
Summary
Vitamin A is essential for growth and differentiation of a number of cells and tissues. Notably during pregnancy and throughout the breastfeeding period, vitamin A has an important role in the healthy development of the fetus and the newborn, with lung development and maturation being particularly important. The German Nutrition Society (DGE) recommends a 40% increase in vitamin A intake for pregnant women and a 90% increase for breastfeeding women. However, pregnant women or those considering becoming pregnant are generally advised to avoid the intake of vitamin A rich liver and liver foods, based upon unsupported scientific findings. As a result, the provitamin A carotenoid β-carotene remains their essential source of vitamin A. Basic sources of provitamin A are orange and dark green vegetables, followed by fortified beverages which represent between 20% and 40% of the daily supply. The average intake of β-carotene in Germany is about 1.5–2 mg a day. Assuming a vitamin A conversion rate for β-carotene for juices of 4:1, and fruit and vegetables between 12:1 and 26:1; the total vitamin A contribution from β-carotene intake represents 10–15% of the RDA. The American Pediatrics Association cites vitamin A as one of the most critical vitamins during pregnancy and the breastfeeding period, especially in terms of lung function and maturation. If the vitamin A supply of the mother is inadequate, her supply to the fetus will also be inadequate, as will later be her milk. These inadequacies cannot be compensated by postnatal supplementation. A clinical study in pregnant women with short birth intervals or multiple births showed that almost 1/3 of the women had plasma retinol levels below 1.4 µmol/l corresponding to a borderline deficiency. Despite the fact that vitamin A and beta-carotene rich food is generally available, risk groups for low vitamin A supply exist in the western world. It is therefore highly critical to restrict the β-carotene supply from diet, particularly from sources of β-carotene with high consumer acceptance such as fortified juices (e.g. “ACE juices”) or dietary supplements (e.g. multivitamins for pregnant women). For the part of the population unable to meet vitamin A requirements according to the DACH recommendations, sufficient intake of β-carotene may be crucial to help improve and maintain adequate vitamin A status and prevention of developmental disorders. At this time it has to be urgently advised against restricting the β-carotene supply or putting warning labels on β-carotene fortified products. It is, however, highly recommended to improve the available data on nutrient intakes in Germany, especially for pregnant and breastfeeding women. For them, recommendations to be aware of potential nutrient intake inadequacies might prove useful.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
There has been a long-standing discussion to limit the use of β-carotene in fortified foods and/or food supplements at European level as well as at National level in some Member States of the European Union. Measures discussed include the setting of upper limits for the use of β-carotene and/or having warning statements on the product labels. These discussions were on the one hand driven by results of the ATBC Trial demonstrating an increased lung cancer risk in long term heavy smokers upon supplementation with high doses of β-carotene (20 mg/day) over several years, while in some countries single entity high dose β-carotene supplements were marketed, at least seasonally during spring and summer. On the other hand, in the course of harmonization of European Law on Food Supplements and Fortified Foods, the scientific advisory panels to the European Commission were asked for safety evaluations of all relevant vitamins and minerals, in order to establish upper levels of these nutrients, which can be safely used in food supplements and in fortified foods.
At the national level, discussions on safe β-carotene levels for food supplements and fortified foods were most intense in those countries in which β-carotene was popular as ingredient for any of these food categories, and thus specifically in Germany and partly also in the UK. There, the Expert Group on Vitamins and Minerals (EVM), a scientific advisory group to the UK Food Standards Agency (FSA), set an Upper Safe Level for daily consumption over a lifetime of 7 mg/day of supplemental β-carotene, for both smokers and non-smokers, consequently recommending to limit the use of β-carotene in food supplement to that dose. In Germany, the German food and food supplement industry committed in 2001 to voluntarily limit the use of β-carotene in fortified foods to 2 mg/100 g or 100 ml, and to 4.8 mg/day in food supplements—4.8 mg being equivalent to one RDA of vitamin A. Food supplements containing higher doses were to carry a statement such as ‘should not be taken by smokers’.
