
Abstract
Human prion diseases are rare neurodegenerative disorders related to prion protein misfolding that can occur as sporadic, familial or acquired forms. In comparison to other more common neurodegenerative disorders, prion diseases show a wider range of phenotypic variation and largely transmit to experimental animals, a feature that led to the isolation and characterization of different strains of the transmissible agent or prion with distinct biological properties. Biochemically distinct PrPSc types have been demonstrated which differ in their size after proteinase cleavage, glycosylation pattern, and possibly other features related to their conformation. These PrPSc types, possibly enciphering the prion strains, together with the naturally occurring polymorphism at codon 129 in the prion protein gene have a major influence on the disease phenotype. In the sporadic form, the most common but perhaps least understood form of human prion disease, there are at least six major combinations of codon 129 genotype and prion protein isotype, which are significantly related to distinctive clinical–pathological subgroups of the disease. In this review, we provide an update on the current knowledge and classification of the disease subtypes of the sporadic human prion diseases as defined by molecular features and pathological changes. Furthermore, we discuss the molecular basis of phenotypic variability taking into account the results of recent transmission studies that shed light on the extent of prion strain variation in humans.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Human prion diseases are grouped into three etiologic categories (Table 1). (a) The largest should be termed idiopathic, although it is generally referred to as “sporadic” and includes sporadic Creutzfeldt–Jakob disease (sCJD), sporadic fatal insomnia (sFI) and the recently characterized variably protease-sensitive prionopathy (VPSPr). It is hypothesized based on the prion theory [104] that sporadic prion diseases may be spontaneous disorders caused either by somatic mutations or by spontaneous prion protein (PrP) misfolding. Alternatively, the disease would arise through an infection from animals or other source, a hypothesis that as yet has not been completely ruled out. (b) Acquired prion diseases include cases accidentally transmitted from affected humans as in iatrogenic CJD (iCJD) or by ritual cannibalism as in kuru, or from BSE-infected animals as in variant CJD (vCJD). (c) The third group encompasses hereditary or familial diseases, which are categorized according to their distinctive clinical and pathological features as genetic CJD (gCJD), Gerstmann-Sträussler-Scheinker syndrome (GSS), and fatal familial insomnia (FFI). These are associated with a large number of mutations of the prion protein gene (PRNP) and clinically and pathologically are either indistinguishable from sporadic prion diseases or may show distinctive characteristics when associated with particular mutations.
The common pathologic denominators of human prion diseases are spongiform change and deposits of protease-resistant PrP (PrPSc). Spongiform change is often widespread and excessive, but in some disease subtypes may be very subtle or even absent. PrPSc deposition and spongiform change can appear in various forms. The vacuoles of the spongiform changes can be relatively small or larger or even coalescing, whereas PrP deposits may assume a synaptic, perivacuolar, perineuronal, or plaque-like morphology. PrPSc-containing amyloid plaques of different morphology such as kuru plaques, multicentric plaques or florid plaques are also seen as variants of prion pathology. While viewed as a continuum of pathological phenomena until not long ago, the wide phenotypic spectrum of human prion disease has been more recently classified in a number of distinct subtypes or variants where spongiform change and PrPSc deposition with or without amyloid plaques are associated in a specific way and often in a particular anatomic distribution. Simultaneously, two major types of PrPSc and subtypes thereof with distinct physicochemical properties, often combined with other truncated forms of abnormal PrPSc, have been described and the critical pathogenetic role played by the polymorphic codon 129 of PRNP in all forms of the disease has been demonstrated. It has become progressively clear that there is a close correlation between the (1) molecular features such as the PrPSc types as defined by western blot and the codon 129 genotype and the (2) pathological phenotype defined by the regional distribution and combination of forms of spongiform change and PrPSc deposition as well as (3) clinical features such as duration of disease, age at onset and clinical signs and symptoms.
This review, focusing on the sporadic human prion diseases, provides an update on the current knowledge and classification of the disease subtypes as defined by molecular features and pathological changes. Furthermore, we discuss the molecular basis of phenotypic variability with special reference to the results of recent transmission studies that shed light on the extent of strain variation in human prion disease.
Historical background
Soon after Jakob’s description of the disease [44] that bears his and Gerhard Creutzfeldt’s names, the phenotypic heterogeneity of CJD became evident, and a number of reports appeared describing a variety of clinical manifestations and distributions of pathological lesions. Heidenhain [38] in 1929 reported three CJD cases, which he believed to have distinctive features. These patients were characterized by shorter disease duration than Jakob’s cases and two of them had cortical blindness associated with a more prominent pathology in the occipital cortex. In two of the three focal spongiform degeneration was also present. Later, Meyer et al. [74] also reported cases that showed prominent visual disturbances, and concluded that they represent a specific variant of CJD, for which they proposed the term amaurotic or Heidenhain variant.
In 1939, Stern [119] published the case of a 51-year-old man with a rapidly progressive dementia of 9 months duration who at autopsy showed extensive degeneration of the thalamus and, to a lesser extent, of the cerebral cortex. Reports of similar cases followed [29, 112], some of them by authors who considered these patients as being affected by a special subtype of CJD, for which they proposed the term “thalamic” variant [29].
Between 1950 and 1960, several developments took place. Most importantly, the spongiform change was increasingly recognized as the most specific lesion of CJD and became the major pathologic criteria for the diagnosis of the disease. This coincided with the introduction of hematoxylin and eosin as standard staining, which allowed a much better visual detection of the spongiform changes. In addition, after the description by Nevin and its collaborators [81] of a large series of cases that clinically showed rapidly progressive dementia and widespread myoclonic jerks, and, pathologically, prominent spongiform vacuolation, increasing diagnostic importance was given to the triad of progressive dementia, myoclonus, and spongiform changes.
Meanwhile, the attempts to classify the disease in distinct variants on the basis of clinical or pathological criteria have continued, and an array of eponyms has been proposed based on clinical presentations or topographic distributions of lesions. For example, Alema and Bignami [2] distinguished a myoclonic form, an amaurotic form (as described by Heidenhain), a transitional form (also with myoclonus), a dyskinetic form, an ataxic form, and an amyotrophic form. Other authors classified the CJD subtypes according to the topographical distribution of the most significant pathology [125]. On this basis, they distinguished the cortical type, the optic type, the striatal type, the thalamic type, the nucleus niger type, and the anterior horn (amyotrophic) type.
In 1965, Brownell and Oppenheimer [12] described four cases, which together with six other cases reported in the literature between 1955 and 1961 seemed to constitute a well-defined clinical and pathological entity. According to these authors, in each of these cases, the disease presented with ataxia, while dementia and abnormal movements appeared later. Pathologically, all showed a selective cell loss in the granular layer of the cerebellum, in addition to typical lesions in the cerebral cortex, striatum, and thalamus. They proposed the term ataxic form of CJD for this entity. Other authors have subsequently adopted this term, and described similar cases [32, 45, 47, 56]. In the early 1970s, similarities between kuru and CJD were underlined with the report of CJD cases characterized by the presence of unicentric amyloid plaques indistinguishable from those described in kuru [1, 21, 60, 111].
Since 1980, several CJD cases characterized by a relatively long duration of illness, and marked alterations of the white matter in addition to the typical neuropathological features of classic CJD have been reported under the designation of panencephalopathic variant [7, 24, 77, 98]. These cases have been consistently found in the Japanese population but have been rarely reported in Europe and in the United States.
Despite significant advances in the understanding of the molecular biology of prions, including the discovery of mutations linked to familial CJD, the 1980s and early 90s brought little progress to the definition of the phenotypic variability of sporadic CJD and its molecular basis. The clinical spectrum of sporadic CJD was further defined in large clinical cohorts [10, 127], but this was not accompanied by detailed neuropathological studies. In addition, there was an increasing tendency to consider the phenotypic variability of sporadic CJD as part of a continuous spectrum with limitless possible combinations [10]. This perception, however, significantly changed after 1992. Following the initial demonstration by different groups of a correlation between the valine allele at codon 129 and the presence of a plaque-like pattern of PrP deposition [25, 27, 76, 113], it has been shown that two types of PrPSc with distinct physicochemical properties in conjunction with the codon 129 genotype specify distinct clinico-pathological variants of sporadic CJD [91]. This has led to a complete re-examination of the clinical and pathological spectrum of sporadic CJD on the basis of a comprehensive and systematic analysis of molecular, clinical and pathological features.
The molecular basis of phenotypic variability in sporadic prion disease
The human prion protein (PrPC) and prion protein gene (PRNP)
Human cellular PrP (PrPC) is a glycoprotein of 253 amino acids before cellular processing [59] with an 85–90% homology to PrP of other mammalian species. PrPC is a membrane protein expressed not only in neurons, but also in astrocytes and a number of other cells [58, 80]. Mature PrPC is attached to the cell surface by a GPI anchor and undergoes endocytosis and recycling. There are two N-glycosylation sites that are glycosylated differently in distinct human disease variants. The N-terminal moiety of the protein contains an octapeptide repeat [(PHGGGWGQ) × 4], which has been suggested to function in copper binding and neuroprotection [9, 39, 42, 75]. PrPSc is generated from PrPC in a post-translational process and, according to the prion hypothesis, it is part and parcel of the infectious agent, the prion. NMR structural studies have shown that the C-terminal half of PrPC contains a two-stranded antiparallel β-sheet (S1 and S2) and three α-helices, whereas the N-terminal moiety is thought to have no definite structure in aqueous solution [106]. PrPC and PrPSc seem to differ mainly in their folded structures. PrPSc purified from hamster brain consisted of 42% α-helical and only 3% β-sheet structure, whereas PrPSc purified from scrapie-infected hamster brain is composed of 30% α-helix and 43% β-sheet [88]. When compared to PrPC, PrPSc shows increased protease resistance, is insoluble in aqueous solution with detergents, and tends to form fibrils that show birefringence after binding of Congo red. The human PRNP is located on the short arm of chromosome 20 and has a simple genomic structure of two exons and a single intron of 13 kb. The entire protein-coding region is located in exon 2. In families with inherited prion diseases, different point mutations and insertion mutations have been described in the open reading frame (ORF) of PRNP.
Polymorphisms of PRNP and susceptibility to sCJD
A polymorphism for methionine (ATG) or valine (GTG) at codon 129 of PRNP is a determining factor for susceptibility to acquired and idiopathic prion disease. A number of studies have demonstrated that methionine homozygotes are more susceptible to sCJD whereas heterozygotes are less susceptible [62, 86, 108, 130]. In a large study of 300 sCJD cases and 544 controls in Europe and the USA in the normal population, 37% were MM, 51% were MV, and 12% were VV. In contrast among the sCJD cases, 71.6% were MM, 11.7% were MV, and 16.7% were VV [94].
Heterozygosity at another polymorphic PRNP codon, E219K, may be associated with resistance to sCJD. This notion is based on a Japanese study in which 6% of the population were found to show the polymorphism [51] and an additional study of 20 definite and 65 probable sCJD cases that were all homozygous EE [115]. However, a probable case of iCJD with the E219K polymorphism was identified in that study.
A number of additional polymorphisms of PRNP have been described. There are silent polymorphisms with no recognizable influence on disease susceptibility or disease phenotype. Among these are the A117A (GCA->GCG) and G124G (GGC->GGG) silent changes [43, 131]. Loss of one octarepeat has been reported with no recognizable influence on disease susceptibility [46, 87], while the loss of two octarepeats has been reported in CJD [6, 18].
It has been hypothesized that other polymorphisms outside the PRNP ORF, but nevertheless altering the expression of PrPC, could influence the susceptibility to CJD. Mead and co-workers [72] identified and analyzed 56 polymorphic sites within 25 kb of the PRNP ORF. They found extensive linkage disequilibrium around PRNP and were able to demonstrate the existence of 11 major European PRNP haplotypes. One common haplotype was over-represented in patients with sCJD but not vCJD or iCJD. There seems to be a significant independent association between sCJD and a polymorphism upstream of PRNP exon 1 in addition to the susceptibility conferred by codon 129. In a different approach, McCormack et al. [71] defined upstream and intronic regulatory regions of PRNP; subsequent sequencing of these regions revealed three polymorphisms that were more common in sCJD than controls. Thus, it seems likely that polymorphisms in the vicinity of PRNP ORF, probably in regulatory parts of PRNP, may be risk factors for sCJD in addition to the polymorphism at codon 129.
