both in absolute and relative terms, is bound to be low and, as indicated in the Introduction, Ẇ int can be neglected and Ẇ tot can be assumed to be equal to the sum of Ẇ k and Ẇ d only.
Total mechanical power (Ẇ tot)
The total mechanical power for aquatic locomotion could be calculated rearranging Eq. 6 (Ẇ tot=Ė·η o); the values of Ẇ tot for all the forms of locomotion investigated here are reported in Table 2 along with the values of η o, η d, η P, C and Ẇ d as calculated for a metabolically equivalent speed of 0.5 kW. These data show that, as previously found for human locomotion on land, the use of passive tools for human locomotion in water (e.g. fins or hulls) leads to a decrease of the energy cost of locomotion at a given speed. At variance with locomotion on land, however, these data show that the decrease in the energy cost of locomotion is associated with an increase, rather than a decrease, in the total mechanical power. This is made possible by a continuous increase of the overall efficiency of locomotion from the slow speeds (and loads) associated with human swimming without aids to the high speeds (and loads) attainable with hulls and boats.
Discussion
In this paper we attempted to draw a complete energy balance of several forms of water locomotion. On the basis (1) of measured values of drag, energy cost and cycle frequency (experimentally determined for each form of locomotion) and (2) of measured or estimated values of propelling efficiency, the drag efficiency and the overall efficiency were computed.
Data reported in this paper show that human locomotion in water can be described by similar rules in spite of the differences in the "type of propeller" (legs or arms), of the different tools that can be utilized for locomotion (fins, kayaks and rowing shells), of the different principles on which the propulsive action is based (e.g. kayaks are propelled by lift on a hydrofoil and rowing shells by drag on the oars, see Alexander 2003) and of the different pattern of movement adopted (the limbs movement is synchronous in rowing and alternate in all other forms of locomotion).
In this paper we aimed to find the trends, similarities and analogies among different strategies of motion in the aquatic environment. The data reported in this study are, therefore, not meant to define unequivocally a form of locomotion but rather to be an example of the relationship between the energetics and mechanics of different strategies of moving in water.
As an example it was shown by Zamparo et al. (1999) that, at speeds between 1 and 2 m·s−1, kayaking with a flat-water sprint K1 scull is more economical (C=0.020 v 2.26) than kayaking with a slalom canoe (C=0.098 v 1.76, see Figs. 1, 2). As a consequence, the difference in C between competitive rowing shells (as reported in this study: C=0.088 v 1.05) and competitive K1 kayaks is less than reported in Fig. 2. It seems also interesting to note here that the energy cost per unit distance of kayaking with a slalom canoe, at the lowest investigated speed (1 m·s−1), is not far from that of sculling a Venetian Gondola (as reported by Capelli et al. 1990) for which C=0.155 v 1.67 (in all these examples C is in kJ·m−1 and v in m·s−1).
The difference in the energy expenditure between the two types of kayaks mentioned above was attributed to drag differences between the sculls. Indeed the work expended to overcome drag is the major determinant of the metabolic expenditure in rowing and kayaking for which: Ẇ d=Ė·0.21 (r 2=0.981, n=30). The slope of this relationship is the drag (performance) efficiency of "boat locomotion"; as shown in Fig. 4, this efficiency is much lower (about 0.07) in all other conditions (L, UWF, AL).
The differences in performance efficiency between aquatic sports in which boats/sculls and paddles/oars are utilized and those in which the human body is moving "alone" in water, propelled only by the upper/lower limbs, can be attributed to several reasons.
The kayak and the rowing shell float on the surface and have a low pressure drag: in these forms of locomotion the wetted surface area is the major determinant of drag. This is not so for the human body where also other factors influence water resistance. Particularly at low speeds (less than 1 m·s−1, e.g. when swimming by kicking the legs only) the swimmers assume an inclined position in the water and their frontal surface area is the major determinant of drag. During forward progression in water the angle of the swimmer with the horizontal, and thus their frontal area, is the result of a balance of torque, or rotational forces around the center of mass, and hydrodynamic lift (Pendergast et al. 1977; Zamparo et al. 1996, 2000; Capelli et al. 1995). As the velocity of progression increases, the hydrodynamic lift increases, the body becomes more horizontal and the frontal area decreases. At these speeds, however, the level of immersion of the body in water becomes important: the larger this underwater weight, the greater the energy cost of swimming. This has also been shown in rowing where drag increased by about 10% for a 20% increase in weight (Secher 1993) and in kayaking where C increased by about 20% for a 20% increase in weight (Pendergast et al. 1989).