However, these national discussions are now part of the general discussion at the European level on setting upper limits for vitamins and minerals in food supplements, as requested by Article 5 of Directive 2002/46/EC harmonizing the legislation on food supplements in the various European Member States—a discussion which is linked to the equivalent work to be done for fortified foods in light of the recently published European Regulation (EC) No. 1925/2006 for this category of foods.
National authorities may provide input to these, as has recently been done by the German Federal Institute for Risk Assessment (BfR) in form of a risk assessment with suggestions for risk management, i.e. optional measurements to be taken [21]. For foods supplements, options are to limit the use of β-carotene to either 2 mg/day, or to 2–4 mg/day, which is the desirable intake according to the German Nutrition Society. For fortified foods, suggested options are either not to allow β-carotene for food fortification at all, or limit the use to 2 mg/100 g or 100 ml. For both food categories, the BfR favours the option of 2 mg/day in supplements and no fortification, because of safety concerns with the second options, which, according to the BfR, would necessitate information statements on product labels and extensive communication of the detrimental effects of β-carotene (related to lung cancer) to consumers.
In the discussion on the safety of (high supplemental dosages) of β-carotene in smokers it is often overlooked that β-carotene is an essential micronutrient for the human being due to its function as a precursor to vitamin A, i.e. as provitamin. It is frequently claimed that the bioavailability of the ‘isolated’ or ‘purified’ β-carotene used in food supplements and fortified foods is much higher than the bioavailability of β-carotene naturally occurring in foods, However, more recent research suggests that due to conversion factors the contribution of the provitamin from foods for vitamin A supply is smaller than previously assumed.
Therefore it is important to investigate the role of β-carotene in ensuring adequate vitamin A status. On the basis of those data, risk groups of inadequate vitamin A supply due to regulatory restrictions can be identified within the population. In Addition, the consequences have to be evaluated. The following report will examine these issues using the situation in Germany as an example.
Vitamin A/β-carotene requirement and occurrence
The recommendations of the German Nutrition Society (DGE) for the daily intake of vitamin A vary for children between 0.6 mg and 1.1 mg, for adults between 0.8 mg and 1 mg, for pregnant women. For breastfeeding women 1.5 mg are recommended [27]. Figure 1 provides an example of the amount of food required to achieve the daily intake of 0.9 mg of vitamin A recommended for women.

Different foods each containing the recommended daily supply of 0.9 mg of vitamin A
Figure 1 clearly demonstrates that the only relevant dietary source for vitamin A is liver. All other foods containing preformed vitamin A need to be consumed in atypically large amounts in order to meet the vitamin A requirement. Only 10–15 g of animal liver is necessary to meet the daily requirement, demonstrating that liver is the most important source of vitamin A for humans. Since vitamin A is effectively stored in humans, just one portion of liver (100 g) every 14 days is enough to replenish these stores. Other foods like meat, butter, eggs and milk only contribute a small portion (<20%) of dietary preformed vitamin A. Liver consumption, however, can essentially be discounted as a significant vitamin A source in Germany since only 500 g of liver are consumed per person each year (41 kg of pork, 10 kg of beef) which is far less than the recommended 100 g liver every two weeks to reach a sufficient vitamin A supply. This inadequacy is especially apparent for young women who are often vegetarians and/or on low caloric diets.
Provitamin A (β-carotene) found in plant foods plays an important role for meeting vitamin A requirements from a diet that excludes the consumption of meat, liver, milk and eggs. According to the National Consumption Study and the DACH-reference values, men obtain 25% and women 30% of their vitamin A intake from the provitamin β-carotene [27, 80].
The German Nutrition Society recommends an intake of 2–4 mg β-carotene per day [27]. Figure 2 illustrates the amount of foods required to meet the recommendations, based on a conversion factor of β-carotene to vitamin A of 6:1.

The amounts of various foods containing the recommended daily supply of 2 mg of β-carotene
When assessing β-carotene sources, absorption of the provitamin β-carotene is strongly dependent on the source and the manner it is served. For example, the absorption from raw carrots is virtually non-existent whereas absorption from carrot juice is up to 60%. For diets lacking fat, the absorption is also markedly reduced.
