
Abstract
Populations of the gecko lizard Gekko hokouensis (Gekkonidae, Squamata) on Okinawajima Island and a few other islands of the Ryukyu Archipelago, Japan, have the morphologically differentiated sex chromosomes, the acrocentric Z chromosome and the subtelocentric W chromosome, although the continental representative of this species reportedly shows no sex chromosome heteromorphism. To investigate the origin of sex chromosomes and the process of sex chromosomal differentiation in this species, we molecularly cloned the homologues of six chicken Z-linked genes and mapped them to the metaphase chromosomes of the Okinawajima sample. They were all localized to the Z and W chromosomes in the order ACO1/IREBP–RPS6–DMRT1–CHD1–GHR–ATP5A1, indicating that the origin of ZW chromosomes in G. hokouensis is the same as that in the class Aves, but is different from that in the suborder Ophidia. These results suggest that in reptiles the origin of sex chromosomes varies even within such a small clade as the order Squamata, employing a variety of genetic sex determination. ACO1/IREBP, RPS6, and DMRT1 were located on the Z long arm and the W short arm in the same order, suggesting that multiple rearrangements have occurred in this region of the W chromosome, where genetic differentiation between the Z and W chromosomes has been probably caused by the cessation of meiotic recombination.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Reptiles exhibit an extraordinary variability in sex chromosomal constitution and sex-determining system (Ciofi and Swingland 1997; Sarre et al. 2004; Valenzuela and Lance 2004). All crocodilians, the tuatara, most turtles, and some lizards have temperature-dependent sex determination system (Ferguson and Joanen 1982; Head et al. 1987; Lang and Andrews 1994; Cree et al. 1995; Ciofi and Swingland 1997; Valenzuela and Lance 2004), in which no karyotypic difference was found between males and females, whereas all snakes exhibit genetic sex determination (GSD) system with female heterogamety as birds (Beçak et al. 1964; Beçak and Beçak 1969; Singh 1972). Other reptilian species showing GSD (some lizards and a few turtles) have both male and female heterogameties (Olmo and Signorino 2005). The origin of sex chromosomes and the process of sex chromosomal differentiation have been still almost unknown in reptiles because the number of species whose sex chromosomes were identified is quite small except for snakes. The sex microchromosomes were recently identified in several reptilian species using molecular cytogenetic techniques (Ezaz et al. 2005, 2006b; Kawai et al. 2007), suggesting that the sex microchromosomes, which have been not identified by gross analyses of standard karyotypes, will be found for more GSD species of reptiles in the future.
The recent physical and genetic linkage maps of the chicken revealed the extensive homology between chicken and human chromosomes (Burt 2002; Groenen et al. 2000; International Chicken Genome Sequencing Consortium 2004; Schmid et al. 2005); however, there was no homology between the human XY and chicken ZW sex chromosomes, suggesting that the sex chromosomes of mammals and birds were derived from different autosomal pairs of the common ancestor (Nanda et al. 1999, 2000, 2002). Snakes share a number of karyotypic characteristics with birds, such as the physical size composition of chromosomes (distinct differentiation into macrochromosomes and microchromosomes) and the highly differentiated ZW-type sex chromosomes with the exception of the boid snakes that have less differentiated sex chromosomes (Beçak et al. 1964; Beçak and Beçak 1969; Singh 1972). The similarity of the karyotypes between snakes and birds leads us to predict that the snake ZW sex chromosomes have the same origin as the avian ZW sex chromosomes (Ohno 1967). However, the comparative cytogenetic mapping between chicken and the Japanese four-striped rat snake (Elaphe quadrivirgata: Colubridae) showed that the chicken Z-linked genes were localized to the short arm of the snake autosome 2, and, by contrast, the chicken homologues of the snake Z-linked gene were located on chicken autosomes (Matsuda et al. 2005; Matsubara et al. 2006; Kawai et al. 2007). The absence of homology with the chicken Z chromosome in the snake Z chromosome was also confirmed by another study using the habu (Protobothrops flavoviridis, formerly Trimeresurus flavoviridis: Viperidae) and the Burmese python (Python molurus bivittatus: Boidae) as snake representatives (Matsubara et al. 2006). Furthermore, the difference in the sex chromosomal origin from birds was also confirmedly demonstrated for the Chinese soft-shelled turtle (Pelodiscus sinensis), which has the ZW-type sex microchromosomes (Matsuda et al. 2005; Kawai et al. 2007). These results strongly suggest that the sex chromosomes of the Reptilia, Aves, and Mammalia were all derived from the different autosomal pairs of the common amniote ancestor, which first appeared around 310 million years ago (MYA; Kumar and Hedges 1998). They also indicate that the sex chromosomes have been independently differentiated at least in a few lineages of these taxa by involving a variety of genes for establishment of genetic sex determination systems.
