
Abstract
Haplotype and allele frequencies of 12 X-STRs included in the Investigator Argus X-12 kit are reported for 255 individuals, representing four Western Mediterranean populations: Valencia (eastern mainland Spain) and the Balearic Islands (Majorca, Minorca, and Ibiza). Ibiza shows the lowest intra-population variability and the highest level of linkage disequilibrium together with an important genetic distance with regard to the geographically close populations, which is consistent with the historical evidence for long-term demographic isolation and its different matrilineal background.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
The Balearic archipelago (Majorca, Minorca, and Ibiza islands, in the Western Mediterranean Sea) has been inhabited for 5,500 years, and different people—especially the Romans (third century BC) and Catalans (early thirteenth century)—contributed to the genetic pool of the current population, but no remarkable additional contribution of foreign genes was received until recently because of immigration. Genetic studies [1, 2] show differences between the three islands and a remarkable differentiation of Ibiza compared to other Western Mediterranean populations, probably because of the Phoenician–Carthaginian origin of the first settlers and the effect of genetic drift. Few data regarding X-chromosome short tandem repeats (STRs) have been reported in Western Mediterranean populations [3, 4] and there are no available data in the Balearic Islands. It is essential to give more insight into the genetic substructure in this area, focusing on known isolates such as the Ibiza population. In this work, we studied the genetic diversity, geographic distribution, and population structure of three insular populations from the Balearic Archipelago and Valencia (eastern coast of the Iberian Peninsula), as a mainland Spanish reference.
Mouthwash samples were obtained from 255 unrelated individuals (160 males and 95 females), after informed consent. DNA was extracted by standard phenol-chloroform method. PCR amplification, capillary electrophoresis, data analysis and calculation of allele frequencies, and additional relevant population and forensic statistical parameters were performed as previously described [4]. To examine the relationship of these populations with neighboring populations, F ST genetic distances, calculated using POPTREE2.69 [5], were performed to generate the multi-dimensional scaling (MDS) plot conducted with SPSS v.15.0 (SPSS, Inc., Chicago, IL, USA).
Allele frequencies are shown in Supplementary Table 1. The highest variability was found in DXS10135, as described elsewhere [6], with 27 alleles and observed heterozygosities ranging between 0.864 and 0.950. The lowest heterozygosity (0.480) was observed for the DXS7423 system in Minorca. No deviations from HWE were observed after Bonferroni correction. Typing of the 160 males from the Western Mediterranean region resulted in 160 different haplotypes when all 12 X-STRs were included. Linkage groups 1–4 revealed 135, 98, 93, and 103 haplotypes, respectively (Table 1). Among all the observed haplotypes, 96.4 % showed frequencies <0.020 and the three most common haplotypes were observed in eight individuals each, displaying a frequency of 0.050. In these populations, linkage group 1 proved to be the most polymorphic group and linkage group 3 the least, in accordance with other studies [6]. The Valencia population showed greater haplotype diversity than Balearic Island populations, with a higher percentage of unique haplotypes in the four linkage groups. Supplementary Table 2 shows haplotype frequencies for each population. The Ibiza population showed the lowest haplotype diversity (HD) values—with a low percentage of unique haplotypes (between 46.3 and 56.1 %) compared to the other populations studied (64.1–94.9 %)—and the highest level of linkage disequilibrium (LD) in most of the loci pairs inside each linkage group, although after Bonferroni correction only one significant p value was observed for DXS10148-DXS10135 pair of loci (p = 0.000) (Supplementary Table 3). LD does not depend exclusively on the physical distance between loci, but may result from random genetic drift, founder effect, mutations, selection, and population admixture or stratification [7]; therefore, the greater LD found in Ibiza could result from the founder effect and genetic drift related to the demographic and historical features of this population (an isolated, consanguineous population with a reduced effective population size), supporting the previous genetic studies [1]. When Balearic Island populations were pooled, significant LD was observed inside each linkage group. Although no significant differences in allelic frequency were observed in the Balearic populations for the 12 X-STRs studied, different studies emphasize the importance of investigating these populations and attempt to describe their roots and genetic substructure [2, 8]; therefore, these LD values could result from the heterogeneous distribution of the haplotypes among the islands. Forensic parameters of interest were calculated for each X-STR and population (Supplementary Table 1). The combined power of discrimination (PD) in females ranged from 1 in 4.74 E + 14 (in Ibiza) to 1 in 1.80 E + 15 (in Mallorca and Valencia). Combined PDmales values ranged from 1 in 5.33 E + 08 (in Ibiza) to 1 in 1.04 E + 09 (in Valencia). A high combined MEC was also obtained for father–daughter duos and father–girl–mother trios (>0.99999 in all cases). Although values differed slightly between populations, the set of loci in the Argus X-12 kit was highly informative in all the Western Mediterranean populations studied. A comparison between the Western Mediterranean population and 12 other European and African (mainly North African) populations with available data for the same set of X-STRS markers was performed. Supplementary Fig. 1 shows a multi-dimensional scaling plot based on pair wise F ST genetic distances. Along the X-axis, the distribution of the populations ranging from Northeast Africa to Northern Europe—with the Mediterranean populations showing an intermediate position—can be observed, as found in X-chromosome SNPs studies [9]. However, Mallorca, Menorca, and Valencia are closer to other European populations than to North Africans, in accordance with other studies that suggest the existence of a relative north–south gene flow barrier in the Western part of the Mediterranean area [2, 10]. Ibiza and Sahrawi populations show a remote position versus their geographic neighboring populations, which may be due to historical and cultural background in the Sahrawi case. Regarding Ibiza, its differentiation versus other European populations was also evidenced by mtDNA analysis, but not by Y-chromosome markers, indicating a sex-biased contribution to the genetic pool of Ibiza [1]. Indeed, as two thirds of the X-chromosomes descend from maternal origin, X-chromosome polymorphisms will mostly behave as matrilineal markers showing similar results to those obtained from mtDNA data. Therefore, the observed displaced plot of Ibiza based on X-STR results may reflect the differential matrilineal background of this population, probably accentuated by the effect of genetic drift. In conclusion, the results of the present study provide a useful X-STR database for the Western Mediterranean region. It is important to highlight that a local haplotype database must be used especially in Ibiza due to the matrilineal genetic features of this population.