The pathological and clinical features of all forms of human prion disease are severely influenced by the codon 129 polymorphism. For example, in sCJD, plaque-like deposits and kuru plaques are only found in cases with at least one valine allele (see detailed discussion below).
Molecular typing of PrPSc
PrPSc types in sporadic human prion disease
Several lines of evidence indicate that human PrPSc exists in a number of molecular subtypes that show differences in their size and glycosylation pattern, degree of protease resistance, aggregation state, or conformational stability. However, the structural basis and functional consequences of this phenomenon still remain to be established. In particular, the question of whether the PrPSc subtypes fully encipher prion strain properties (i.e. the capability to reliably induce a specific disease phenotype in a given host genotype after transmission), or only reflect differing PrP molecular interactions, remains unresolved. Whatever the case, there is wide agreement that each particular PrPSc variety correlates with specific clinical and neuropathological features of the disease. In human prion disease, it has been convincingly shown that there is a very strong correlation between two PrPSc types with distinct physicochemical properties and the PRNP codon 129 genotype on one hand, and the pathological and clinical disease phenotype on the other [36, 94, 96]. In a comprehensive analysis of 300 sCJD cases, six subtypes of sCJD with distinctive phenotypic features were identified, each resulting from a specific codon 129 genotype/PrPSc type combination with two exceptions. MM1 and MV1 cases were phenotypically indistinguishable and, therefore, merged in one subtype (MM/MV 1), whereas, on the contrary, the combination of codon 129 MM and PrPSc type 2 was associated with two phenotypes with distinctive histopathological features in the cerebral cortex and in the thalamus and designated accordingly [MM2-cortical (=MM 2C) and MM2-thalamic (=MM 2T)] [94].
Based on the electrophoretic mobility of the fragment of PrPSc that is obtained after proteinase K (PK) digestion, Parchi et al. [90, 91] originally distinguished two major human PrPSc types (Fig. 1a). PrPSc type 1 has a relative electrophoretic mobility of 21 kDa and a primary cleavage site at residue 82 while PrPSc type 2 has a relative molecular mass of 19 kDa and a primary cleavage site at residue 97 [97]. Interestingly, it has also been found that PrPSc types 1 and 2 characterize all subtypes of CJD, independent of the apparent etiology of the disease, i.e. sporadic, inherited or acquired by infection [90, 97], thus suggesting a common mechanism of PrPSc formation.

Heterogeneity of PrPSc biochemical properties in sporadic CJD. a Western blot of proteinase K-treated brain homogenates run in a Tris–glycine PAGE 13% gel (15 cm long) and stained with the 3F4 mAb (residues 109–111). The three bands are composed of (from top to bottom) diglycosylated PrP, monoglycosylated PrP, and unglycosylated PrP. The panel shows two distinct patterns of human PrPSc, which differ with respect to the electrophoretic mobility of the protein core. Type 1 has a relative molecular mass of 21 kDa, whereas type 2 has a mass of 19 kDa. The mobility of the core varies between the two PrPSc types because different numbers of amino acids are removed from the N-terminal of PrP by proteinase K digestion. b Distinction of subtypes of PrPSc type 1 and type 2. In the upper panel, samples are run in a Tris–glycine PAGE 13% gel (6.5 cm long) and stained with 3F4. PrPSc type 2 from MV cases shows a unique doublet band that differs from the other type 2 proteins, while type 1 PrPSc from VV cases migrates faster than type 1 PrPSc from MM1 subjects. However, this difference is best detected when samples are run in a Tris–glycine PAGE 15% gel (15 cm long) (compare sample marked with asterisk between the upper panel and the one indicated by the arrow). c Western blot profiles of PrPSc (3F4 antibody) from sCJD subjects with PrPSc types 1 and 2 co-occurrence carrying different codon 129 PRNP genotypes. PrPSc type 1 from a MM1 case, and PrPSc type 2 from a VV2 case are included as controls. The PrPSc profile associated with the sCJDMV 2K is also shown. Note that the upper band of the PrPSc doublet in sCJDMV 2K migrates faster than the 21 kDa type 1 band. d Upper panel PK-treated and deglycosylated samples from sCJD MM1 and MM 2C probed with SAF60 (residues 157–161). In addition to the PrPSc types 1 and 2 cores, a 18.5 and a 13 kDa fragment are seen in the MM1 sample, whereas only a 17 kDa fragment is seen in MM 2C. d Lower panel Samples from a MM1 and a VV1 subject were PK treated in denaturing conditions and deglycosylated. Membranes were probed with an antiserum against the C-terminus (residues 220–231). An additional truncated fragment of 16 kDa is generated in these conditions in the MM1 sample. Approximate molecular masses in all panels are in kiloDaltons
Following this original classification [94], molecular PrPSc typing in human CJD attracted considerable interest in the scientific community as the molecular basis of prion strains constitutes a central issue of prion biology. In particular, molecular classification schemes consisting of more than two principal PrPSc types were proposed and claimed to better reflect phenotypic heterogeneity in human prion disease. Specifically, different typing systems distinguishing two PrPSc isoforms within MM subjects carrying the PrPSc type 1 described by Parchi et al. [91] were published [22, 126, 133]. In this regard, some confusion was caused by the fact that the same nomenclature proposed by Parchi et al. (MM1, MM2, etc.) was used to identify putative PrPSc forms that do not correspond to the respective types in the classification by Parchi et al. Besides the problem of the inappropriate nomenclature, recent insights into the biochemistry of PrPSc cleavage by proteases have raised doubts on the biological relevance of further distinguishing the MM1 subtype in two molecular subtypes. In a systematic analysis of the potential pitfalls regarding PrPSc typing by western blot, it was found that there are significant pH variations among CJD brain homogenates prepared in the standard buffers used in these studies, and that these variations in pH influence both PK activity and the size of the PK-resistant PrPSc fragment [15, 83]. In particular, it was shown that the variations in size that distinguish between PrPSc types 1 and 2 in the “London” typing system [22, 126] reflect differences in the pH of the brain homogenates analyzed, since they disappear by converging to one molecular type (type 1 of Parchi et al. classification), when analyzed at the same pH [83].
To test the reliability of a molecular classification based on PrPSc typing and PRNP genotyping, a collaborative study between seven laboratories involved in prion disease surveillance has recently been performed [95]. Sixteen well-characterized brain specimens from sCJD, vCJD and sFI cases were distributed blindly for molecular characterization according to the classification by Parchi et al. [83, 94] together with detailed protocols for sample preparation and western blot analysis. Using these standard conditions, the inter-institutional agreement between laboratories in the classification of samples was excellent, indicating that the general technique and in particular the classification system based on the distinction between types 1 and 2 [83, 94] are robust and represent a reliable basis for diagnostic and epidemiologic purposes.
The identification of an excess of pathological phenotypes and prion strains (see paragraph on transmission studies) with respect to the PrPSc types 1 and 2 dichotomy has led more recently to further attempts to identify PrPSc properties that would correlate with the disease phenotype. The study of PrPSc in rigorously controlled experimental conditions using highly sensitive gel electrophoresis technique led to the demonstration that PrPSc types 1 and 2 are indeed heterogeneous species, which can be further distinguished into molecular subtypes that correspond to the current histopathological classification of sCJD. Using a standardized high buffer strength for brain homogenization, PK digestion at pH 6.9 with a high enzyme concentration, and long running gels, Notari et al. [83] showed that distinctive PrPSc properties can be found in sCJD phenotypes sharing the same PrPSc type. For example, (a) PrPSc type 2 from MV cases shows a unique doublet band that differs from PrPSc type 2 in MM and VV cases, and (b) type 1 PrPSc from VV cases migrates faster than type 1 PrPSc from MM1 and MV1 samples when PK digestion is performed at pH under 7.2 (Fig. 1b). However, the recognition of these subtle differences between subtypes of the type 1 and type 2 groups, respectively, was less satisfactory when tested among CJD surveillance laboratories, probably because it requires highly standardized gel electrophoresis protocols that are unsuitable for routine diagnostic needs [95].
Co-occurrence of PrPSc types 1 and 2
Interestingly, PrPSc types 1 and 2 have also been found to co-occur in the same brain. Following the original report of this phenomenon [94, 105], a series of studies indicated that the prevalence of cases with co-occurrence of PrPSc types 1 and 2 varies between 12 and 44% [36, 37, 66, 105, 110, 124]. This finding has led to the proposition that the incidence of sCJD cases with both type 1 and 2 PrPSc reflects the extent of brain sampling and the sensitivity with which a minority type may be detected in the presence of larger amounts of the other protein [36]. Following this reasoning, subsequent studies that were based on specific antibodies recognizing an epitope between residues 82 and 96 (i.e. not detecting type 2) reached the conclusion that virtually all cases of sCJD characterized by the presence of PrPSc type 2 also contain PrPSc type 1 [103, 132]. Based on these findings, it was suggested that PrPSc typing is a quantitative, not a qualitative, phenomenon. However, recent studies have addressed important technical aspects relevant to this issue [14, 84]. It has been shown that there are pitfalls associated with the use of “type 1 selective” antibodies. Especially, when relatively mild digestion conditions are used, these antibodies are incapable of distinguishing the bona fide types 1 and 2 “core” fragments from the fragments obtained by incomplete protease digestion as there are several PK cleavage sites in the PrPSc N-terminus [84].
The issue of co-occurrence of PrPSc types has been recently re-examined comprehensively with an in-depth assessment of PrPSc type concurrence and its effect on disease phenotype in a large series of sCJD cases. To this aim, an extensive brain sampling and the most accurate methodology available for the detection of the PrPSc type concurrence, which provides good sensitivity combined with absolute specificity (i.e. the distinction between the bona fide types 1 and 2 “core” fragments from the fragments that are produced by incomplete protease digestion), has been applied [84]. Regarding sensitivity, the use of gels with increased resolution and a relatively high PK activity led to the finding that the detection sensitivity of either type 1 or 2, when concurrent, can be raised to amounts corresponding to 3–5% of the total PrPSc signal (i.e. weak band of one type/total PrPSc) [84]. Furthermore, the use of 1E4, a recently characterized monoclonal antibody with a higher affinity for type 2 than for type 1, allowed to increase the protein loading in the gel and led to a better identification of PrPSc type 2 when present in relatively low amounts within the mixture [14, 96]. The results obtained in a largely unselected series of 200 cases demonstrated an overall prevalence of PrPSc types 1 and 2 concurrence of 35% [96]. Although co-occurrence was detected in all codon 129 genotypes (Fig. 1c), PrPSc types 1 and 2 co-occurred more frequently in MM (43%) than in MV (23%) or VV (15%) subjects. Most MM and MV cases with mixed type 1 + 2 had PrPSc type 1 in all areas and PrPSc type 2 detectable only focally, usually in the cerebral cortex or the thalamus, whereas only a small group of subjects showed a predominant PrPSc type 2 accumulation. In the latter group, PrPSc type 2 was the dominant type in most areas, whereas the cerebellum consistently showed PrPSc type 1. Among the few VV cases with concurrent PrPSc types, type 2 was always dominant and detectable in almost all samples, whereas type 1 was usually present focally in the cerebral cortex, in the striatum, or in both. Taken together, these results indicate that the deposition of either type 1 or 2, when concurrent, is not random and always characterized by the co-existence of phenotypic features previously described for the “pure” subtypes. Given the importance of the recognition and correct classification of such cases for future epidemiological and transmission studies aimed to identify the etiology and extent of strain variation in sCJD, a protocol of PrPSc typing has been proposed that is based on analysis of at least four critical regions such as the temporal, parietal and occipital neocortices, and medial thalamus [96].