Lizards (suborder Lacertila) are the largest group in reptiles and are composed of more than 4,500 species of 20 families. The Gekkonidae comprises approximately 1,000 species, and the karyotypes have been reported for 145 species (Ota et al. 2001; Olmo and Signorino 2005). Out of these species, 13 have been shown to possess heteromorphic sex chromosomes including both female heterogamety (ZW) and male heterogamety (XY and X1X2Y). Cytogenetic studies on the sex chromosomes less progress for lizards than for snakes, and the origin of the lizard sex chromosomes and their chromosomal homology with the chicken Z chromosome, the mammalian X chromosomes, and the sex chromosomes of other reptilian taxa have never been studied.
The gecko lizard Gekko hokouensis of the Gekkonidae is widely distributed in southeastern continental China, Taiwan, and some islands of the Ryukyu Archipelago and a part of Kyushu, Japan (Ota 1989a; Toda et al. 2001, 2008). The karyotype of G. hokouensis (2n = 38) was first reported for the continental population, in which no morphologically heteromorphic sex chromosomes were identified (Chen et al. 1986). No differences in the diploid chromosome numbers have been found among this and other populations of this species; however, a prominent female-specific chromosomal heteromorphism was found in populations of the Ryukyu Archipelago as represented by samples from Okinawajima Island, Suwanosejima Island, and Nakanoshima Island (Kohno et al. unpublished data). In the Ryukyu populations, therefore, the presence of a genetic sex-determining system with female heterogamety (i.e., ZZ/ZW system) is highly likely.
Here, we molecularly cloned homologues of six chicken Z-linked genes in G. hokouensis with the putative ZZ/ZW system and mapped them to G. hokouensis chromosomes to examine the chromosomal homology of the G. hokouensis Z sex chromosome with the chicken Z sex chromosome and other reptilian sex chromosomes. We report a contrasting case that the ZW sex chromosomes of the Ryukyu G. hokouensis have the same origin as the avian ZW chromosomes. We also show the structural difference between the Z and W sex chromosome of G. hokouensis and discuss the process of sex chromosomal differentiation in this species.
Materials and methods
Animals
Nine adult males and nine adult females of G. hokouensis were captured in Nakagusuku, the southern part of Okinawajima Island, the Ryukyu Archipelago, Japan. Of these, three males and three females were subjected to chromosome preparation through cell culture and chromosome mapping (see below). The remaining six males and six females were karyotyped by simple bone-marrow air-dry method following Ota (1989b).
Cell culture and chromosome preparation
After anesthetization, heart tissues were collected from each individual and used for cell culture. All experimental procedures with the animals conformed to the guidelines established by the Animal Care Committee, Hokkaido University. The tissues were minced, and the fibroblast cells were cultured in DMEM medium (Invitrogen-GIBCO) supplemented with 15% fetal bovine serum (Invitrogen-GIBCO), 100 μg/ml kanamycin, and 1% antibiotic–antimycotic (PSA) (Invitrogen-GIBCO). The cell cultures were incubated at 26.5°C in a humidified atmosphere of 5% CO2 in air. Primarily, cultured fibroblast cells were harvested using trypsin and then subcultured.
After colcemid treatment (120 ng/ml) for 45 min, the cells were harvested, suspended in 0.075 M KCl for 20 min at room temperature, and fixed with 3:1 methanol/acetic acid three times following a standard protocol. After centrifugation, the cell suspension was dropped on glass slides and air-dried. Replication-banded chromosome preparation was performed for chromosome mapping of functional genes using fluorescence in situ hybridization (FISH) as described previously (Matsuda and Chapman 1995) with slight modification. Briefly, BrdU (12 μg/ml; Sigma-Aldrich) was added to the cell cultures at log phase, and the cell culture was continued for 12 h including 45 min of colcemid treatment before harvest. After BrdU treatment during the late replication stage, the cells were collected, and chromosome slides were made as mentioned previously. After the chromosome slides were stained with Hoechst 33258 (1 μg/ml) for 5 min, replication bands were obtained by heating the slides at 65°C for 3 min and exposing them to UV light at 65°C for an additional 6 min. The slides were kept at −80°C until use.