Average intake of vitamin A and β-carotene
Mean daily intake data of total vitamin A and carotenoids is available from the National Consumption Study (1985–1988) on food and nutrient intake in Germany [107]. In Table 1 average intake data for vitamin A are listed for various age groups. Daily vitamin A intakes is notably insufficient in children and adolescents, in particular when recommended intakes of β-carotene are not being reached.
Similar results were obtained in the DONALD study, which examined children aged between 2 and 15 years [118]. Boys aged between 13 and 15 years showed the highest mean intakes of 0.88 mg of vitamin A per day. However, fortified foods increasingly contributed to the vitamin A intakes. During the 1996–2000 study period, intake of vitamin A from fortified food increased significantly whereas total vitamin A intake from all sources remained unchanged. Thus, vitamin A intake from conventional foods, that is foods without fortification, decreased to only 62% of total vitamin A intake in 2000 (Fig. 3).

Suboptimal supply of carotenoids has clearly been demonstrated in several other studies. Table 2 describes the results of the National Consumption Study on carotenoid intake, evaluated by Pelz et al. (1998), and gives data from the VERA Study, published by Schneider et al. (1995), as well as from two other studies by Müller (1996) and Riedl et al. (1997) [78, 97, 104, 107]. Since carotenoid intake in the general population is not normally distributed and arithmetic means are thus not meaningful, the median intake has to be used for consideration. Medians are usually appreciably lower than the arithmetic means. In Table 2 both medians and arithmetic means are shown, where available.
According to Müller (1996), the β-carotene intake of almost half of the population is less than 1 mg per day while 64.1% have intakes less than the recommended intake of 2 mg per day (Fig. 4) [78].

Frequeny distribution of daily intake of β-carotene [78]
Therefore the β-carotene intake of 75% of the population is less than 3 mg per day including food supplements and fortified foods. Considering that qualified and scientifically justified requirements exist to increase the β-carotene supply via foods (including fortified ones), ways of increasing the availability of foods fortified with β-carotene (possibly with good bioavailability) should be examined. In addition to the important role of β-carotene as an antioxidant it should be remembered that the provitamin is an important source of vitamin A. Vitamin A intake can be considered critical in certain population groups, especially adolescents and young women, for the reasons discussed above.
Pregnancy and breastfeeding
The need for vitamin A and β-carotene is increased during pregnancy and the breastfeeding period. On average, intakes should be one third higher during pregnancy, and during the breastfeeding period intake should be 0.7 mg/day higher than that for non-pregnant or non-breastfeeding women. Due to its importance for fetal lung development and maturation, adequate vitamin A intake is especially important during the second and third trimesters of pregnancy. The source of vitamin A, which shows the highest bioavailability is liver. However, depending on animal feeding practices, liver may contain very high concentrations of retinol (vitamin A). This has led the Federal Institute for Consumer Protection and Veterinary Medicine to advise pregnant women to avoid consuming liver [14]. Most often this results in an insufficient supply of vitamin A in pregnant and breastfeeding women who are therefore reliant on β-carotene as a source of vitamin A.
Therefore a diet rich in including fortified food and beverages has to be recommended to women at child-bearing age; especially to women with multiple births, short birth intervals, low social economic status and to those who are breastfeeding women. Intake of dietary β-carotene supplements should be recommended as well since the bioavailability of isolated β-carotene, which is used in fortified foods and dietary supplements, is better than from food matrices such as vegetables.
Vitamin A/β-carotene intake of pregnant and breastfeeding women: identification of a risk group
Doyle et al. (2001) examined the diet of women who gave birth to children with low birth weight [36]. The group with an adequate diet, compared to the group with a diet not providing adequate amounts of nutrients, had 20% lower vitamin A intakes. Most likely, the actual vitamin A intake was even lower because a conversion factor of 1:6 was used, which overestimates the contribution of β-carotene to vitamin A supply, as further discussed below. In addition, the total energy intake of the inadequately nourished group was only 1633 kcal/d, i.e. below 1800 kcal/d, which are considered borderline for a diet providing adequate levels of micronutrients. Doyle’s study (2001) clearly shows that young, pregnant and breastfeeding women have to be considered as a group at high risk for low intakes of micronutrients since those are largely provided by animal based foods. These groups should be advised to consume foods fortified with β-carotene or, even better, with vitamin A plus β-carotene, in order to avoid nutrition deficits, which should not exist in industrialized countries.