References
Tomàs C, Jiménez G, Picornell A, Castro JA, Ramon MM (2006) Differential maternal and paternal contributions to the genetic pool of Ibiza Island, Balearic Archipelago. Am J Phys Anthropol 129:268–278
Rodríguez V, Tomàs C, Sánchez JJ, Castro JA, Ramon MM, Barbaro A, Morling N, Picornell A (2009) Genetic sub-structure in western Mediterranean populations revealed by 12 Y-chromosome STR loci. Int J Legal Med 123:137–141
Barbaro A, Cormaci P, Falcone G, Votano S, La Marca A (2012) Distribution of 8 X-chromosomal STR loci in an Italian population sample (Calabria). Forensic Sci Int Genet 6:174–175
Bentayebi K, Picornell A, Bouabdeallah M, Castro JA, Aboukhalid R, Squalli D, Misericòrdia M, Amzazi S (2012) Genetic diversity of 12 X-chromosomal short tandem repeats in the Moroccan population. Forensic Sci Int Genet 6:48–49
Takezaki N, Nei M, Tamura K (2010) POPTREE2: software for constructing population trees from allele frequency data and computing other population statistics with windows-interface. Mol Biol Evol 27:747–752
Tomàs C, Pereira V, Morling N (2012) Analysis of 12 X-STRs in Greenlanders, Danes and Somalis using Argus X-12. Int J Legal Med 126:121–128
Chakravarti A (1999) Population genetics-making sense out of sequence. Nat Genet 21:56–60
López-Escribano H, Parera MM, Guix P, Serra JM, Gutierrez A, Balsells D, Oliva-Berini E, Castro JA, Ramon MM, Picornell A (2013) Balearic archipelago: three islands, three beta-thalassemia population patterns. Clin Genet 83:175–180
Tomás C, Sanchez JJ, Barbaro A, Brandt-Casadevall C, Hernandez A, Ben Dhiab M, Ramon M, Morling N (2008) X-chromosome SNP analyses in 11 human Mediterranean populations show a high overall genetic homogeneity except in North-west Africans (Moroccans). BMC Evol Biol 8:75
Comas D, Calafell F, Benchemsi N, Helal A, Lefranc G, Stoneking M, Batzer MA, Bertranpetit J, Sajantilla A (2000) Alu insertion polymorphisms in NW Africa and the Iberian peninsula: Evidence for a strong genetic boundary through the Gibraltar straits. Hum Genet 107:312–329
Acknowledgments
This work was partially supported by grant AAEE133/09 from the Direcció General de R + D + I (Comunitat autònoma de les Illes Balears) and European Regional Development Fund (ERDF).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
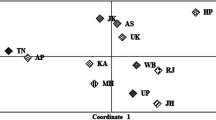
Supplementary Figure 1
Multidimensional scaling analysis (MDS) based on Reynolds genetic distances calculated between populations. African populations are represented by circular points, European populations by square dots and Balearic and Valencia populations are labelled with stars. (PDF 212 kb)
Supplementary Table 1
Allele frequencies of 12X-STR in 350 chromosomes (Majorca (MAJ): 41 men and 22 women; Minorca (MIN): 39 men and 25 women; Ibiza (IBI): 41 males and 20 women; Valencia (VAL): 39 men and 28 women) (PDF 47 kb)
Supplementary Table 2
X-chromosome haplotypes of four linked STR trios in 121 males from Balearic Islands (41 from Majorca, 39 from Minorca and 41 from Ibiza) and 39 males from Valencia populations. More common haplotypes (5%) are labelled with * (PDF 337 kb)
Supplementary Table 3
Linkage disequilibrium inside the 4 Linkage Groups (LG) in Mallorca, Minorca, Ibiza and Valencia populations. In bold, significant p-values after Bonferroni correction (PDF 135 kb)
Rights and permissions
About this article

Cite this article
Ferragut, J.F., Bentayebi, K., Castro, J.A. et al. Genetic analysis of 12 X-chromosome STRs in Western Mediterranean populations. Int J Legal Med 129, 253–255 (2015). https://doi.org/10.1007/s00414-014-1071-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00414-014-1071-4