Molecular cloning of G. hokouensis homologues of chicken Z-linked genes
We molecularly cloned G. hokouensis homologues of six chicken Z-linked genes: soluble aconitase 1/iron-responsive element binding protein (ACO1/IREBP), ATP synthase, H+ transporting, mitochondrial F1 complex, alpha subunit, isoform 1, cardiac muscle (ATP5A1), chromodomain helicase DNA binding protein 1 (CHD1), growth hormone receptor (GHR), doublesex and mab-3-related transcription factor 1 (DMRT1), and ribosomal protein S6 (RPS6). Nucleotide sequences of the primers used for cDNA cloning of the genes are listed in Table 1. Testes were homogenized and lysed with TRIzol Reagent (Invitrogen), and total RNAs were extracted following the manufacturer’s instruction. Reverse transcription-polymerase chain reaction (RT-PCR) was performed to amplify the homologues of the chicken Z-linked genes. For the cDNA synthesis, 3 μg of total RNA was mixed with 0.5 μg Oligo (dT)12–18 Primer (Invitrogen) and incubated for 10 min at 70°C. After cooling on ice, the solution was mixed with 1× first strand buffer, 0.1 M DTT, 10 mM deoxyribonucleotide triphosphate (dNTP), and 200 U of SuperScript II RNase H− Reverse Transcriptase (Invitrogen). Reactions were performed for 50 min at 42°C and terminated by incubating for 15 min at 70°C. The cDNA amplification was carried out in 20 μl of 1× ExTaq buffer containing 1.5 mM MgCl2, 0.2 mM dNTPs, 5.0 μM degenerate primers, and 0.25 U of TaKaRa Ex Taq (Takara Bio). The PCR conditions were as follows: an initial denaturation at 94°C for 2 min, followed by 35 cycles at 94°C for 30 s, 58°C for 30 s, and 72°C for 35 s; and a final extension at 72°C for 5 min. The PCR products were electrophoresed on a 1% agarose gel and stained with ethidium bromide. The target DNA bands were isolated from the gel and extracted using a QIAquick Gel Extraction Kit (Qiagen). The DNA fragments were cloned using pGEM-T Easy Vector System I (Promega) and were transformed into Escherichia coli JM109 competent cells (Takara Bio). Nucleotide sequences were determined using an ABI PRISM3100 DNA Analyzer (Applied Biosystems) after sequencing reactions with a Big Dye Terminator v1.1 Cycle Sequencing Kit (Applied Biosystems).
Chromosome mapping
FISH was performed for chromosomal localization of G. hokouensis homologues of chicken Z-linked genes as described previously (Matsuda and Chapman 1995). For chromosome mapping of the ACO1/IREBP, ATP5A1, and CHD1 genes, two cDNA fragments isolated for each gene were used as probes to cover a wide region of the gene (Table 2). The cDNA fragments were labeled with biotin-16-dUTP (Roche Diagnostics) by nick translation following a standard protocol. After hybridization, the slides were incubated with goat anti-biotin antibody (Vector Laboratories) and then reacted with Alexa Fluor 488 rabbit anti-goat IgG (H+L) conjugate (Invitrogen-Molecular Probes) and stained with 0.75 μg/ml propidium iodide (PI). The hybridization signals were observed under a Nikon fluorescence microscope using Nikon filter sets B-2A and UV-2A, and the FISH images were microphotographed with DYNA HG ASA100 film (Kodak).
Results
Karyotype of G. hokouensis from Okinawajima Island
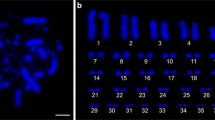
The diploid chromosome numbers of the nine males and nine females of G. hokouensis were invariably 38. The heteromorphism was observed in chromosome 4 for all nine females but not for any males: in the former one element of chromosome 4 was acrocentric and the other was subtelocentric (Fig. 1). Although there is no evidence that this heteromorphic chromosome pair has a sex-determining capacity, this sex-correlated chromosome heteromorphism is regarded as the ZZ/ZW-type sex chromosome system in the Okinawajima Island population (and other Ryukyu populations, see above). The autosomes were characterized by two large submetacentric (chromosomes 1 and 2), two small submetacentric (chromosomes 16 and 19), four large subtelocentric (chromosomes 6, 13, 14, and 15), one small subtelocentric (chromosome 17), one small metacentric (chromosome 18), and nine acrocentric pairs.