We conducted a clinical pilot study in pregnant women with multiple births or short birth intervals to evaluate the vitamin A and β-carotene supply during pregnancy and after delivery in this vulnerable population group [108]. Twenty-nine volunteers aged between 21 and 36 years were evaluated for 48 h after delivery. During this time frame a food frequency protocol considering 3 months retrospective was obtained from all participants. In order to establish overall supply retinol and β-carotene levels were determined in maternal plasma, cord blood and colostrum via HPLC analysis. Regardless of the high to moderate socio-economic background, 27.6% of participants showed plasma retinol levels below 1.4 µmol/l. This value is considered as borderline deficiency. In addition, 46.4% showed retinol intake <66% of RDA and 50.0% did not consume liver at all although liver contributes as a main source for preformed retinol. Despite high total carotenoid intake of 6.9 +/− 3.6 mg/d, 20.7% of mothers showed plasma levels <0.5 µmol/l β-carotene. Retinol and β-carotene levels were highly significantly correlated between maternal plasma versus cord blood and colostrum. In addition, significantly lower levels were found in cord blood: 31.2 +/− 13.0% of retinol and 4.1 +/− 1.4% of beta-carotene compared with maternal plasma. We conclude that despite the fact that vitamin A and β-carotene rich food is generally available, risk groups for low vitamin A supply may exist in Germany and that this may be representative for the western world.
Relevance of the mother’s vitamin A/β-carotene intake for the fetal vitamin A status
Several epidemiological studies show that an insufficient intake of vitamin A poses a risk for fetal development and also during the newborn period. As the child is dependent on the mother in terms of its vitamin A supply during the newborn period, expectant mother’s vitamin A intake during pregnancy is of critical importance for the later supply to the child. In addition liver stores of the baby only last for a couple of days and will be depleted quickly upon sudden strains or malabsorption states.
When assessing the liver stores in relation to the birth weight of the child and the intakes of the mother, Shah et al. (1987) observed a significant difference between intake groups: Insufficient Vitamin A intake resulted in low liver stores, low birth weights and, as discussed above, a higher risk of further complications [110] (Table 3).
Foetal liver stores increase with increasing gestational age, but strongly depend on the vitamin A status of the mother. Supplementation in the second trimester of pregnancy with physiological doses of vitamin A may lead to an improvement in fetal vitamin A liver stores. In addition, supplementation may also increase retinol levels in the milk, which positively affects the baby during the postnatal period.
Figure 5 illustrates that there is a direct correlation between the levels of vitamin A in the mother’s plasma and the retinol concentration in the umbilical cord. Research is urgently required to validate the importance of sufficient vitamin A intakes during pregnancy and also during the breastfeeding period for ensuring a sufficient supply to the baby.

Vitamin A concentration of the umbilical cord compared to the vitamin A concentration of the mother [117]
Relevance of breastfeeding for the vitamin A/β-carotene supply of the newborn
Since the fetal liver is only able to store a small amount of vitamin A during pregnancy, almost all babies are born with marginal vitamin A deficiency [12, 23, 130]. This is usually corrected quickly via the vitamin A supply from the mother’s milk and the extremely high vitamin A concentrations of the colostrum (up to 7 µmol/l) [130]. However during lactation maternal intake of vitamin A and β-carotene strongly affects the amount of these micronutrients secreted into breast milk [4]. The average American newborn has a mean liver vitamin A content of 5 µmol (assuming that the liver is about 4% of the body’s weight). By comparison, the breastfed infant obtains approximately 310 µmol vitamin A from mother’s milk in the first 6 months. Thus, during the first 6 months, vitamin A intake from breastfeeding is usually 60 times higher than the intake that can be attained during the 9 months’ pregnancy [102].