Truncated PrPSc fragments
Although for many years, full-length PrPSc and its core PrP27-30 were thought to be the only prion-specific PrP components, it has been more recently recognized that the abnormal PrP that accumulates in the diseased brains comprises additional truncated protein fragments. Unglycosylated PrPSc fragments of 7–8 kDa, truncated both the N termini and the C termini, were first identified in GSS [93, 101, 121]. Later, Capellari et al. [17] and Zou et al. [136] described two C-terminal fragments with a relative mass of about 12 and 13 kDa (PrP-CTF12/13), originating from PrPSc cleavage at residues 162–167 and 154–156, respectively. It has also been shown that the relative amount of PrP-CTF13 varies between sCJD subtypes; in particular, all PrPSc type 2-associated sCJD subtypes but the MM 2T show only traces of this fragment, whereas the peptide is present in significant amount in MM1 cases and is particularly abundant in VV1 subjects [85] (Fig. 1d).
Notari et al. also identified a novel C-terminally truncated PrPSc fragment showing an apparent molecular mass of either ~18.5 kDa (when associated with type 1) or ~17 kDa (when associated with type 2) (Fig. 1d). This fragment shares the primary N-terminal sequence with either type 1 or type 2 but lacks the very end of the C-terminus together with the GPI anchor (PrPAF 18.5–17) [85]. Finally, a fragment with an apparent molecular mass of about 16 kDa, which is only generated in partially denaturing conditions (DCF 16), has been detected in sCJD MM1/MV1 (Fig. 1d). Epitope mapping indicates that the fragment has an intact C-terminal end and is truncated in the region between residue 112 and residue 144. Taken together, these data suggest that each sCJD subtype can be associated with a specific profile of abnormal PrP fragments (PrP27-30, PrPAF 18.5–17, DCF 16, PrP-CTF12/13), possibly reflecting subtype-specific structural characteristics of the abnormal protein aggregate.
Glycosylation of PrPSc and molecular typing
Human PrP is post-translationally modified by the addition of two N-linked glycosylation moieties at residues 181 and 197. The type and combination of these sugar molecules can produce a high degree of heterogeneity, potentially affecting both protein conformation and intermolecular interactions [28, 118, 120]. Furthermore, the non-obligatory addition of one or two sugar chains results in three differently glycosylated isoforms (di-, mono-, or unglycosylated) in both PrPC and PrPSc. It has been recognized for a long time that PrPSc shows distinct ratios of the three differently glycosylated form (the so-called glycoform ratio) that correlate with prion disease subtypes. These differences are widely used as diagnostic tool for prion disease subtype recognition, especially between BSE and natural scrapie in sheep, or between vCJD and sCJD or iCJD in humans [5, 40, 90, 117]. In the large majority of CJD cases, PrPSc glycosylation is characterized by an over-representation of the monoglycosylated form. A rather grossly major distinction with undoubted diagnostic relevance has been introduced to distinguish the above-described “pattern A”, from “pattern B” characterized by a predominance of the fully glycosylated form, the latter being found in vCJD [22, 90] or in gCJD and FFI linked to the E200K or D178N mutations, respectively [78, 97]. However, finer significant differences in PrPSc glycoform ratio have also been described among CJD subtypes with either “pattern A” or “pattern B” using either mono- or two-dimensional gel electrophoresis [41, 89, 94].
Since glycosylation is a co-translational process that is known to differ among cell types, and indeed the glycotype of PrPC also varies among different brain regions [61], the different glycoform ratios may reflect the involvement or the targeting of distinct neuronal populations, which would in turn lead to strain-specific lesion profiles [16, 26, 65, 116]. Alternatively, different prion strains may preferentially convert certain PrPC glycoforms. Data against the hypothesis of a critical role of PrP glycosylation for the maintenance of strain-dependent prion neurotropism have also been published [102]. Finally, in inherited prion diseases, the glycoform ratio of PrPSc may also reflect altered PrPC processing induced by specific PRNP mutations [17, 100].
PrPSc typing by methods other than western blot
Besides the strain typing approaches based on the analyses of the PrPSc fragments generated by PK cleavage and glycoform ratio, there have been several efforts to identify strain-related characteristics of PrPSc using novel approaches, such as the study of PrPSc solubility and aggregation size, degree of protease resistance or conformational stability. However, the large majority of such data were obtained in animals or in vitro studies [20, 64, 99, 107, 122, 123], whereas, to date, only little has been accomplished with human prions [3, 14, 54]. On ATR–FTIR spectra of purified PrPSc after PK digestion, the percentage of β-sheet secondary structure in purified PrPSc from six sCJD patients was evaluated and a positive correlation between amount of β-sheet and degree of protease resistance was observed [3], but no apparent correlation with the PrPSc type or disease phenotype. More recently, Kobayashi et al. [54] studied PrPSc aggregation in MM 1 and MM 2T sCJD (sFI) cases and found that PrPSc type 2 has a larger aggregation size than PrPSc type 1, a result which they also confirmed in mixed type 1/type 2 cases. Moreover, Cali et al. by comparing the PrPSc conformational stability between sCJD MM 1 and sCJD MM 2C have recently observed that PrPSc type 1 is more stable than PrPSc type 2. However, this difference seems to be lost when PrPSc type 1 and type 2 co-occurred in the same brain [14]. Although of potential interest, the latter group of studies awaits further research, since they have been conducted on a limited number of cases and are only partially representative of the wide phenotypic spectrum of sporadic human prion disease.
Clinical and pathological types of sporadic prion disease (see also Table 2)
General neuropathological features (Figs. 2, 3)
Neuronal loss, spongiform change, astrocytic gliosis, and microglial activation are the defining features of sCJD observed by light microscopy. Neuronal loss may at times be difficult to assess without systematic morphometry or may be most severe with practically no discernible neurons left in the cerebral cortex and basal ganglia as well as extreme loss of cells of the internal granular layer of the cerebellum. Although the hippocampus, areas CA1–CA4, and the fascia dentata are spared from neuronal loss and spongiform change in the majority of cases, they are affected to some degree in sCJD brains of VV1, VV2, and MV2 patients (see below). The thalamus shows variable degrees of atrophy and is most severely affected in the MM 2T subtype (sFI), in particular the ventral anterior and dorsomedial nuclei. In the cerebellum, Purkinje cells appear normal in the vast majority of sCJD brains even with severe atrophy of the internal granular layer, but they sometimes show shrinkage and axonal swellings (torpedoes). In H&E-stained section, brain stem pathology is inconspicuous in most cases but shows spongiform change, nerve cell loss, and gliosis in the midbrain tectum and substantia nigra in VV2, MV2, and to a lesser degree in MM 2C cases.

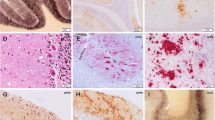
Histological findings in sCJD. a Small vacuoles are seen in the H&E stain in a MM/MV1 sCJD case. There are fine, partially opaque vacuoles in the neuropil. b H&E stain of confluent vacuoles as typically seen in MM 2C and mixed MM/MV 1 + 2C cases. c Slightly larger vacuoles than in MM/MV 1, often with a more distinct rim, are seen in VV1 cases. H&E. d Status spongiosus. There is almost complete neuronal loss with severe astrocytic gliosis and numerous shrinkage clefts and vacuoles in the remaining neuropil of the cortex. H&E stain. e A kuru plaque is shown in the cerebellar granular cell layer in a MV2 sCJD case. H&E stain. f Synaptic immunohistochemical PrPSc staining in the molecular and granular layer of the cerebellum. Delicate diffuse staining is seen in the molecular layer while in the granular cell layers the cerebellar glomeruli are stained

Histological findings in sCJD. a Confluent vacuoles with perivacuolar PrPSc staining as typically seen in MM 2C and MM/MV1 and MM 2C mixed cases. Typically dense deposits of PrPSc are seen surrounding confluent vacuoles. IHC with antibody L42. b Perineuronal PrPSc staining, which often includes neuronal staining, is typically observed in VV2 cases. IHC with antibody L42. c Plaque-like deposits in the cerebellum of a VV2 sCJD case. Plaque-like deposits are not visible in routine H&E- or PAS-stained sections. IHC with antibody L42. d The cerebellum in MM 2C may not show any PrP staining in IHC or may show focal patchy staining as in this case. e PET blot of the cerebellum of a VV2 case. The molecular and internal granular layers show a strong homogeneous staining, the Purkinje cells are negative, the white matter shows patchy immunohistochemical staining for PrPSc. f PET blot of the cortex of a VV2 case. A strong positive diffuse staining is seen in all cortical layers of the cortex more pronounced in upper layers while the lower layers also show focal perineuronal and some granular PrPSc deposits
Vacuolar changes comprise a number of descriptive forms. Most commonly, spongiform change consists of small, round, sometimes opaque vacuoles of 2–5 μm in diameter. Spongiform change may be found in the cerebral and cerebellar cortex, in the basal ganglia, thalamus, and brain stem. It is found in almost all cases of human prion diseases, but may be restricted to very small areas such as the entorhinal cortex in sFI. Typical spongiform change with small vacuoles must be distinguished from “confluent vacuoles” that are seen in MM 2C cases. Confluent vacuoles are observed in the cerebral neocortex and basal ganglia; they are absent from the cerebellum and brain stem as well as from the allocortex. Spongiform change has to be distinguished from status spongiosus. Status spongiosus was defined by Masters and Richardson in 1978 [69] and was described as being “… characterized by the appearance of cavitation of the neuropil in the presence of a dense glial meshwork. The cavitation was situated between glial fibers, and the cavities were of irregular size and shape. This lesion corresponds to what was originally described by Probst (1903), Fischer (1911) and Spielmeyer (1922)”. Status spongiosus is not specific for CJD, it is also occasionally observed in other degenerative diseases, in particular Pick’s disease and Alzheimer’s disease, and is seen in end-stage metabolic diseases including Leigh syndrome, and mitochondrial disorders, in particular MELAS. It should be distinguished from the laminar necrosis seen in anoxic-ischemic encephalopathies and boundary zone infarcts. Moreover, spongiform change, confluent vacuoles, and status spongiosus must be distinguished from other types of vacuolar changes, in particular spongy degeneration of the 2nd cortical layer in late stages of a variety of neurodegenerative diseases with severe cortical atrophy, including Alzheimer disease, Pick disease, motor neuron disease with dementia, and prion disease. Among the glial changes associated with the disease, microglial activation has attracted particular interest for its presumed pathogenetic role in neurodegeneration. Although still controversial [33], the idea that microglial activation is associated with a “neuroinflammatory state”, which contributes negatively to the development of neurodegeneration, has gained significant support in recent years. In prion disease, microglial activation parallels PrPSc deposition, and precedes both neuronal death and the appearance of clinical signs [30, 128]. Significant variation in the degree of microglial activation and its regional distribution occurs in both CJD and experimental prion disease, and there is good evidence indicating that this is influenced by both the prion strain and host genetic factors [4, 129].
PrPSc depositions
Light microscopy of H&E-stained sections allows the recognition of tissue deposits of PrPSc only when they lead to the formation of amyloid plaques such as kuru plaques. Various other forms of PrPSc depositions can be visualized using immunohistochemistry (IHC) and PET blotting [114].
Kuru plaques are compact PrPSc depositions that are visible in routine H&E- or PAS-stained sections, whereas plaque-like deposits are visible only on IHC staining with antibodies against PrP. Kuru plaques are pathognomonic for prion disease but are seen only in a minority of sCJD cases, where they are almost exclusively associated with heterozygosity for methionine and valine at codon 129 of PRNP and PrPSc type 2. In addition, they are typically seen in kuru and sometimes also in vCJD and GSS. In sCJD and kuru, kuru plaques are most commonly seen in the internal granule cell layer in the cerebellum, whereas they are difficult to find in the neocortex without IHC staining.