Giemsa-stained karyotype of female Gekko hokouensis from Okinawajima Island. The Z and W chromosomes (chromosome 4) are acrocentric and subtelocentric chromosomes, respectively
Molecular cloning of the chicken Z-linked gene homologues and their nucleotide sequences
The G. hokouensis homologues of the six chicken Z-linked genes were molecularly cloned (Table 2). The nucleotide sequences were determined for the cDNA fragments, and their consensus sequences were deposited in DNA Data Bank of Japan (DDBJ, http://www.ddbj.nig.ac.jp/Welcome.html). The nucleotide sequences of the six G. hokouensis homologues were compared in the equivalent regions with the cDNA fragments of three reptilian species, P. sinensis, E. quadrivirgata, and the Siamese crocodile (Crocodylus siamensis) (Table 3; Kawai et al. 2007). The highest identities were observed for ACO1/IREBP (84.6%), DMRT1 (83.1%), and GHR (75.9%) between G. hokouensis and E. quadrivirgata; for CHD1 (88.7%) and RPS6 (85.5%) between G. hokouensis and P. sinensis; and for ATP5A1 (83.8%) between G. hokouensis and C. siamensis.
Chromosomal locations of G. hokouensis homologues of chicken Z-linked genes
The G. hokouensis homologues of the six chicken Z-linked genes were all localized to both the Z and the W sex chromosomes (Fig. 2). More than 70 metaphase spreads were observed for each cDNA fragment. The hybridization efficiency was different between the cDNA fragments, ranging from around 30% to 70% of metaphase spreads. The order of the six genes was ACO1/IREBP–RPS6–DMRT1–CHD1–GHR–ATP5A1 from the centromere to the distal end on the long arm of the Z chromosome (Fig. 3). On the W chromosome, three genes, ACO1/IREBP, RPS6, and DMRT1, were located on the short arm in the same order from the distal to the centromere, and the other three genes were located at the same positions on the long arm as those of the Z chromosome.

Chromosomal localization of the six chicken Z-linked gene homologues to metaphase chromosome spreads of female G. hokouensis from Okinawajima Island. The cDNA fragments are used for chromosome mapping as biotin-labeled probes. a, b The GHR gene is localized to the Z and W chromosomes of PI-stained metaphase spread (a). The fluorescent Hoechst-stained pattern of the same metaphase spread as (a) is shown in (b). c, d The ATP5A1 gene is located on the Z and W chromosomes of PI-stained metaphase spreads. e–h FISH patterns of the CHD1 gene (e, g) and Hoechst-stained patterns (f, h) of the same metaphase spreads, respectively. The GHR, ATP5A1, and CHD1 genes are arranged in the same locations on the long arms of the Z and W chromosomes. i–t Chromosomal localization of the ACO1/IREBP (i–l), RPS6 (m–p), and DMRT1 (q–t) genes. The Hoechst-stained patterns of the same PI-stained metaphase spreads as (i, k, m, o, q, and s) are shown in (j, l, n, p, r, and t), respectively. The ACO1/IREBP, RPS6, and DMRT1 genes are localized to the long arm of the Z chromosome and the short arm of the W chromosome. Arrows indicate the hybridization signals. A scale bar represents 5 μm. All the pictures are shown in the same magnification

Comparative cytogenetic map of the six chicken Z-linked gene homologues on the Z and W chromosomes of G. hokouensis from Okinawajima Island
Discussion
There is a large diversity of sex-determining systems in lizards: temperature-dependent sex determination with no differentiation of sex chromosomes, and genetic sex determination with male and female heterogameties (Ciofi and Swingland 1997; Valenzuela and Lance 2004; Olmo and Signorino 2005). G. hokouensis from Okinawajima Island examined in this study exhibited the ZZ/ZW-type female heterogametic sex chromosomes. The homologues of the six chicken Z-linked genes, ACO1/IREBP, RPS6, DMRT1, CHD1, GHR, and ATP5A1, were all mapped to the Z and W sex chromosomes of G. hokouensis, indicating that the G. hokouensis ZW sex chromosomes have the same origin as the avian ZW sex chromosomes. This is the first case in reptiles, which represents the same sex-linkage as the chicken Z chromosome, and is in contrast to our previous data that the chicken Z sex chromosome corresponded to autosomes of three snake species and one turtle species (Matsuda et al. 2005; Matsubara et al. 2006; Kawai et al. 2007). These results collectively suggest that in reptiles the origin of the sex chromosomes varies even within such a small clade as the order Squamata, and a variety of sex-determining genes are employed for genetic sex determination in those taxa.