Vitamin A stores in the fetal liver accumulate during the last trimester of pregnancy but stores are related to maternal plasma concentrations [83, 84]. In cases of zinc and vitamin A deficiency during pregnancy, daily supplementation with β-carotene (4.5 mg) and zinc (30 mg) improved vitamin A status in expectant mothers and in their newborns [33]. Breast milk concentrations of retinol and β-carotene were higher at 6 months of lactation after supplementation with β-carotene. The authors calculated the median daily retinol intake from breast milk would be 216 RE (Retinol Equivalents) in the supplemented and 148 RE in the control group. The UK estimated average requirement and lower reference nutrient intake for retinol in this age group are 250 RE and 150 RE respectively [33].
The World Health Organization recommends a daily minimum intake of 0.63 µmol/l retinol via breast milk for babies in order to meet basic requirements [8]. For the development of liver stores, however, 1.2–1.3 µmol of retinol per day during the first year and 1.4 µmol/day during the second and third year are necessary [38]. Assuming an average milk intake of about 750 ml/day by the baby, the mother’s milk needs to contain 1.6 µmol retinol per liter in order to provide the recommended intake requirements of the breastfed baby. According to data available for Germany, the mean retinol concentration of mother’s milk is about 2.8 µmol/l, but was less than 1.6 µmol/l in more than 20% of the women, i.e. concentrations, which are considered critically low for the babies supply. Insufficient vitamin A intake of the baby may have serious consequences, especially regarding susceptibility to infections, the development and function of breathing organs, and the integrity of mucous membranes.
In a recent randomized controlled trial 150 women were supplemented with a single dose of retinol (209 µmol/l) soon after delivery and were advised to breastfed for 6 months [11]. Pre-supplementation mean serum retinol was 0.98 µmol and breast milk retinol 3.85 µmol. Serum and breast milk retinol increased immediately after supplementation compared to the control group, and breast milk retinol remained significantly higher for 4 months compared to the control group (Fig. 6). Further, in the supplemented group a decreased incidence and duration of various diseases was observed.

Frequency distribution of vitamin A concentrations in mother’s milk
Vitamin A and healthy development of the fetus and newborn
Vitamin A is essentially required for healthy development of the fetus and the newborn. A number of intervention studies indicate that the number of neural tube defects in children born to women taking multivitamin supplements compared to women taking no supplements was significantly lower. In addition to folic acid, vitamin A supplementation (4000–6000 IU) was mentioned as important factor for decreasing such defects [60]. Further, insufficient micronutrient intakes by pregnant woman influence the postpartum intake of the child. Several studies have shown an increased risk of bronchopulmonary dysplasia (BPD) in preterm infants with insufficient vitamin A status [7, 25, 61, 132].
Effects of vitamin A on gene expression during fetal lung development
The vitamin A metabolite retinoic acid (RA) plays an important role in modulating gene expression during fetal lung development. RA can inhibit the expression of the surfactant protein A (SPA), a component of the surfactant synthesized and secreted in type II alveolar cells, in a concentration-dependent manner [73, 143]. Equally insulin, TGF-β and high concentrations of glucocorticoids can lead to an inhibition of SPA mRNA expression [119, 135]. Lower glucocorticoid concentrations stimulate the expression of this gene [89]. The SPA mRNA expression is increased in human fetal lung explants via RA as well as via hyperoxia (data from rats) and dexamethasone (human fetal lung explants) [73, 87, 89, 119, 135]. Thus, the synthesis of the individual surfactant proteins is regulated selectively and differently via RA together with glucocorticoids.
Prostaglandins of the PGE2 type can increase the surfactant protein synthesis [59, 67]. Under the influence of EGF (epidermal growth factor), the formation of prostaglandins and, in particular of PGE2, increases [65, 126]. The expression of the EGF receptor is increased by RA. EGF increases the proliferation of the lung tissues and leads to an increased formation of surfactant phospholipids [44, 49, 124]. RA as well as EGF increase (40%, 80%) the PGE2 secretion in fetal lung cells of the rat in vitro by 40% and 80%, respectively, while the combination of RA and EGF increases PGE2 secretion more than 6-fold [88]. Therefore RA influences lung development through its modulating effect on EGF expression and the resulting PGE2 induced surfactant synthesis. Important for the time-dependent regulation of lung development are a sufficient continuous supply of retinol (from blood or tissue stores) and a timely formation of the active metabolite RA.