Kuru plaques must be distinguished from multicentric plaques which are typical of GSS and most often consist of a central large plaque that is surrounded by smaller “satellites”, and florid plaques, characteristic of vCJD that show a central core surrounded by a ring of spongiform change. In human prion diseases, they are encountered in high density in vCJD but are also seen in small numbers in a minority of iCJD cases after dura mater implantation.
Kuru plaques, multicentric plaques, and florid plaques can be seen in H&E-stained sections and have been shown to contain PrPSc by antibodies against PrP. In addition, a number of PrPSc deposits are only visible using IHC:
-
1.
Plaque-like depositions consist of small accumulations of PrPSc not visible in H&E-stained sections and are primarily seen in VV2 cases.
-
2.
Synaptic staining is a form of delicate and often hazy PrPSc deposition that was first described by Kitamoto [50]. It is commonly found in the molecular and granular cell layers in the cerebellum and in the isocortex in MM1 and MV1 cases.
-
3.
Perivacuolar staining is associated with large confluent vacuoles and is typical of MM 2C sCJD but also seen in many cases with mixed phenotypic features (MM/MV 1 + 2C and MV 2K + 2C see below).
-
4.
Perineuronal PrPScaccumulations are typically found in VV2 sCJD cases.
Immunohistochemical techniques for PrPSc detection do not differ significantly from other IHC applications. However, it is of paramount importance to use denaturing tissue pretreatments such as immersion in guanidium hydrochloride, autoclaving in H2O, and hydrolytic autoclaving in a solution of 10 mM HCl, or a combination of these pretreatments. Many groups use pretreatment with formic acid for enhancement of PrPSc staining [49] and tissue decontamination [11] at the same time. Pretreatment with PK is not regularly used. It often leads to enhanced PrP staining but the tissue dissolves in the process. PrPC is not seen after the described tissue pretreatment; therefore, PK pretreatment is not necessary for a distinction from PrPSc. PK treatment can conveniently be used in the PET blot technique in which nitrocellulose-bound PrPSc can be digested apparently without significant loss [114]. This is particularly useful in cases where minimal amounts of PrP are present in histological sections such as sFI.
Phenotypic variability of sporadic prion disease and its molecular basis
Six molecular and corresponding phenotypic subtypes of sCJD, one of which also designated as sFI, have been originally characterized based on a comprehensive clinical, pathological and molecular analysis of a large series of cases [94]. Later studies have confirmed the basis of this classification [15, 36, 66, 96, 110]. Furthermore, the spectrum of phenotypes with mixed pathological features and the co-occurrence of PrPSc types, which accounts for about one-third of all sCJD cases, have been recently more rigorously defined [14, 96]. The main features of both pure and mixed sCJD subtypes are summarized in Table 2.
sCJD types with “pure” molecular and clinico-pathological features
Type MM/MV 1
The most common phenotype is the MM/MV 1 type, which in its pure form comprises about 40% of all sCJD cases, and is seen in subjects who are homozygous for methionine or heterozygous at codon 129 and have PrPSc type 1 [94, 96]. The mean age at onset of symptoms is 70.1 years and the average clinical duration 4 months. Signs at onset, which are often combined, vary and include cognitive decline in about two-third of cases, ataxia in approximately one-third, language dysfunction, visual signs of central origin, myoclonus or other involuntary movements, each occurring in about one-fourth of patients. Neurological signs are unilateral at onset in about 25% of cases. Diffusion-weighted (DW)-MRI shows a signal increase in the basal ganglia in about 70% of cases, whereas the involvement of the cerebral cortex in at least three different regions is detected in half the patients. The absence of hippocampal and thalamic signal increase is a distinctive feature of this subtype [73]. Proteins 14-3-3 and total tau are markedly elevated in the CSF and their measurement provides a diagnostic pre-mortem test with a reported sensitivity that ranged from 91 to 100% [19, 23, 31, 109]. Periodic sharp-wave complexes (PSWCs) are recorded in the EEG in about 80% of cases, usually within the first 3 months of symptoms [23, 94]. Crucial for the presumed clinical diagnosis of this subtype are the very rapid course, which is usually already apparent a few weeks after onset, and the early presence of heterogeneous clinical signs indicating a multisystem involvement. A positive DW-MRI in both basal ganglia and cerebral cortex or the appearance of PSWCs in the EEG within the first 2 months of evolution further strengthens the clinical diagnosis of MM/MV 1.
This phenotype shows the classical pathology of CJD: spongiform change with small (2–10 μm) round to oval vacuoles, predominantly located to neural processes. In the cerebral neocortex, the vacuolation is seen in all layers and is often more prominent in the occipital lobe. The basal ganglia and thalamus are similarly affected, whereas the hippocampus and brain stem are remarkably spared. In the cerebellum, spongiform change is seen in the molecular layer, often in a focal distribution. IHC shows a synaptic and sometimes punctate type of PrPSc staining in the cerebral cortex, basal ganglia, thalamus and cerebellum. The hippocampus is most often negative.
Type VV 2
The second most common phenotype comprises about 15% of sCJD cases and includes subjects homozygous for valine at codon 129 with PrPSc type 2 (VV2) [94, 96]. The mean age at onset and clinical duration are 64.5 years and 6.3 months, respectively. Clinically, VV2 patients show prominent gait ataxia at onset, while dementia often occurs later in the course of the illness. Prominent myoclonus is absent in about one-third of the cases, and most patients also lack PSWCs in the EEG. DW-MRI shows involvement of the basal ganglia and thalamus, with the highest frequency among the sCJD subtypes. In contrast, the cerebral cortical signal increase is usually limited and most frequently found in the limbic cortex such as the cingulate gyrus rather than in the neocortex. The reported diagnostic sensitivity of 14-3-3 and total tau detection in the CSF varies from 84 to 100% [19, 23, 31, 109]. The presumed clinical diagnosis of the VV2 phenotype is raised in cases presenting with a rapidly progressive neurological syndrome of recent onset dominated by ataxia and other cerebellar signs in the absence of significant involvement of other systems. A positive DW-MRI in the basal ganglia, or both in the basal ganglia and thalamus (but not in the cerebral cortex) together with a “negative” EEG (e.g. no PSWCs), and a VV genotype at codon 129 further corroborate the clinical suspicion.
Spongiform change is microvacuolar, and in the neocortex it is typically confined to the deep layers (4–6). There is also prominent involvement of subcortical nuclei including the basal ganglia, thalamus and brain stem. The hippocampus is also remarkably affected in this type, particularly in the subiculum and CA1 regions. Severe spongiform change is noted in the molecular layer of the cerebellum and there is often marked atrophy of the granular layer. IHC shows plaque-like focal PrPSc deposits as well as prominent perineural staining in layers 4–6 in the neocortex and often in the CA4 region of the hippocampus. The cerebellum shows widespread plaque-like deposits in the molecular and granular layers and significantly in the white matter of the cerebellar folia. The brain stem also shows positivity for PrPSc.
Type MV 2K
The third most common phenotype affects about 8% of cases, and comprises the kuru-plaque variant, which is linked to MV at codon 129 and PrPSc type 2 (MV 2K) [94, 96]. The mean age at onset and clinical duration are 65.4 years and 15.8 months, respectively. Clinically ataxia and dementia are prominent. A long clinical duration of disease exceeding 2 years is not infrequent in this type and may raise diagnostic difficulties at the clinical stage. Typically, there are no PSWCs on EEG recording. DW-MRI shows, like in the VV2 type, a quite constant involvement of basal ganglia and thalamus. In particular, the thalamic signal increase is most frequently observed in the pulvinar, sometimes giving rise to a typical “pulvinar sign” [135], followed by the mediodorsal nuclei and the anterolateral nuclei. Involvement of the cerebral cortex is less frequent and most often includes the frontal lobes and cingulate gyri. The reported diagnostic sensitivity of 14-3-3 and total tau detection in the CSF is one of the lowest among the sCJD subtypes ranging from 57 to 89% [19, 23, 31, 109]. The presumed clinical diagnosis of the MV 2K phenotype is rather difficult in the early disease stages. However, it may become a reasonable clinical hypothesis when dealing with a progressive neurological syndrome with both cortical and cerebellar signs lasting at least 8 months and associated with an increased DW-MRI signal in the basal ganglia and thalamus and an EEG lacking PSWCs. A weakly positive or even negative 14-3-3 test in the CSF does not argue against the clinical suspicion of sCJD MV 2K.
The distinctive features of this type are cerebellar kuru plaques, while other pathological changes are similar to VV2 with more consistent plaque-like deposits. Kuru plaques are round, dense, PrPSc-positive amyloid plaques, which—in contrast to plaque-like deposits—are visible on routine HE staining. They are regularly found in the molecular and granular layers of the cerebellum but are less common in cerebral structures.
Type MM/MV 2C
Two distinct phenotypes affect subjects who are homozygous for methionine at codon 129 and show PrPSc type 2 [94, 96]. The first (MM2-cortical or MM 2C) comprises about 1% of the sCJD population. The mean age at onset in these subjects is 67.8 years, and the average disease duration 20 months. Clinically, these subjects present with cognitive impairment, while sustained myoclonus, visual signs and the typical EEG changes are usually absent. Cerebellar signs are mild or absent, even late in the course of the disease. A widespread DW-MRI cortical signal increase, which typically includes the temporal lobes, is characteristic of this subtype. Basal ganglia involvement is instead rather limited, whereas, compared to the MM1 type, a thalamic signal increase occurs more frequently in these cases. The reported diagnostic sensitivity of 14-3-3 and total tau detection in the CSF is relatively low and ranges between 61 and 78% [19, 23, 31, 109]. The presumed clinical diagnosis of MM 2C phenotype should be raised in case of a patient in his 50s or older suffering from a progressive neurological syndrome dominated by dementia and showing an increased DW-MRI signal that is restricted to the cerebral cortex. VV at codon 129 must be ruled out.
The hallmark of this type are widespread large confluent vacuoles in the cerebral cortex, with perivacuolar and focal coarse PrPSc staining on IHC. The basal ganglia and thalamus are less severely affected, the hippocampus is typically spared. Notably, the cerebellum shows no or focal pathological changes, depending on the disease duration. Recently, this phenotype was also described in at least one MV subject [96].
Type MM 2T
The second phenotype that is linked to PrPSc type 2 and methionine homozygosity also affects about 1% of subjects with sCJD [94]. This variant (MM2-thalamic or MM 2T) is virtually indistinguishable from FFI [79], and there is now convincing evidence that both the sporadic MM 2T variant and FFI are caused by the same prion strain (see below). Clinically, insomnia with inability to initiate and maintain sleep, frequent arousals, and enacted dreams is often an early and dominant symptom, particularly in subjects homozygotes for methionine at codon 129. Motor signs also develop but are generally more prominent in the heterozygous patients. They include diplopia, dysarthria, dysphagia, pyramidal signs and gait abnormalities. Myoclonus, spontaneous and evoked, is always found. As a rule, there are no PSWCs on EEG. MM 2T patients usually show no signal alterations on DW-MRI [35] and this can be attributed to the absence or limited extent of spongiform changes in the basal ganglia, thalamus, and cerebral cortex [68]. However, two recent studies [34, 67] have documented by combined DW-MRI and MRS a metabolic pattern indicating gliosis restricted to the thalamus, which may be quite specific for the MM 2T phenotype. The diagnostic sensitivity of 14-3-3 and total tau detection in the CSF is also unrewarding, since in most cases the CSF levels of both proteins do not reach the threshold for a positive test. The clinical suspicion of MM 2T phenotype is rather difficult and mainly based on the rapid progressive course and the presence of the sleep disturbances described above combined with motor signs such as diplopia or ataxia. The MM genotype at codon 129 corroborates the clinical impression.
Neuropathologically, there is prominent atrophy of the thalamus and inferior olive with only minor pathologic changes in other areas of the brain. Spongiform change may be absent or focal and is virtually limited to the cerebral cortex. Unique among sCJD subtypes is also the relative preservation of the striatum. PrPSc IHC and the PET blot may be positive only in the entorhinal cortex.