Despite the different factors determining drag, each form of water locomotion can be characterized by its specific relationships between the speed v and Ė, D and η d. These relationships can be described by power functions, as reported in the legends of Figs. 1, 3 and 4 (nb Ė is in kW, D in N and η d is dimensionless). They are interdependent, so only two out of three are necessary to fully describe the characteristics of the locomotion at stake:
and since Ė=Ẇ d/η d
On the basis of these equations it is possible not only to characterize different forms of locomotion in water but also to calculate an unknown parameter (e.g. the active drag) from the other two (e.g. Ė and η d). As an example, for kayaking with a flat-water sprint K1 scull and for sculling a Venetian Gondola only the relationship between Ė and v (Eq. c) is known from experimental data (Zamparo et al. 1999; Capelli et al. 1990). It is fair to assume that the relationship between η d and v computed for kayaking and rowing in this study (Eq. b, where k 2=0.16 and m=0.2) holds also for these two (similar) forms of locomotion. Hence, by combining Eqs. b and c it is possible to compute the D vs. v relationship (Eq. a) for the flat water K1 skull (D=3.2·v 2.46) and for the Venetian Gondola (D=24.8·v 1.87). These data show that, as expected, the boat opposing the greater resistance in water among these four is the Venetian Gondola and the active drag of a competitive K1 skull is very close to that of a rowing shell.
As pointed out in this paper and by several authors in recent decades (e.g. Toussaint et al. 1988) the drag efficiency is only a "partial efficiency" of water locomotion since it takes into account only the effects of water resistance on the energy expenditure. However, as underlined in studies of fish locomotion (Daniel 1991), this parameter is rather important in practical terms since it is a measure of the efficiency with which the energy is transformed into useful work (the thrust) for progression in water.
The overall efficiency (total work performed/total energy expended) of locomotion can be easily computed once the propelling efficiency (e.g. the efficiency with which the total work production is transformed in useful thrust) of each form of locomotion is known (η ο=η d/ η P).
As pointed out at the beginning of the Discussion, the values of η P utilized in this study are not meant to define unequivocally a form of locomotion but rather to be indicative of the ratio of power "wasted" to total power production in this broad range of aquatic activities. Whereas several authors report a value of η P of about 0.7 for boat locomotion and the propelling efficiency for leg kicking with and without fins was experimentally determined, there is no general agreement on the value of η P for the front crawl (see also Table 1). Hence the calculated value of η ο for this form of locomotion in water is less certain than in the other cases. As shown by Fig. 5, disregarding the data points referring to the front crawl, the η o vs. speed relationship could be described by two separate functions: η o=0.14·v 0.80, n=15, r 2=0.824 for the leg kick (L and UWF) and η ο=0.21·v 0.28, n=30, r 2=0.994 for boat locomotion (R and K) with the calculated data of η ο for the front crawl lying between the two.
In turn, the reported values of η o for rowing and kayaking are close to the values estimated by using rowing ergometers or by indirect calculations of mechanical power during actual competitions for these forms of locomotion (about 0.2–0.3, for a review see Hagerman 2000).
From Fig. 5 it is also possible to see that, as a general rule, the overall efficiency of aquatic locomotion increases from the slow speeds (and loads) associated with human swimming without aids to the high speeds (and loads) attainable with hulls and boats. In the latter case the overall efficiency attains values (0.25–0.35) similar to those observed for land locomotion at optimal contraction speeds (Woledge et al. 1985; Sargeant and Jones 1995; Minetti et al. 2001).