Vitamin A kinetics during fetal lung development
Local i.e. extrahepatic stores, of retinyl esters were demonstrated in fibroblast-like cells located close to alveolar cells, in type II cells as well as in the respiratory epithelium [15, 90, 144]. The importance of these retinyl esters as “acute reserve” during lung development was confirmed in rats: during late phases of pregnancy, when lung maturation starts, these retinyl ester stores are quickly depleted [42]. Such depletion is the result of an increased requirement during lung development, brought on by the ‘acute’ need for retinoic acid for necessary cellular differentiation (proximalization) and metabolic purposes (surfactant).
Prenatal lung development is also influenced by glucocorticoids, both, in parallel and complementary to vitamin A. This is not surprising since the receptor for steroids and for retinoids belongs to the same multi-receptor family. The action of the glucocorticoids, however, does not only begin at the level of gene expression but much earlier in that it seems to regulate the release of the vitamin. For example, the administration of dexamethasone leads to increased levels of the maternal and fetal retinol binding protein resulting in an improvement in vitamin A supply via the normal route, i.e. activation of liver stores. Such increases in vitamin A concentrations in the systemic circulation however decrease the morbidity and the mortality of preterm infants due to bronchopulmonary dysplasia [113, 115]. Dexamethasone and glucocorticoids not only lead to an improved total vitamin A status by mobilization of liver stores but also influence the metabolization of the vitamin A esters stored in the lung [42]. Following administration of dexamethasone, but also without application of steroids, the levels of retinyl esters were noticeably reduced. This may explain the therapeutic successes of steroids but also their failures in the treatment of lung distress syndrome in preterm infants. If there are insufficient retinyl ester stores due to insufficient supply to the fetal lung during the late phases of pregnancy, the regulating effect of glucocorticoids on the vitamin A metabolism of the lung cell cannot take place.
Low vitamin A plasma levels are relatively often observed in preterm infants, particularly in those with lung distress syndrome due to the relatively undeveloped capacity of the liver to synthesize retinol-binding protein [111]. The newborn is almost entirely dependent on its mother for vitamin A supply, i.e., the retinyl ester stores in its lung will only be replenished–either by direct intake or via synthesis from circulating retinal–if the mother has had a sufficient vitamin A supply during pregnancy.
Effects of insufficient vitamin A intake on the postnatal development of the lung
Preterm infants and particularly newborns are dependent on a sufficient supply of vitamin A to ensure regulation of cellular differentiation in the respiratory and lung epithelia. The more premature the child, the lower its serum retinol levels are [77]. The plasma levels are critical at birth in terms of lung development because serum retinol and RBP levels decrease further following birth.
Several studies consistently show that serum retinol and RBP levels are significantly lower in preterm infants than in newborns [48, 52, 109]. Compared to newborns there are also very low retinol levels in the liver of preterm infants [116]. Plasma values lower than 20 µg/dl are common and are indicative of a relative vitamin A deficiency.
Low vitamin A plasma levels in the first months after birth significantly affect the development of the baby in general and its susceptibility to infection. Recurrent infections have often been observed when retinol plasma levels were low [10, 41, 98]. Such infections rank among the main complications of vitamin A deficiencies, which occur in developing countries, for example. Moreover serum vitamin A levels decrease further during infections, particularly of the respiratory tract, because of increased metabolic needs as well as increased renal excretion of retinol and RBP during acute infections [82, 121].
Bronchopulmonary Dysplasia in preterm infants
Bronchopulmonary dysplasia (BPD) is a chronic disease of the lung that develops during the neonatal period (mainly, but not exclusively) in preterm infants. Potential causes are artificial respiration, barotrauma, respiratory infections, excessive liquid supply, as well as an insufficient vitamin A status of the preterm infant. BPD is a syndrome defined by three characteristics: oxygen dependency, radiographic abnormalities and chronic restriction of lung function in preterm infants for a time period longer than 28 days. Today, BPD has become rare in children with birth weights of more than 1500 g, possibly due to advances in respiration technology and intensive care. Since increasing numbers of preterm infants with very low birth weight survive today, the prevalence of BPD has increased again, and it is the most common chronic lung disease in newborns. For example, in Tennessee, USA, the incidence of BPD is seven times higher than that of cystic fibrosis, and 15 times higher than that of chronic interstitial lung disease or congenital malformations [50]. As BPD is a multifactorial disease, its therapy is also complex, and there are no standard recommendations for therapy.