Type VV1
This rare phenotype affects about 1% of the sCJD population and is linked to PrPSc type 1 and VV at codon 129 [94]. VV1-affected subjects show a mean age at onset of 39.3 years, which is by far the youngest among the sCJD variants, and a mean duration of 15.3 months. VV1 subjects show a quite homogeneous and consistent phenotype, both clinically and pathologically. Symptoms at onset include progressive dementia, mainly of the fronto-temporal type, which may evolve for some months without significant motor signs, such as ataxia or myoclonus. In addition, typical PSWCs are absent on EEG examination. The MRI shows involvement of the cerebral cortex with the highest frequency among the sCJD subtypes. In contrast, a positive signal in the basal ganglia or in the thalamus is rarely observed. The reported diagnostic sensitivity of 14-3-3 and total tau detection in the CSF is the highest among the sCJD subtypes, with a 100% value in three out of four studies [19, 23, 31, 109]. The presumed clinical diagnosis of VV1 phenotype should be raised in case of a relatively young patient (<50 years) suffering from a progressive neurological syndrome dominated by dementia and showing an increased DW-MRI signal that is limited to the cerebral cortex. The detection of VV at codon 129 further strengthens the clinical diagnosis.
The pathology in these subjects predominantly affects the cortico-striatal regions, while other subcortical structures, including the cerebellum, are relatively spared [94]. Additional pathological features are the relative sparing of the occipital lobe in comparison to the frontal and temporal lobes. The presence of ballooned neurons in the cerebral cortex has also been described. Routine PrP immunohistochemistry shows a faint punctate staining in the cerebral cortex, despite the severe spongiform degeneration.
sCJD types with “mixed” molecular and pathological features
Type MM/MV 1 + 2C
The molecular type MM/MV 1 + 2C is by far the most common mixed type of sCJD. As many as 43% of all MM cases turned out to be mixed MM 1 + 2C cases on very close inspection [96]. This includes cases where confluent vacuoles are only found histologically in very restricted regions of the cerebral cortex or the thalamus making the demonstration of PrPSc types 1 and 2 co-occurrence by western blot very difficult. Given the rarity of the other mixed phenotypes, this is the only subtype in which clinical correlations were drawn. It was found that both disease duration and frequency of symptoms at onset in MM/MV 1 + 2C subjects vary according to the relative load of the two protein types. Thus, while the clinical phenotype in MM/MV 1 + 2C with a focal PrPSc type 2 deposition (up to 5 brain regions involved) does not significantly differ from those of the MM/MV 1 phenotype, the disease duration becomes significantly longer and cerebellar signs less frequent at onset with increasing type 2 load.
Similarly, the histopathology in this type shows varying relative amounts of features previously described in the pure MM/MV 1 and MM/MV 2C subtypes [94]. More precisely, this type is characterized by confluent foci of spongiform degeneration with large vacuoles in addition to the classic microvacuolation, and by a mixed synaptic/perivacuolar or coarse pattern of PrP deposition. There is a significant quantitative variation of these combined features from case to case which are overall more common in the cerebral cortex than in subcortical areas. The temporal, parietal and occipital cortices are the most affected cortical areas, whereas among subcortical nuclei, the thalamus most often shows the perivacuolar or coarse staining. Similarly, the lesion profile in MM/MV 1 + 2C largely overlaps with that of pure MM/MV 1 cases, but more often shows a relative sparing of the cerebellum, which is more typical of the MM/MV 2C subtype [96].
Type VV 2 + 1
At variance with patients carrying the M allele, the occurrence of mixed phenotypic features in VV cases is essentially based on the demonstration of PrPSc types 1 and 2 co-existence by immunoblotting. Indeed, the lesion profile and the pattern of PrP staining in the rare VV 2 + 1 type show features that are largely overlapping with those of typical VV2 sCJD cases. As the only distinctive feature, the two cases with the most significant type 1 accumulation showed a less consistent laminar pattern of spongiform degeneration in the cerebral cortex and a milder cerebellar pathology when compared to VV2 cases with similar disease duration [96].
Types MV 2K + 2C and MM 2T + 2C
The remaining two neuropathologic types with mixed features are also rare [96]. Furthermore, they are only identifiable histopathologically since they are characterized by the co-occurrence of phenotypic features that are associated with PrPSc type 2. The first comprises MV cases showing both kuru plaques in the cerebellum and confluent vacuoles associated with a perivacuolar and coarse focal PrPSc deposition in the cerebral cortex or less constantly other areas such as the striatum or the thalamus. According to the most recent nomenclature, this type has been defined as MV 2K + 2C.
The latter type named MM 2T + 2C, and so far reported in only two individuals carrying MM at codon 129, would include cases showing the co-occurrence of thalamic and inferior olivary atrophy and the confluent vacuoles in the cerebral cortex [96].
Thus, it seems that large confluent vacuoles and the perivacuolar pattern of PrPSc deposition may be found in sCJD associated with all phenotypes linked to MM or MV at codon 129. Although this observation remains difficult to interpret at present, it appears relevant for our future understanding of the molecular basis and the extent of strain variation in sCJD.
“Atypical” cases of sporadic human prion disease
Thousands of cases of sporadic prion disease have been diagnosed to date by surveillance centers worldwide and the vast majority of them have been shown to fit the classification system described above. However, a few exceptions were noted. Most of such “atypical” cases have been diagnosed in the United States and are now recognized to belong to a novel form of human sporadic prion disease designated VPSPr [137], while the remaining ones stand at the moment as a small group mainly published in case reports awaiting further observations and characterizations [52, 57, 134].
Characterization of human prion strains by experimental transmission
Prion strains, defined as isolates that cause distinct disease phenotypes upon transmission to syngenic animals, which persist on serial transmission, are believed to be the main cause of phenotypic diversity and are therefore also the main candidates to explain the heterogeneity of human prion disease. Since according to the prion theory sCJD may arise de novo, the term ‘strain’ may denote the particular clinical and pathological properties seen on experimental transmission and caused by a newly arising infectious agent.
Compared to the number of strain typing transmission studies conducted with the scrapie agent since the early 70s, the characterization of human prion strains has been significantly delayed. Early studies in non-human primates aimed to prove the transmissibility of human prion disease but lacked the characterization of the isolated strains. Furthermore, transmission of sCJD to wild-type mice did not often result in clinical disease [13], whereas transmissions to bank voles have only recently proved more efficient in terms of clinical outcome, although mainly for sCJD isolates linked to the M allele (MM1, MV1, and MM 2C) [82]. To facilitate the transmission of CJD, various lines of transgenic mice have been produced that express full-length or chimeric human and mouse PrP genes [40, 48, 55].
In the late 1990s, most transmission studies were prompted by the BSE crisis. This has led to the demonstration that vCJD and the large majority of BSE cases are caused by the same prion strain, which is distinct from prions found in sCJD, iCJD and kuru [13, 40, 63]. At about the same time, the first characterization of human sporadic prion isolates was accomplished in transgenic mice. Inocula from a single sFI (=MM 2T) case produced disease characteristics that differed from those induced by sCJD MM1 as well as gCJD E200K-129M, V210I-129M inocula [70].
Preliminary data concerning other sCJD subtypes became available later [55, 82]; but only very recently, the results obtained by a re-evaluation of the NIH series of prion disease transmitted to non-human primates and by more comprehensive experimental transmissions to transgenic mice have clarified the issue of the extent of strain variation in sporadic human prion disease and provided answers to the crucial question of how the current classification relates to different strains of sCJD [8, 53, 92].
The results of these studies indicate that, besides the MM 2T variant already mentioned above, four out of five of the other neuropathologic and molecular “pure” types of sCJD defined by Parchi et al. [94, 96] behave indeed as different strains of agent. Most importantly, sCJD MM1 and MV1 isolates have identical transmission properties, which significantly differ from those of sCJD VV2 or MV 2K. Furthermore, both the sCJD MM 2C and sCJD VV1 subtypes behave differently from each other and from the other isolates after transmission [8]. However, at variance with the sCJD MM1/MV1 and VV2/MV 2K strains, only single cases of sCJD MM 2C, MM 2T, and VV 1 have been examined, with the assumption that transmission characteristics of a single case will be representative of the particular subgroup. Thus, the results obtained for these rare subtypes, although clear and somehow expected, await confirmation.
Taken together, the results of the concluded transmission studies conducted with human affected tissues indicate that at least six distinct strains of prions affect humans:
-
1.
Strain A, related to the most common CJD phenotype or myoclonic variant (MM1/MV1 subtype).
-
2.
Strain B, related to the ataxic and kuru-plaque variants of CJD (VV2 and MV 2K subtypes), as well as kuru and the plaque-type of dura mater graft-associated CJD (p-dCJD).
-
3.
Strain C, related to vCJD.
-
4.
Strain D, related to fatal insomnia, either sporadic or familial (MM 2T subtype).
-
5.
Strain E, related to the cortico-striatal variant with synaptic deposits (VV1 subtype).
-
6.
Strain F, related to the cortical variant with confluent vacuoles (MM/MV 2C subtype).
These data, if confirmed, may provide the basis for a potential novel classification of human prion diseases based on the strain of the agent. Given the significant effect of host genotype at least for some strains (i.e. strain B), this variable should also be somehow taken into account. The rationale, applicability and convenience of such an approach compared to the current classification will likely be a matter of discussion among laboratories involved in human prion disease surveillance worldwide in the years to come.
Conclusion and future directions
The characterization of PrPSc types with distinct physicochemical properties has provided a powerful molecular tool for the diagnosis and characterization of human prion diseases. This molecular approach, combined with rigorous classic and modern neuropathology, has led to significant improvements of our knowledge of human prion diseases and to a systematic classification. It has become clear that the clinico-pathologic variability of sporadic human prion diseases parallels that of animal diseases such as scrapie or BSE in which phenotypic heterogeneity has been related to both agent strain variation and host genetic factors, namely the PRNP genotype. The current evidence indicates the existence of at least six distinct strains of prions in humans. In addition, the polymorphic codon 129, alone as in sporadic and acquired forms of CJD, or in conjunction with PRNP mutations as in gCJD and FFI, modulates strain susceptibility and phenotypic expression. Starting from this recently gained knowledge, future studies should aim to further define the molecular basis of prion strains and their distinct cellular targeting and the origin of sporadic prion disease in humans, which appear central to the understanding of these diseases. In the context of the prion hypothesis, efforts should be made to define reproducible conformational differences for each human prion strain that goes beyond the type 1/type 2 distinction and can be used for diagnostic purposes. Future epidemiologic studies should test the hypothesis of whether different human sporadic strains and their distribution in the population are linked to distinct risk factors. Furthermore, a more exhaustive study of CJD subjects carrying both the M and V allele at codon 129 should be performed, given the profoundly different behavior of these two alleles in terms of strain susceptibility and associated phenotypes. It is a quite intriguing observation that at least five different strains have been linked to the M allele, whereas only two have so far been associated with the V allele. If indeed the prion strains are enciphered by differences in protein conformation or misfolding, how can a single amino acid change at codon 129 cause such a difference between the M and V alleles in terms of protein misfolding? If sCJD is due to age-related stochastic events such as somatic mutations or spontaneous protein misfolding, why do rare strains such as those causing the VV1 and the MM 2T phenotypic variants preferentially affect people in their 30s and 40s who are significantly younger than those affected by the more common strains? These are the type of questions that the deep and systematic study of the naturally occurring disease in humans is generating and that researchers should try to answer in the near future.