The acrocentric Z sex chromosome of paleognathous birds is the ancestral type of bird sex chromosomes (Ansari et al. 1988; Ogawa et al. 1998; Nishida-Umehara et al. 1999, 2007; Shetty et al. 1999; Tsuda et al. 2007), and the metacentric chicken Z chromosome probably resulted from multiple intrachromosomal rearrangements that independently occurred in the lineage of the Phasianidae of the Galliformes (Shibusawa et al. 2004). The order of the six chicken Z-linked genes, ACO1/IREBP–RPS6–DMRT1–CHD1–GHR–ATP5A1, has been found on the Z sex chromosomes of the ostrich (Struthio camelus) and the elegant crested tinamou (Eudromia elegans) of the Palaeognathae (Tsuda et al. 2007), whose lineage was estimated to have diverged around 120 million years ago (MYA) (van Tuinen and Hedges 2001). The same order of the genes has been conserved on the Z sex chromosome of G. hokouensis from Okinawajima Island, as well as on the chromosome 6 of P. sinensis and chromosome 2p of E. quadrivirgata, which correspond to the G. hokouensis Z sex chromosome (Fig. 4). These results suggest that the linkage of the avian Z-linked genes and their order that had been located on an autosomal pair of the common ancestor of sauropsids have been conserved in both the lineages of the Archosauromorpha and the Lepidosauria since they diverged 260–290 MYA (Kumar and Hedges 1998; Kumazawa and Nishida 1999; Janke et al. 2001; Iwabe et al. 2005; Benton and Donoghue 2007; Kumazawa 2007).

Comparative cytogenetic maps of the six chicken Z-linked gene homologues in the ostrich (Struthio camelus, SCA) and three reptilian species, the Chinese soft-shelled turtle (Pelodiscus sinensis, PSI), the Japanese four-striped rat snake (Elaphe quadrivirgata, EQU), and the gecko lizard (G. hokouensis, GHO). The ideograms of the Z chromosomes of the ostrich and two reptilian species and the chromosomal locations of the six genes were taken from our previous studies (Kawai et al. 2007; Tsuda et al. 2007). The E. quadrivirgata chromosome 2p is arranged upside down to make the gene order correspond to those on the S. camelus Z chromosome (SCA Z), the P. sinensis chromosome 6 (PSI 6), and the G. hokouensis chromosome Z (GHO Z)
Ohno (1967) asserted that the heteromorphic sex chromosomes are originated from a homologous autosomal pair through the occurrence of dominant sex determinator on one of the pair. It is generally accepted that a pericentric inversion is the most likely mechanism as an initial step of sex chromosomal differentiation. The ACO1/IREBP, RPS6, and DMRT1 genes were localized to the long arm of the Z chromosome and the short arm of the W chromosome in G. hokouensis, and the order of the three genes, ACO1/IREBP–RPS6–DMRT1, from the centromere to the distal on the Z long arm, was the same as that from the distal to the centromere on the W short arm. This suggests that several chromosome rearrangements, not a simple pericentric inversion alone, were involved in the process of the W chromosome differentiation in G. hokouensis: first, a pericentric inversion probably occurred in the region of a Z chromosome containing ACO1/IREBP, RPS6, and DMRT1, and then, positions of these three genes on the short arm of the resultant W chromosome were rearranged presumably through a series of paracentric inversions, becoming the same as those on the Z long arm in order. Further chromosome mapping data will allow us to precisely identify the chromosome rearrangements that actually occurred between the Z and W chromosomes in this species.
Meiotic recombination is suppressed in the differentiated region between the Z and W chromosomes, and the cessation of recombination is considered to favor the accumulation of gene mutations on the W chromosome. This leads us to predict that the W homologues of the Z-linked genes in the region have been diverged in nucleotide sequence as gametologous genes as found in the avian ZW sex chromosomes (Griffiths et al. 1996; de Kloet and de Kloet 2003; Handley et al. 2004; Tsuda et al. 2007). The DMRT1 gene, which encodes the DM domain that is a conserved component of the vertebrate sex-determining pathway, exhibits a gonad-specific and sexually dimorphic expression pattern (Raymond et al. 1998). This gene is located on the Z sex chromosome but not on the W chromosome in the chicken (Nanda et al. 2000). This is also true in the emu that has extensively homomorphic sex chromosomes (Shetty et al. 2002), suggesting that DMRT1 is a strong candidate for an avian sex-determining gene, which may represent a dosage-sensitive testis determinant (Nanda et al. 1999; Smith and Sinclair 2004). DMRT1 was localized to both the Z and W sex chromosomes in G. hokouensis; however, their locations were different between the Z and W chromosomes, indicating a possibility that the W homologue has been differentiated from the Z homologue. The accumulation of gene mutations should lead to the functional inactivation of the genes followed by the partial deletion of euchromatin and the accumulation of heterochromatin, which advance sex chromosome differentiation. This process of sex chromosome differentiation was clearly shown in the elegant crested tinamou, which is at more advanced stage of sex chromosome differentiation than the extensively homomorphic and euchromatic Z and W sex chromosomes of paleognathous ratites (Nishida-Umehara et al. 2007; Tsuda et al. 2007). The sex chromosome of G. hokouensis from Okinawajima Island, therefore, is considered to be at the early stage of sex chromosome differentiation. The presence of a conspecific population in the southeastern continental China with no sex chromosome heteromorphism at all (Chen et al. 1986, see above) circumstantially supports this view.