This message is also apparent by inspection of Table 2 which shows that, at variance with locomotion on land, the decrease in the energy requirements of water locomotion is associated with an increase, rather than a decrease, in the mechanical power output which is offset by an even larger increase in efficiency.
These data suggest that propulsion in water is limited by the amount of force that can be applied to the water. The reason of the "sub-optimal" values of overall efficiency in swimming (below 0.22 in "non" boat locomotion) could therefore be attributed to the fact that, in water, the muscles are forced to work far from the optimum in their force/length–force/speed relationship.
A further interesting outcome of the "global" analysis of aquatic locomotion proposed in this study can be derived by inspection of Figs. 6 and 7. At least in the range of speeds considered here (aerobic), the increase in speed is obtained by an increase in the frequency of the movement of the "propeller" and by maintaining the same distance per stroke (with the exception of kayaking). Hence, each form of locomotion in water can be almost univocally described by the distance the body/hull covers per cycle (d c, m·cycle−1) which, in turn, is related to the efficiency of locomotion: the higher d c the higher the propelling and overall efficiencies.
This finding is quite interesting since the link between d c and efficiency has been suggested and discussed previously by several authors. Particularly in the case of the front crawl, the distance the subject travels per stroke was proposed as an index of the propelling efficiency of the swimmer (e.g. Craig and Pendergast 1979): the higher the distance per stroke for any given speed the more efficient the propulsion in water (see, for a brief review, also Toussaint and Beek 1992). As shown in the literature (e.g. swimming: Termin and Pendergast 2001) subjects could indeed be trained to increase the distance per stroke (at a given set of speeds). It was also shown that the obtained increase in d c was associated with a decrease in the energy expenditure (when swimming/paddling) at the same speed or over the same distance. This indeed is the case when the efficiency of propulsion in water is improved.
References
Abbott AV, Brooks AN, Wilson DG (1995) Human-powered watercraft. In: Abbott AV, Wilson DG (eds) Human-powered vehicles. Human Kinetics, Champaigin, Ill., pp 49–67
Alexander R McN (1983) Motion in fluids. In: Animal mechanics. Blackwell Scientific, Oxford, pp 183–233
Alexander R McN (2003) Principles of animal locomotion. Princeton University Press, Princeton, N.J., pp 249–287
Berger MA, Hollander AP, De Groot G (1997) Technique and energy losses in front crawl swimming. Med Sci Sports Exerc 29:1491–1498
Capelli C, Donatelli C, Moia C, Valier C, Rosa G, di Prampero PE (1990) Energy cost and efficiency of sculling a venetian Gondola. Eur J Appl Physiol 60:175–178
Capelli C, Zamparo P, Gigalotto A, Francescato MP, Soule RG, Termin B, Pendergast DR, di Prampero PE (1995) Bioenergetics and biomechanics of front crawl swimming. J Appl Physiol 78(2):674–679
Capelli C, Pendergast DR, Termin B (1998) Energetics of swimming at maximal speeds in humans. Eur J Appl Physiol 78:385–393
Celentano F, Cortili G, di Prampero PE, Cerretelli P (1974) Mechanical aspects of rowing. J Appl Physiol. 36:642–647
Craig AB Jr, Pendergast DR (1979) Relationships of stroke rate, distance per stroke, and velocity in competitive swimming. Med Sci Sports Exerc 11(3):278–283
Dal Monte A, Komor A (1989) Rowing and sculling mechanics. In: Vaughan CL (ed) Biomechanics of sport. CRC Press, Boca Raton, Fla., pp 53–120
Daniel T (1991) Efficiency in aquatic locomotion: limitations from single cells to animals. In: Blake RW (eds) Efficiency and economy in animal physiology. Cambridge University Press, Cambridge, pp 83–95