References
Adams H, Beck E, Shenkin AM (1974) Creutzfeldt–Jakob disease: further similarities with kuru. J Neurol Neurosurg Psychiatry 37:195–200
Alema G, Bignami A (1959) Subacute degenerative presenile polioencephalopathy with akinetic stupor and decorticate rigidity with myoclonus (“myoclonic” variety of the Jakob–Creutzfeldt disease). Riv Sper Freniatr Med Leg Alien Ment 83(4 Suppl):1485–1623
Aucouturier P, Kascsak RJ, Frangione B, Wisniewski T (1999) Biochemical and conformational variability of human prion strains in sporadic Creutzfeldt–Jakob disease. Neurosci Lett 274:33–36
Baker CA, Lu ZY, Zaitsev I, Manuelidis L (1999) Microglial activation varies in different models of Creutzfeldt–Jakob disease. J Virol 73:5089–5097
Baron TG, Madec JY, Calavas D, Richard Y, Barillet F (2000) Comparison of French natural scrapie isolates with bovine spongiform encephalopathy and experimental scrapie infected sheep. Neurosci Lett 284:175–178
Beck JA, Mead S, Campbell TA, Dickinson A, Wientjens DP, Croes EA, Van Duijn CM, Collinge J (2001) Two-octapeptide repeat deletion of prion protein associated with rapidly progressive dementia. Neurology 57:354–356
Berciano J, Berciano MT, Polo JM, Figols J, Ciudad J, Lafarga M (1990) Creutzfeldt–Jakob disease with severe involvement of cerebral white matter and cerebellum. Virchows Arch A Pathol Anat Histopathol 417:533–538
Bishop MT, Will RG, Manson JC (2010) Defining sporadic Creutzfeldt–Jakob disease strains and their transmission properties. Proc Natl Acad Sci USA 107:12005–12010
Brown DR, Qin K, Herms JW, Madlung A, Manson J, Strome R, Fraser PE, Kruck T, von Bohlen A, Schulz-Schaeffer W, Giese A, Westaway D, Kretzschmar H (1997) The cellular prion protein binds copper in vivo. Nature 390:684–687
Brown P, Cathala F, Castaigne P, Gajdusek DC (1986) Creutzfeldt–Jakob disease: clinical analysis of a consecutive series of 230 neuropathologically verified cases. Ann Neurol 20:597–602
Brown P, Wolff A, Gajdusek DC (1990) A simple and effective method for inactivating virus infectivity in formalin-fixed tissue samples from patients with Creutzfeldt–Jakob disease. Neurology 40:887–890
Brownell B, Oppenheimer DR (1965) An ataxic form of subacute presenile polioencephalopathy (Creutzfeldt–Jakob Disease). J Neurol Neurosurg Psychiatry 28:350–361
Bruce ME, Will RG, Ironside JW, McConnell I, Drummond D, Suttie A, McCardle L, Chree A, Hope J, Birkett C, Cousens S, Fraser H, Bostock CJ (1997) Transmissions to mice indicate that ‘new variant’ CJD is caused by the BSE agent. Nature 389:498–501
Cali I, Castellani R, Alshekhlee A, Cohen Y, Blevins J, Yuan J, Langeveld JP, Parchi P, Safar JG, Zou WQ, Gambetti P (2009) Co-existence of scrapie prion protein types 1 and 2 in sporadic Creutzfeldt–Jakob disease: its effect on the phenotype and prion-type characteristics. Brain 132:2643–2658
Cali I, Castellani R, Yuan J, Al-Shekhlee A, Cohen ML, Xiao X, Moleres FJ, Parchi P, Zou WQ, Gambetti P (2006) Classification of sporadic Creutzfeldt–Jakob disease revisited. Brain 129:2266–2277
Cancellotti E, Bradford BM, Tuzi NL, Hickey RD, Brown D, Brown KL, Barron RM, Kisielewski D, Piccardo P, Manson JC (2010) Glycosylation of PrPC determines timing of neuroinvasion and targeting in the brain following transmissible spongiform encephalopathy infection by a peripheral route. J Virol 84:3464–3475
Capellari S, Parchi P, Russo CM, Sanford J, Sy MS, Gambetti P, Petersen RB (2000) Effect of the E200K mutation on prion protein metabolism. Comparative study of a cell model and human brain. Am J Pathol 157:613–622
Capellari S, Parchi P, Wolff BD, Campbell J, Atkinson R, Posey DM, Petersen RB, Gambetti P (2002) Creutzfeldt–Jakob disease associated with a deletion of two repeats in the prion protein gene. Neurology 59:1628–1630
Castellani RJ, Colucci M, Xie Z, Zou W, Li C, Parchi P, Capellari S, Pastore M, Rahbar MH, Chen SG, Gambetti P (2004) Sensitivity of 14-3-3 protein test varies in subtypes of sporadic Creutzfeldt–Jakob disease. Neurology 63:436–442
Caughey B, Raymond GJ, Kocisko DA, Lansbury PT Jr (1997) Scrapie infectivity correlates with converting activity, protease resistance, and aggregation of scrapie-associated prion protein in guanidine denaturation studies. J Virol 71:4107–4110
Chou SM, Martin JD (1971) Kuru-plaques in a case of Creutzfeldt–Jakob disease. Acta Neuropathol 17:150–155
Collinge J, Sidle KC, Meads J, Ironside J, Hill AF (1996) Molecular analysis of prion strain variation and the aetiology of ‘new variant’ CJD. Nature 383:685–690
Collins SJ, Sanchez-Juan P, Masters CL, Klug GM, van Duijn C, Poleggi A, Pocchiari M, Almonti S, Cuadrado-Corrales N, de Pedro-Cuesta J, Budka H, Gelpi E, Glatzel M, Tolnay M, Hewer E, Zerr I, Heinemann U, Kretszchmar HA, Jansen GH, Olsen E, Mitrova E, Alperovitch A, Brandel JP, Mackenzie J, Murray K, Will RG (2006) Determinants of diagnostic investigation sensitivities across the clinical spectrum of sporadic Creutzfeldt–Jakob disease. Brain 129:2278–2287
Cruz-Sanchez F, Lafuente J, Gertz HJ, Stoltenburg-Didinger G (1987) Spongiform encephalopathy with extensive involvement of white matter. J Neurol Sci 82:81–87
de Silva R, Ironside JW, McCardle L, Esmonde T, Bell J, Will R, Windl O, Dempster M, Estibeiro P, Lathe R (1994) Neuropathological phenotype and ‘prion protein’ genotype correlation in sporadic Creutzfeldt–Jakob disease. Neurosci Lett 179:50–52
DeArmond SJ, Sanchez H, Yehiely F, Qiu Y, Ninchak-Casey A, Daggett V, Camerino AP, Cayetano J, Rogers M, Groth D, Torchia M, Tremblay P, Scott MR, Cohen FE, Prusiner SB (1997) Selective neuronal targeting in prion disease. Neuron 19:1337–1348
Doh-ura K, Kitamoto T, Sakaki Y, Tateishi J (1991) CJD discrepancy. Nature 353:801–802
Endo T, Groth D, Prusiner SB, Kobata A (1989) Diversity of oligosaccharide structures linked to asparagines of the scrapie prion protein. Biochemistry 28:8380–8388
Garcin RBS, Khochneviss AA (1963) Le syndrome de Creutzfeldt–Jakob et les syndromes corticotrices du presenium (à l’occasion d’ observations anatomo-cliniques). Rev Neurol (Paris) 109:419–441
Giese A, Brown DR, Groschup MH, Feldmann C, Haist I, Kretzschmar HA (1998) Role of microglia in neuronal cell death in prion disease. Brain Pathol 8:449–457
Gmitterova K, Heinemann U, Bodemer M, Krasnianski A, Meissner B, Kretzschmar HA, Zerr I (2009) 14-3-3 CSF levels in sporadic Creutzfeldt–Jakob disease differ across molecular subtypes. Neurobiol Aging 30:1842–1850
Gomori AJ, Partnow MJ, Horoupian DS, Hirano A (1973) The ataxic form of Creutzfeldt–Jakob disease. Arch Neurol 29:318–323
Graeber MB, Streit WJ (2010) Microglia: biology and pathology. Acta Neuropathol 119:89–105
Haik S, Galanaud D, Linguraru MG, Peoc’h K, Privat N, Faucheux BA, Ayache N, Hauw JJ, Dormont D, Brandel JP (2008) In vivo detection of thalamic gliosis: a pathoradiologic demonstration in familial fatal insomnia. Arch Neurol 65:545–549
Hamaguchi T, Kitamoto T, Sato T, Mizusawa H, Nakamura Y, Noguchi M, Furukawa Y, Ishida C, Kuji I, Mitani K, Murayama S, Kohriyama T, Katayama S, Yamashita M, Yamamoto T, Udaka F, Kawakami A, Ihara Y, Nishinaka T, Kuroda S, Suzuki N, Shiga Y, Arai H, Maruyama M, Yamada M (2005) Clinical diagnosis of MM2-type sporadic Creutzfeldt–Jakob disease. Neurology 64:643–648
Head MW, Bunn TJ, Bishop MT, McLoughlin V, Lowrie S, McKimmie CS, Williams MC, McCardle L, MacKenzie J, Knight R, Will RG, Ironside JW (2004) Prion protein heterogeneity in sporadic but not variant Creutzfeldt–Jakob disease: UK cases 1991–2002. Ann Neurol 55:851–859
Head MW, Ironside JW (2009) Sporadic Creutzfeldt–Jakob disease: discrete subtypes or a spectrum of disease? Brain 132:2627–2629
Heidenhain A (1929) Klinische und anatomische Untersuchungen über eine eigenartige organische Erkrankung des Zentralnervensystems im Praesenium. Z Ges Neurol Psychiatr 118:49–114
Herms J, Tings T, Gall S, Madlung A, Giese A, Siebert H, Schurmann P, Windl O, Brose N, Kretzschmar H (1999) Evidence of presynaptic location and function of the prion protein. J Neurosci 19:8866–8875
Hill AF, Desbruslais M, Joiner S, Sidle KC, Gowland I, Collinge J, Doey LJ, Lantos P (1997) The same prion strain causes vCJD and BSE. Nature 389:448–450, 526
Hill AF, Joiner S, Beck JA, Campbell TA, Dickinson A, Poulter M, Wadsworth JD, Collinge J (2006) Distinct glycoform ratios of protease resistant prion protein associated with PRNP point mutations. Brain 129:676–685
Hornshaw MP, McDermott JR, Candy JM, Lakey JH (1995) Copper binding to the N-terminal tandem repeat region of mammalian and avian prion protein: structural studies using synthetic peptides. Biochem Biophys Res Commun 214:993–999
Hsiao K, Dlouhy SR, Farlow MR, Cass C, Da Costa M, Conneally PM, Hodes ME, Ghetti B, Prusiner SB (1992) Mutant prion proteins in Gerstmann-Straussler-Scheinker disease with neurofibrillary tangles. Nat Genet 1:68–71
Jakob A (1921) Über eigenartige Erkrankungen des Zentralnervensystems mit bemerkenswertem anatomischem Befunde (spastische Pseurosklerose-Encephalomyelopathie mit disseminierten Degenerationsherden). Dtsch Z Nervenheilkd 70:132–146
Jellinger K, Heiss WD, Deisenhammer E (1974) The ataxic (cerebellar) form of Creutzfeldt–Jakob disease. J Neurol 207:289–305
Jeong BH, Nam JH, Lee YJ, Lee KH, Jang MK, Carp RI, Lee HD, Ju YR, Ahn Jo S, Park KY, Kim YS (2004) Polymorphisms of the prion protein gene (PRNP) in a Korean population. J Hum Genet 49:319–324
Jones HR Jr, Hedley-Whyte ET, Freidberg SR, Baker RA (1985) Ataxic Creutzfeldt–Jakob disease: diagnostic techniques and neuropathologic observations in early disease. Neurology 35:254–257
Kitamoto T, Mohri S, Ironside JW, Miyoshi I, Tanaka T, Kitamoto N, Itohara S, Kasai N, Katsuki M, Higuchi J, Muramoto T, Shin RW (2002) Follicular dendritic cell of the knock-in mouse provides a new bioassay for human prions. Biochem Biophys Res Commun 294:280–286