The genomic DNA clone of the DMRT1 gene was reported to be localized to the Z chromosome in the tiger snake (Notechis scutatus) (Ezaz et al. 2006a). By contrast, the cDNA clones of DMRT1 were commonly located on the short arm of chromosome 2 in other three snake species, P. flavoviridis of the Viperidae, E. quadrivirgata of the Colubridae, and P. molurus bivittatus of the Boidae (Matsubara et al. 2006). These results suggest a possibility of the multiple origins of snake sex chromosomes, but more detailed analysis is needed to clarify the origin of sex chromosomes and the diversity of sex-determining systems in squamates. The difference in the chromosomal location of this gene even within the Squamata implies the wide variety of the critical sex-determining genes in reptiles and that the mechanism of sex determination has evolved independently in each lineage. Further studies on the molecular structures of the sex chromosomes for more other reptilian species are required to clarify the process of sex chromosome differentiation and the evolution of genetic sex determination systems in reptiles.
References
Ansari HA, Takagi N, Sasaki M (1988) Morphological differentiation of sex chromosomes in three species of ratite birds. Cytogenet Cell Genet 47:185–188
Beçak W, Beçak ML (1969) Cytotaxonomy and chromosomal evolution in Serpentes. Cytogenetics 8:247–262
Beçak W, Beçak ML, Nazareth HRS, Ohno S (1964) Close karyological kinship between the reptilian suborder Serpentes and the class Aves. Chromosoma 15:606–617
Benton MJ, Donoghue PCJ (2007) Paleontological evidence to date the tree of life. Mol Biol Evol 24:26–53
Burt DW (2002) Origin and evolution of avian microchromosomes. Cytogenet Genome Res 96:97–112
Chen JC, Peng XB, Yu DW (1986) Studies on the karyotypes of three species of the genus Gekko. Acta Herpetologica Sinica 5:24–29
Ciofi C, Swingland IR (1997) Environmental sex determination in reptiles. Appl Anim Behav Sci 51:251–265
Cree A, Thompson MB, Daugherty CH (1995) Tuatara sex determination. Nature 375:543
de Kloet RS, de Kloet SR (2003) Evolution of the spindlin gene in birds: independent cessation of the recombination of sex chromosomes at the spindlin locus in neognathous birds and tinamous, a palaeognathous avian family. Genetica 119:333–342
Ezaz T, Quinn AE, Miura I, Sarre SD, Georges A, Graves JAM (2005) The dragon lizard Pogona vitticeps has ZZ/ZW micro-sex chromosomes. Chromosome Res 13:763–776
Ezaz T, Stiglec R, Veyrunes F, Grave JAM (2006a) Relationships between vertebrate ZW and XY sex chromosome systems. Curr Biol 16:R736–R743
Ezaz T, Valenzuela N, Grützner F, Miura I, Georges A, Burke RL, Graves JAM (2006b) An XX/XY sex microchromosome system in a freshwater turtle, Chelodina longicollis (Testudines: Chelidae) with genetic sex determination. Chromosome Res 14:139–150
Ferguson MWJ, Joanen T (1982) Temperature of egg incubation determines sex in Alligator mississippiensis. Nature 296:850–853
Griffiths R, Daan S, Dijkstra C (1996) Sex identification in birds using two CHD genes. Proc R Soc Lond B 263:1251–1256
Groenen MAM, Cheng HH, Bumstead N, Benkel BF, Briles WE, Burke T, Burt DW, Crittenden LB, Dodgson J, Hillel J, Lamont S, Ponce de Leon A, Soller M, Takahashi H, Vignal A (2000) A consensus linkage map of the chicken genome. Genome Res 10:137–147
Handley L-JL, Ceplitis H, Ellegren H (2004) Evolutionary strata on the chicken Z chromosome: implications for sex chromosome evolution. Genetics 167:367–376
Head G, May RM, Pendleton L (1987) Environmental determination of sex in the reptiles. Nature 329:198–199
International Chicken Genome Sequencing Consortium (2004) Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature 432:695–716