di Prampero PE, Cortili G, Celentano F, Cerretelli P (1971) Physiological aspect of rowing. J Appl Physiol 31:853–857
di Prampero PE, Pendergast DR, Wilson DR, Rennie DW (1974) Energetics of swimming in man. J Appl Physiol 37(1):1–5
Fenn WO (1930) Frictional and kinetic factors in the work of sprint running. Am J Physiol 92:583–611
Hagerman FC (2000) Physiology of competitive rowing. In: Garrett WE, Kirkendall DT (eds) Exercise and sport science. Lippincott Williams and Wilkins, Philadelphia, pp 843–874
Lighthill MJ (1975) Hydrodynamics of aquatic animal propulsion. In: Mathematical biofluidodynamics. Society for Industrial and Applied Mathematics, Philadelphia
Martin RB, Yeater RA, White MK (1980) A simple analytical model for the crawl stroke. J Biomech 14:539–548
Minetti AE (1998) A model equation for the prediction of mechanical internal work of terrestrial locomotion. J Biomech 31:463–468
Minetti AE, Pinkerton J, Zamparo P (2001) Form bipedalism to bicyclism: evolution in energetics and biomechanics of historical bicycles. Proc R Soc Lond B 268:1351–1360
Pendergast DR, di Prampero PE, Craig AB Jr, Wilson DR, Rennie DW (1977) Quantitative analysis of the front crawl in men and women. J Appl Physiol: 43(3):475–479
Pendergast DR, Bushnell D, Wilson DR, Cerretelli P (1989) Energetics of kayaking. Eur J Appl Physiol 59:342–350
Pendergast DR, Tedesco M, Nawrocki DM, Fisher NM (1996) Energetics of underwater swimming with SCUBA. Med Sci Sports Exerc 28:573–580
Samimy S (2002) A theoretical and experimental analysis of diver and fin performance in underwater swimming. Masters Thesis, Department of Mechanical Engineering, University of New York at Buffalo, Buffalo, N.Y.
Sanders RH, Cappaert J, Devlin RK (1995) Wave characteristics of butterfly swimming. J Biomech 28:9–16
Sargeant AJ, Jones DA (1995) The significance of motor unit variability in sustaining mechanical output of muscle. In: Gandevia S et al. (eds) Neural and neuromuscular aspects of muscle fatigue. Plenum, New York, pp 323–338
Secher NH (1993) Physiological and biomechanical aspects of rowing. Sports Med 15:24–42
Termin B, Pendergast DR (2001) Training using the stroke frequency-velocity relationship to combine biomechanical and metabolic paradigms. J Swim Res 14:9–17
Toussaint HM, Beek PJ (1992) Biomechanics of competitive front crawl swimming. Sports Med 13:8–24
Toussaint HM, Beelen A, Rodenburg A, Sargent AJ, de Groot G, Hollander AP, van Ingen Schenau GJ (1988) Propelling efficiency of front crawl swimming. J Appl Physiol 65:2506–2512
Toussaint HM, Janssen T, Kluft M (1991) Effect of propelling surface size on the mechanics and energetics of front crawl swimming. J Biomech 24:205–211
Woledge RC, Curtin NA, Homsher E (1985) Energetic aspects of muscle contraction. Academic, London
Zamparo P (2003) Optimization and transmission efficiency in human locomotion, Doctoral Thesis, Department of Exercise and Sport Sciences, Manchester Metropolitan University, UK
Zamparo P, Antonutto G, Capelli C, Francescato MP, Girardis M, Sangoi R, Soule RG, Pendergast DR (1996) Effects of body size, body density, gender and growth on underwater torque. Scand J Med Sci Sports 6:273–280
Zamparo P, Capelli C, Guerrini G (1999) Energetics of kayaking at submaximal and maximal speeds. EurJ Appl Physiol 80:542–548
Zamparo P, Capelli C, Cautero M, di Nino A (2000) Energy cost of front-crawl swimming at supra-maximal speeds and underwater torque in young swimmers. Eur J Appl Physiol 83:487–491
Zamparo P, Pendergast DR, Termin B, Minetti AE (2002) How fins affect the economy and efficiency of human swimming. J Exp Biol 205:2665–2676
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Pendergast, D., Zamparo, P., di Prampero, P.E. et al. Energy balance of human locomotion in water. Eur J Appl Physiol 90, 377–386 (2003). https://doi.org/10.1007/s00421-003-0919-y
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-003-0919-y