Kitamoto T, Ogomori K, Tateishi J, Prusiner SB (1987) Formic acid pretreatment enhances immunostaining of cerebral and systemic amyloids. Lab Invest 57:230–236
Kitamoto T, Shin RW, Doh-ura K, Tomokane N, Miyazono M, Muramoto T, Tateishi J (1992) Abnormal isoform of prion proteins accumulates in the synaptic structures of the central nervous system in patients with Creutzfeldt–Jakob disease. Am J Pathol 140:1285–1294
Kitamoto T, Tateishi J (1994) Human prion diseases with variant prion protein. Philos Trans R Soc Lond B Biol Sci 343:391–398
Kobayashi A, Arima K, Ogawa M, Murata M, Fukuda T, Kitamoto T (2008) Plaque-type deposition of prion protein in the damaged white matter of sporadic Creutzfeldt–Jakob disease MM1 patients. Acta Neuropathol 116:561–566
Kobayashi A, Sakuma N, Matsuura Y, Mohri S, Aguzzi A, Kitamoto T (2010) Experimental verification of a traceback phenomenon in prion infection. J Virol 84:3230–3238
Kobayashi A, Satoh S, Ironside JW, Mohri S, Kitamoto T (2005) Type 1 and type 2 human PrPSc have different aggregation sizes in methionine homozygotes with sporadic, iatrogenic and variant Creutzfeldt–Jakob disease. J Gen Virol 86:237–240
Korth C, Kaneko K, Groth D, Heye N, Telling G, Mastrianni J, Parchi P, Gambetti P, Will R, Ironside J, Heinrich C, Tremblay P, DeArmond SJ, Prusiner SB (2003) Abbreviated incubation times for human prions in mice expressing a chimeric mouse–human prion protein transgene. Proc Natl Acad Sci USA 100:4784–4789
Kott E, Bornstein B, Sandbank U (1967) Ataxic form of Creutzfeldt–Jakob disease. Its relation to subacute spongiform encephalopathy. J Neurol Sci 5:107–113
Krebs B, Bader B, Klehmet J, Grasbon-Frodl E, Oertel WH, Zerr I, Stricker S, Zschenderlein R, Kretzschmar HA (2007) A novel subtype of Creutzfeldt–Jakob disease characterized by a small 6 kDa PrP fragment. Acta Neuropathol 114:195–199
Kretzschmar HA, Prusiner SB, Stowring LE, DeArmond SJ (1986) Scrapie prion proteins are synthesized in neurons. Am J Pathol 122:1–5
Kretzschmar HA, Stowring LE, Westaway D, Stubblebine WH, Prusiner SB, Dearmond SJ (1986) Molecular cloning of a human prion protein cDNA. DNA 5:315–324
Krucke W, Beck E, Vitzthum HG (1973) Creutzfeldt–Jakob disease. Some unusual morphological features reminiscent of kuru. Z Neurol 206:1–24
Kuczius T, Koch R, Keyvani K, Karch H, Grassi J, Groschup MH (2007) Regional and phenotype heterogeneity of cellular prion proteins in the human brain. Eur J Neurosci 25:2649–2655
Laplanche JL, Delasnerie-Laupretre N, Brandel JP, Chatelain J, Beaudry P, Alperovitch A, Launay JM (1994) Molecular genetics of prion diseases in France. French Research Group on Epidemiology of Human Spongiform Encephalopathies. Neurology 44:2347–2351
Lasmezas CI, Fournier JG, Nouvel V, Boe H, Marce D, Lamoury F, Kopp N, Hauw JJ, Ironside J, Bruce M, Dormont D, Deslys JP (2001) Adaptation of the bovine spongiform encephalopathy agent to primates and comparison with Creutzfeldt–Jakob disease: implications for human health. Proc Natl Acad Sci USA 98:4142–4147
Legname G, Baskakov IV, Nguyen HO, Riesner D, Cohen FE, DeArmond SJ, Prusiner SB (2004) Synthetic mammalian prions. Science 305:673–676
Levavasseur E, Laffont-Proust I, Morain E, Faucheux BA, Privat N, Peoc’h K, Sazdovitch V, Brandel JP, Hauw JJ, Haik S (2008) Regulating factors of PrP glycosylation in Creutzfeldt–Jakob disease—implications for the dissemination and the diagnosis of human prion strains. PLoS One 3:e2786
Lewis V, Hill AF, Klug GM, Boyd A, Masters CL, Collins SJ (2005) Australian sporadic CJD analysis supports endogenous determinants of molecular-clinical profiles. Neurology 65:113–118
Lodi R, Parchi P, Tonon C, Manners D, Capellari S, Strammiello R, Rinaldi R, Testa C, Malucelli E, Mostacci B, Rizzo G, Pierangeli G, Cortelli P, Montagna P, Barbiroli B (2009) Magnetic resonance diagnostic markers in clinically sporadic prion disease: a combined brain magnetic resonance imaging and spectroscopy study. Brain 132:2669–2679
Manners DN, Parchi P, Tonon C, Capellari S, Strammiello R, Testa C, Tani G, Malucelli E, Spagnolo C, Cortelli P, Montagna P, Lodi R, Barbiroli B (2009) Pathologic correlates of diffusion MRI changes in Creutzfeldt–Jakob disease. Neurology 72:1425–1431
Masters CL, Richardson EP Jr (1978) Subacute spongiform encephalopathy (Creutzfeldt–Jakob disease). The nature and progression of spongiform change. Brain 101:333–344
Mastrianni JA, Nixon R, Layzer R, Telling GC, Han D, DeArmond SJ, Prusiner SB (1999) Prion protein conformation in a patient with sporadic fatal insomnia. N Engl J Med 340:1630–1638
McCormack JE, Baybutt HN, Everington D, Will RG, Ironside JW, Manson JC (2002) PRNP contains both intronic and upstream regulatory regions that may influence susceptibility to Creutzfeldt–Jakob Disease. Gene 288:139–146
Mead S, Mahal SP, Beck J, Campbell T, Farrall M, Fisher E, Collinge J (2001) Sporadic—but not variant—Creutzfeldt–Jakob disease is associated with polymorphisms upstream of PRNP exon 1. Am J Hum Genet 69:1225–1235
Meissner B, Kallenberg K, Sanchez-Juan P, Collie D, Summers DM, Almonti S, Collins SJ, Smith P, Cras P, Jansen GH, Brandel JP, Coulthart MB, Roberts H, Van Everbroeck B, Galanaud D, Mellina V, Will RG, Zerr I (2009) MRI lesion profiles in sporadic Creutzfeldt–Jakob disease. Neurology 72:1994–2001
Meyer A, Leigh D, Bagg CE (1954) A rare presenile dementia associated with cortical blindness (Heidenhain’s syndrome). J Neurol Neurosurg Psychiatry 17:129–133
Mitteregger G, Vosko M, Krebs B, Xiang W, Kohlmannsperger V, Nolting S, Hamann GF, Kretzschmar HA (2007) The role of the octarepeat region in neuroprotective function of the cellular prion protein. Brain Pathol 17:174–183
Miyazono M, Kitamoto T, Doh-ura K, Iwaki T, Tateishi J (1992) Creutzfeldt–Jakob disease with codon 129 polymorphism (valine): a comparative study of patients with codon 102 point mutation or without mutations. Acta Neuropathol 84:349–354
Mizutani T, Okumura A, Oda M, Shiraki H (1981) Panencephalopathic type of Creutzfeldt–Jakob disease: primary involvement of the cerebral white matter. J Neurol Neurosurg Psychiatry 44:103–115
Monari L, Chen SG, Brown P, Parchi P, Petersen RB, Mikol J, Gray F, Cortelli P, Montagna P, Ghetti B et al (1994) Fatal familial insomnia and familial Creutzfeldt–Jakob disease: different prion proteins determined by a DNA polymorphism. Proc Natl Acad Sci USA 91:2839–2842
Montagna P, Gambetti P, Cortelli P, Lugaresi E (2003) Familial and sporadic fatal insomnia. Lancet Neurol 2:167–176
Moser M, Colello RJ, Pott U, Oesch B (1995) Developmental expression of the prion protein gene in glial cells. Neuron 14:509–517
Nevin S, Mc MW, Behrman S, Jones DP (1960) Subacute spongiform encephalopathy—a subacute form of encephalopathy attributable to vascular dysfunction (spongiform cerebral atrophy). Brain 83:519–564
Nonno R, Di Bari MA, Cardone F, Vaccari G, Fazzi P, Dell’Omo G, Cartoni C, Ingrosso L, Boyle A, Galeno R, Sbriccoli M, Lipp HP, Bruce M, Pocchiari M, Agrimi U (2006) Efficient transmission and characterization of Creutzfeldt–Jakob disease strains in bank voles. PLoS Pathog 2:e12
Notari S, Capellari S, Giese A, Westner I, Baruzzi A, Ghetti B, Gambetti P, Kretzschmar HA, Parchi P (2004) Effects of different experimental conditions on the PrPSc core generated by protease digestion: implications for strain typing and molecular classification of CJD. J Biol Chem 279:16797–16804
Notari S, Capellari S, Langeveld J, Giese A, Strammiello R, Gambetti P, Kretzschmar HA, Parchi P (2007) A refined method for molecular typing reveals that co-occurrence of PrP(Sc) types in Creutzfeldt–Jakob disease is not the rule. Lab Invest 87:1103–1112
Notari S, Strammiello R, Capellari S, Giese A, Cescatti M, Grassi J, Ghetti B, Langeveld JP, Zou WQ, Gambetti P, Kretzschmar HA, Parchi P (2008) Characterization of truncated forms of abnormal prion protein in Creutzfeldt–Jakob disease. J Biol Chem 283:30557–30565
Palmer MS, Dryden AJ, Hughes JT, Collinge J (1991) Homozygous prion protein genotype predisposes to sporadic Creutzfeldt–Jakob disease. Nature 352:340–342
Palmer MS, Mahal SP, Campbell TA, Hill AF, Sidle KC, Laplanche JL, Collinge J (1993) Deletions in the prion protein gene are not associated with CJD. Hum Mol Genet 2:541–544
Pan KM, Baldwin M, Nguyen J, Gasset M, Serban A, Groth D, Mehlhorn I, Huang Z, Fletterick RJ, Cohen FE et al (1993) Conversion of alpha-helices into beta-sheets features in the formation of the scrapie prion proteins. Proc Natl Acad Sci USA 90:10962–10966
Pan T, Colucci M, Wong BS, Li R, Liu T, Petersen RB, Chen S, Gambetti P, Sy MS (2001) Novel differences between two human prion strains revealed by two-dimensional gel electrophoresis. J Biol Chem 276:37284–37288
Parchi P, Capellari S, Chen SG, Petersen RB, Gambetti P, Kopp N, Brown P, Kitamoto T, Tateishi J, Giese A, Kretzschmar H (1997) Typing prion isoforms. Nature 386:232–234
Parchi P, Castellani R, Capellari S, Ghetti B, Young K, Chen SG, Farlow M, Dickson DW, Sima AA, Trojanowski JQ, Petersen RB, Gambetti P (1996) Molecular basis of phenotypic variability in sporadic Creutzfeldt–Jakob disease. Ann Neurol 39:767–778
Parchi P, Cescatti M, Notari S, Schulz-Schaeffer WJ, Capellari S, Giese A, Zou WQ, Kretzschmar H, Ghetti B, Brown P (2010) Agent strain variation in human prion disease: insights from a molecular and pathological review of the National Institutes of Health series of experimentally transmitted disease. Brain 133:3030–3042
Parchi P, Chen SG, Brown P, Zou W, Capellari S, Budka H, Hainfellner J, Reyes PF, Golden GT, Hauw JJ, Gajdusek DC, Gambetti P (1998) Different patterns of truncated prion protein fragments correlate with distinct phenotypes in P102L Gerstmann-Straussler-Scheinker disease. Proc Natl Acad Sci USA 95:8322–8327
Parchi P, Giese A, Capellari S, Brown P, Schulz-Schaeffer W, Windl O, Zerr I, Budka H, Kopp N, Piccardo P, Poser S, Rojiani A, Streichemberger N, Julien J, Vital C, Ghetti B, Gambetti P, Kretzschmar H (1999) Classification of sporadic Creutzfeldt–Jakob disease based on molecular and phenotypic analysis of 300 subjects. Ann Neurol 46:224–233