Iwabe N, Hara Y, Kumazawa Y, Shibamoto K, Saito Y, Miyata T, Katoh K (2005) Sister group relationship of turtles to the bird–crocodilian clade revealed by nuclear DNA-coded proteins. Mol Biol Evol 22:810–813
Janke A, Expenbeck D, Nilsson M, Arnason U (2001) The mitochondrial genomes of the iguana (Iguana iguana) and the caiman (Caiman crocodylus): implications for amniote phylogeny. Proc R Soc Lond B 268:623–631
Kawai A, Nishida-Umehara C, Ishijima J, Tsuda Y, Ota H, Matsuda Y (2007) Different origins of bird and reptile sex chromosomes inferred from comparative mapping of chicken Z-linked genes. Cytogenet Genome Res 117:92–102
Kumar S, Hedges SB (1998) A molecular timescale for vertebrate evolution. Nature 392:917–920
Kumazawa Y (2007) Mitochondrial genomes from major lizard families suggest their phylogenetic relationships and ancient radiations. Gene 388:19–26
Kumazawa Y, Nishida M (1999) Complete mitochondrial DNA sequences of the green turtle and blue-tailed mole skink: statistical evidence for Archosaurian affinity of turtles. Mol Biol Evol 16:784–792
Lang JW, Andrews HV (1994) Temperature-dependent sex determination in crocodilians. J Exp Zool 270:28–44
Matsubara K, Tarui H, Toriba M, Yamada K, Nishida-Umehara C, Agata K, Matsuda Y (2006) Evidence for different origin of sex chromosomes in snakes, birds, and mammals and step-wise differentiation of snake sex chromosomes. Proc Natl Acad Sci USA 103:18190–18195
Matsuda Y, Chapman VM (1995) Application of fluorescence in situ hybridization in genome analysis of the mouse. Electrophoresis 16:261–272
Matsuda Y, Nishida-Umehara C, Tarui H, Kuroiwa A, Yamada K, Isobe T, Ando J, Fujiwara A, Hirao Y, Nishimura O, Ishijima J, Hayashi A, Saito T, Murakami T, Murakami Y, Kuratani S, Agata K (2005) Highly conserved linkage homology between birds and turtles: birds and turtle chromosomes are precise counterparts of each other. Chromosome Res 13:601–615
Nanda I, Shan Z, Schartl M, Burt DW, Koehler M, Nothwang H-G, Grützner F, Paton IR, Windsor D, Dunn I, Engel W, Staeheli P, Mizuno S, Haaf T, Schmid M (1999) 300 million years of conserved synteny between chicken Z and human chromosome 9. Nat Genet 21:258–259
Nanda I, Zend-Ajusch E, Shan Z, Grützner F, Schartl M, Burt DW, Koehler M, Fowler VM, Goodwin G, Schneider WJ, Mizuno S, Dechant G, Haaf T, Schmid M (2000) Conserved synteny between the chicken Z sex chromosome and human chromosome 9 includes the male regulatory gene DMRT1: a comparative (re)view on avian sex determination. Cytogenet Cell Genet 89:67–78
Nanda I, Haaf T, Schartl M, Schmid M, Burt DW (2002) Comparative mapping of Z-orthologous genes in vertebrates: implications for the evolution of avian sex chromosomes. Cytogenet Genome Res 99:178–184
Nishida-Umehara C, Fujiwara A, Ogawa A, Mizuno S, Abe S, Yoshida MC (1999) Differentiation of Z and W chromosomes revealed by replication banding and FISH mapping of sex-chromosome-linked DNA markers in the cassowary (Aves, Ratitae). Chromosome Res 7:635–640
Nishida-Umehara C, Tsuda Y, Ishijima J, Ando J, Fujiwara A, Matsuda Y, Griffin DK (2007) The molecular basis of chromosome orthologies and sex chromosomal differentiation in palaeognathous birds. Chromosome Res 15:721–734
Ogawa A, Murata K, Mizuno S (1998) The location of Z- and W-linked marker genes and sequence on the homomorphic sex chromosomes of the ostrich and the emu. Proc Natl Acad Sci U S A 95:4415–4418
Ohno S (1967) Sex chromosomes and sex-linked genes. Springer, Berlin
Olmo E, Signorino G (2005) Chromorep: a reptiles chromosomes database. Internet references. Retrieved from: http://193.206.118.100/professori/chromorep.pdf
Ota H (1989a) A review of the geckos (Lacertilia: Reptilia) of the Ryukyu Archipelago and Taiwan. In: Matsui M, Hikida T, Goris RC (eds) Current herpetology in east Asia. Herpetological Society of Japan, Kyoto, pp 222–261
Ota H (1989b) Karyotypes of five species of Gekko (Gekkonidae: Lacertilia) from East and Southeast Asia. Herpetologica 45:438–443
Ota H, Hikida T, Nabhitabhata J, Panha S (2001) Cryptic taxonomic diversity in two broadly distributed lizards of Thailand (Mabuya macularia and Dixonius siamensis) as revealed by chromosomal investigations (Reptilia: Lacertilia). Nat Hist J Chulalongkorn Univ 1:1–7
Raymond CS, Shamu CE, Shen MM, Seifert KJ, Hirsch B, Hodgkin J, Zarkower D (1998) Evidence for evolutionary conservation of sex-determining genes. Nature 391:691–695
Sarre SD, Georges A, Quinn A (2004) The ends of a continuum: genetic and temperature-dependent sex determination in reptiles. BioEssays 26:639–645
Schmid M, Nanda I, Hoehn H, Schartl M, Haaf T, Buerstedde J-M, Arakawa H, Caldwell RB, Weigend S, Burt DW, Smith J, Griffin DK, Masabanda JS, Groenen MAM, Crooijmans RPMA, Vignal A, Fillon V, Morisson M, Pitel F, Vignoles M, Garrigues A, Gellin J, Rodionov AV, Galkina SA, Lukina NA, Ben-Ari G, Blum S, Hillel J, Twito T, Lavi U, David L, Feldman MW, Delany ME, Conley CA, Fowler VM, Hedges SB, Godbout R, Katyal S, Smith C, Hudson Q, Sinclair A, Mizuno S (2005) Second report on chicken genes and chromosomes 2005. Cytogenet Genome Res 109:415–479
Shetty S, Griffin DK, Graves JAM (1999) Comparative painting reveals strong chromosome homology over 80 million years of bird evolution. Chromosome Res 7:289–295
Shetty S, Kirby P, Zarkower D, Graves JAM (2002) DMRT1 in a ratite bird: evidence for a role in sex determination and discovery of a putative regulatory element. Cytogenet Genome Res 99:245–251
Shibusawa M, Nishibori M, Nishida-Umehara C, Tsudzuki M, Masabanda J, Griffin DK, Matsuda Y (2004) Karyotypic evolution in the Galliformes: an examination of the process of karyotypic evolution by comparison of the molecular cytogenetic findings with the molecular phylogeny. Cytogenet Genome Res 106:111–119
Singh L (1972) Evolution of karyotypes in snakes. Chromosoma 38:185–236
Smith CA, Sinclair AH (2004) Sex determination: insights from the chicken. BioEssays 26:120–132
Toda M, Hikida T, Ota H (2001) Discovery of sympatric cryptic species within Gekko hokouensis (Gekkonidae: Squamata) from the Okinawa Islands, Japan, by use of allozyme data. Zool Scr 30:1–11
Toda M, Sengoku S, Hikida T, Ota H (2008) Description of two new species of the genus Gekko (Squamata: Gekkonidae) from the Tokara and Amami Island groups in the Ryukyu Archipelago, Japan. Copeia 2008:452–466
Tsuda Y, Nishida-Umehara C, Ishijima J, Yamada K, Matsuda Y (2007) Comparison of the Z and W sex chromosomal architectures in elegant crested tinamou (Eudromia elegans) and ostrich (Struthio camelus) and the process of sex chromosome differentiation in palaeognathous birds. Chromosoma 116:159–173
Valenzuela N, Lance V (eds) (2004) Temperature-dependent sex determination in vertebrates. Smithsonian Books, Washington
van Tuinen M, Hedges SB (2001) Calibration of avian molecular clocks. Mol Biol Evol 18:206–213
Acknowledgments
This work was supported by Grants-in-Aid for Scientific Research (Nos. 15370001 and 16086201) from the Ministry of Education, Culture, Sports, Science and Technology, Japan.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Y. Hiraoka
Rights and permissions
About this article
Cite this article
Kawai, A., Ishijima, J., Nishida, C. et al. The ZW sex chromosomes of Gekko hokouensis (Gekkonidae, Squamata) represent highly conserved homology with those of avian species. Chromosoma 118, 43–51 (2009). https://doi.org/10.1007/s00412-008-0176-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00412-008-0176-2