Parchi P, Notari S, Weber P, Schimmel H, Budka H, Ferrer I, Haik S, Hauw JJ, Head MW, Ironside JW, Limido L, Rodriguez A, Strobel T, Tagliavini F, Kretzschmar HA (2009) Inter-laboratory assessment of PrPSc typing in Creutzfeldt–Jakob disease: a Western blot study within the NeuroPrion Consortium. Brain Pathol 19:384–391
Parchi P, Strammiello R, Notari S, Giese A, Langeveld JP, Ladogana A, Zerr I, Roncaroli F, Cras P, Ghetti B, Pocchiari M, Kretzschmar H, Capellari S (2009) Incidence and spectrum of sporadic Creutzfeldt–Jakob disease variants with mixed phenotype and co-occurrence of PrPSc types: an updated classification. Acta Neuropathol 118:659–671
Parchi P, Zou W, Wang W, Brown P, Capellari S, Ghetti B, Kopp N, Schulz-Schaeffer WJ, Kretzschmar HA, Head MW, Ironside JW, Gambetti P, Chen SG (2000) Genetic influence on the structural variations of the abnormal prion protein. Proc Natl Acad Sci USA 97:10168–10172
Park TS, Kleinman GM, Richardson EP (1980) Creutzfeldt–Jakob disease with extensive degeneration of white matter. Acta Neuropathol 52:239–242
Peretz D, Scott MR, Groth D, Williamson RA, Burton DR, Cohen FE, Prusiner SB (2001) Strain-specified relative conformational stability of the scrapie prion protein. Protein Sci 10:854–863
Petersen RB, Parchi P, Richardson SL, Urig CB, Gambetti P (1996) Effect of the D178N mutation and the codon 129 polymorphism on the metabolism of the prion protein. J Biol Chem 271:12661–12668
Piccardo P, Dlouhy SR, Lievens PM, Young K, Bird TD, Nochlin D, Dickson DW, Vinters HV, Zimmerman TR, Mackenzie IR, Kish SJ, Ang LC, De Carli C, Pocchiari M, Brown P, Gibbs CJ Jr, Gajdusek DC, Bugiani O, Ironside J, Tagliavini F, Ghetti B (1998) Phenotypic variability of Gerstmann-Straussler-Scheinker disease is associated with prion protein heterogeneity. J Neuropathol Exp Neurol 57:979–988
Piro JR, Harris BT, Nishina K, Soto C, Morales R, Rees JR, Supattapone S (2009) Prion protein glycosylation is not required for strain-specific neurotropism. J Virol 83:5321–5328
Polymenidou M, Stoeck K, Glatzel M, Vey M, Bellon A, Aguzzi A (2005) Coexistence of multiple PrPSc types in individuals with Creutzfeldt–Jakob disease. Lancet Neurol 4:805–814
Prusiner SB (1998) Prions. Proc Natl Acad Sci USA 95:13363–13383
Puoti G, Giaccone G, Rossi G, Canciani B, Bugiani O, Tagliavini F (1999) Sporadic Creutzfeldt–Jakob disease: co-occurrence of different types of PrP(Sc) in the same brain. Neurology 53:2173–2176
Riek R, Hornemann S, Wider G, Billeter M, Glockshuber R, Wuthrich K (1996) NMR structure of the mouse prion protein domain PrP(121-321). Nature 382:180–182
Safar J, Wille H, Itri V, Groth D, Serban H, Torchia M, Cohen FE, Prusiner SB (1998) Eight prion strains have PrP(Sc) molecules with different conformations. Nat Med 4:1157–1165
Salvatore M, Genuardi M, Petraroli R, Masullo C, D’Alessandro M, Pocchiari M (1994) Polymorphisms of the prion protein gene in Italian patients with Creutzfeldt–Jakob disease. Hum Genet 94:375–379
Sanchez-Juan P, Green A, Ladogana A, Cuadrado-Corrales N, Saanchez-Valle R, Mitrovaa E, Stoeck K, Sklaviadis T, Kulczycki J, Hess K, Bodemer M, Slivarichova D, Saiz A, Calero M, Ingrosso L, Knight R, Janssens AC, van Duijn CM, Zerr I (2006) CSF tests in the differential diagnosis of Creutzfeldt–Jakob disease. Neurology 67:637–643
Schoch G, Seeger H, Bogousslavsky J, Tolnay M, Janzer RC, Aguzzi A, Glatzel M (2006) Analysis of prion strains by PrPSc profiling in sporadic Creutzfeldt–Jakob disease. PLoS Med 3:e14
Schoene WC, Masters CL, Gibbs CJ Jr, Gajdusek DC, Tyler HR, Moore FD, Dammin GJ (1981) Transmissible spongiform encephalopathy (Creutzfeldt–Jakob disease). Atypical clinical and pathological findings. Arch Neurol 38:473–477
Schulman S (1957) Bilateral symmetrical degeneration of the thalamus: a clinicopathological study. J Neuropathol Exp Neurol 16:446–470
Schulz-Schaeffer WJ, Giese A, Windl O, Kretzschmar HA (1996) Polymorphism at codon 129 of the prion protein gene determines cerebellar pathology in Creutzfeldt–Jakob disease. Clin Neuropathol 15:353–357
Schulz-Schaeffer WJ, Tschoke S, Kranefuss N, Drose W, Hause-Reitner D, Giese A, Groschup MH, Kretzschmar HA (2000) The paraffin-embedded tissue blot detects PrP(Sc) early in the incubation time in prion diseases. Am J Pathol 156:51–56
Shibuya S, Higuchi J, Shin RW, Tateishi J, Kitamoto T (1998) Protective prion protein polymorphisms against sporadic Creutzfeldt–Jakob disease. Lancet 351:419
Somerville RA, Hamilton S, Fernie K (2005) Transmissible spongiform encephalopathy strain, PrP genotype and brain region all affect the degree of glycosylation of PrPSc. J Gen Virol 86:241–246
Stack MJ, Chaplin MJ, Clark J (2002) Differentiation of prion protein glycoforms from naturally occurring sheep scrapie, sheep-passaged scrapie strains (CH1641 and SSBP1), bovine spongiform encephalopathy (BSE) cases and Romney and Cheviot breed sheep experimentally inoculated with BSE using two monoclonal antibodies. Acta Neuropathol 104:279–286
Stahl N, Baldwin MA, Hecker R, Pan KM, Burlingame AL, Prusiner SB (1992) Glycosylinositol phospholipid anchors of the scrapie and cellular prion proteins contain sialic acid. Biochemistry 31:5043–5053
Stern K (1939) Severe dementia associated with bilateral symmetrical degeneration of the thalamus. Brain 62:157–171
Stimson E, Hope J, Chong A, Burlingame AL (1999) Site-specific characterization of the N-linked glycans of murine prion protein by high-performance liquid chromatography/electrospray mass spectrometry and exoglycosidase digestions. Biochemistry 38:4885–4895
Tagliavini F, Prelli F, Ghiso J, Bugiani O, Serban D, Prusiner SB, Farlow MR, Ghetti B, Frangione B (1991) Amyloid protein of Gerstmann-Straussler-Scheinker disease (Indiana kindred) is an 11 kDa fragment of prion protein with an N-terminal glycine at codon 58. EMBO J 10:513–519
Thackray AM, Hopkins L, Klein MA, Bujdoso R (2007) Mouse-adapted ovine scrapie prion strains are characterized by different conformers of PrPSc. J Virol 81:12119–12127
Tixador P, Herzog L, Reine F, Jaumain E, Chapuis J, Le Dur A, Laude H, Beringue V (2010) The physical relationship between infectivity and prion protein aggregates is strain-dependent. PLoS Pathog 6:e1000859
Uro-Coste E, Cassard H, Simon S, Lugan S, Bilheude JM, Perret-Liaudet A, Ironside JW, Haik S, Basset-Leobon C, Lacroux C, Peoch K, Streichenberger N, Langeveld J, Head MW, Grassi J, Hauw JJ, Schelcher F, Delisle MB, Andreoletti O (2008) Beyond PrP res type 1/type 2 dichotomy in Creutzfeldt–Jakob disease. PLoS Pathog 4:e1000029
Van Rossum A (1968) Spastic pseudosclerosis (Creutzfeldt–Jakob disease). In: Vinken PJ, Bruyn GW (eds) Handbook of clinical neurology. North Holland, Amsterdam, pp 726–760
Wadsworth JD, Hill AF, Joiner S, Jackson GS, Clarke AR, Collinge J (1999) Strain-specific prion-protein conformation determined by metal ions. Nat Cell Biol 1:55–59
Will RG, Matthews WB (1984) A retrospective study of Creutzfeldt–Jakob disease in England and Wales 1970–79. I. Clinical features. J Neurol Neurosurg Psychiatry 47:134–140
Williams A, Lucassen PJ, Ritchie D, Bruce M (1997) PrP deposition, microglial activation, and neuronal apoptosis in murine scrapie. Exp Neurol 144:433–438
Williams AE, Lawson LJ, Perry VH, Fraser H (1994) Characterization of the microglial response in murine scrapie. Neuropathol Appl Neurobiol 20:47–55
Windl O, Dempster M, Estibeiro JP, Lathe R, de Silva R, Esmonde T, Will R, Springbett A, Campbell TA, Sidle KC, Palmer MS, Collinge J (1996) Genetic basis of Creutzfeldt–Jakob disease in the United Kingdom: a systematic analysis of predisposing mutations and allelic variation in the PRNP gene. Hum Genet 98:259–264
Wu Y, Brown WT, Robakis NK, Dobkin C, Devine-Gage E, Merz P, Wisniewski HM (1987) A PvuII RFLP detected in the human prion protein (PrP) gene. Nucleic Acids Res 15:3191
Yull HM, Ironside JW, Head MW (2009) Further characterisation of the prion protein molecular types detectable in the NIBSC Creutzfeldt–Jakob disease brain reference materials. Biologicals 37:210–215
Zanusso G, Farinazzo A, Fiorini M, Gelati M, Castagna A, Righetti PG, Rizzuto N, Monaco S (2001) pH-dependent prion protein conformation in classical Creutzfeldt–Jakob disease. J Biol Chem 276:40377–40380
Zanusso G, Ferrari S, Conte S, Mellina V, Sacchi V, Rizzuto N, Monaco S (2006) A 49-year-old man with neuropsychiatric symptoms followed by progressive cognitive decline. Brain Pathol 16:237–238
Zeidler M, Sellar RJ, Collie DA, Knight R, Stewart G, Macleod MA, Ironside JW, Cousens S, Colchester AC, Hadley DM, Will RG (2000) The pulvinar sign on magnetic resonance imaging in variant Creutzfeldt–Jakob disease. Lancet 355:1412–1418
Zou WQ, Capellari S, Parchi P, Sy MS, Gambetti P, Chen SG (2003) Identification of novel proteinase K-resistant C-terminal fragments of PrP in Creutzfeldt–Jakob disease. J Biol Chem 278:40429–40436
Zou WQ, Puoti G, Xiao X, Yuan J, Qing L, Cali I, Shimoji M, Langeveld JP, Castellani R, Notari S, Crain B, Schmidt RE, Geschwind M, Dearmond SJ, Cairns NJ, Dickson D, Honig L, Torres JM, Mastrianni J, Capellari S, Giaccone G, Belay ED, Schonberger LB, Cohen M, Perry G, Kong Q, Parchi P, Tagliavini F, Gambetti P (2010) Variably protease-sensitive prionopathy: a new sporadic disease of the prion protein. Ann Neurol 68:162–172
Acknowledgments
This study was supported by the RFO of the University of Bologna, the Gino Galletti Foundation and the Robert-Koch Institut (Berlin).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Parchi, P., Strammiello, R., Giese, A. et al. Phenotypic variability of sporadic human prion disease and its molecular basis: past, present, and future. Acta Neuropathol 121, 91–112 (2011). https://doi.org/10.1007/s00401-010-0779-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00401-010-0779-6