One main cause of BPD may be the vitamin A status. Some morphological changes are strongly reminiscent of changes observed in vitamin A deficiency in humans and animals. Specifically, these include the focal loss of ciliary cells with keratinized metaplasia, necrosis of the bronchial mucosa as well as the increase in mucous secreting cells [120, 122]. The effect of vitamin A deficiency in particular on focal keratinized metaplasias suggests a disturbance in differentiation at the level of gene expression.
Preterm infants at birth have much lower concentrations of vitamin A, vitamin A (retinol) binding protein (RBP), and of hepatic vitamin A stores compared to full-term infants [20, 56, 76, 81, 111]. Such relative vitamin A deficiency in preterm infants is made even more complex because of further decreases in the vitamin A blood levels, which occur during intensive care [47, 112, 141]. Furthermore, preterm infants develop biochemical signs of vitamin A deficiency at the time point of term delivery [92, 96, 140].
In addition to postnatal treatment (enteral or parenteral vitamin A), prenatal vitamin A supply of the developing child is the most important form of treatment. Postnatal treatment of BPD with vitamin A does not lead to uniform results (see chapter 6: Vitamin A therapy of newborns). However a sufficient prenatal supply of the expectant mother will improve vitamin A status of the preterm infant.
Vitamin A therapy of newborns
Babies with extremely low birth weights are at obvious risk of vitamin A deficiencies as they show low retinol concentrations in plasma during or just after birth with mean values indicative of marginal [<0.7 µmol/l (200 µg/l)] or manifest vitamin A deficiency [<0.35 µmol/l (100 µg/l)] [47, 62]. According to the recommendations of the American Academy of Pediatrics [145], vitamin A intakes of preterm infants should be 210–450 µg retinol per kg body weight per day. However, further data are necessary to establish if these intake levels shall also apply to babies with extremely low birth weight. Nevertheless, all children with birth weights lower than 1500 g who were given 120 µg of retinol per 100 kcal or about 122 µg per kg body weight for a month still had hyporetinolaemia [62]. This was particularly evident in children who were given formula milk, mother’s milk providing only 100 µg of vitamin A per kg per day, or parenteral nutrition over a longer period of time without additional vitamin A supplementation.
The role of vitamin A in epithelial differentiation has led to studies investigating the effect of high-dose vitamin A supplementation in children with very low birth weight, with special regards to the re-epithelization of the lung tissue after acute damage caused by barotrauma or hyper oxygenation.
Common goal of these studies was to prevent the development of BPD. They were evaluated according to the criteria of the Cochrane Reviews by Darlow and Graham (1999) [28]. Overall, the studies examined the effect of vitamin A supplementation on vitamin A plasma levels as well as on decreasing mortality and morbidity in children with a birth weight between 700 g and 1500 g. Morbidity was defined as chronic lung disease, BPD and retinopathy of the newborn. Five out of ten studies met the selection criteria of the Cochrane Reviews. All studies were randomized or “quasi-randomized” and tested a high dose of vitamin A (intramuscular or oral administration) against placebo or no treatment.
The meta-analysis included 149 children who were given vitamin A, and 141 untreated children. Four of the studies used water-soluble retinyl palmitate (600–1200 µg vitamin A every second day or three times a week) given intramuscularly for 28 days [13, 28, 93, 95]. In the fifth study approximately 750 µg (2500 IU) were administered daily intravenously using a lipid emulsion [136]. During the examination period, no or nearly no prenatal and postnatal steroids and surfactant proteins were given.
One result of the meta-analysis of Darlow and Graham (1999) was that high-dose vitamin A did not influence the mortality of one-month-old children but showed a clear tendency for decreased dependency on oxygen therapy [28]. There was a significant effect of high-dose vitamin A when mortality and oxygen therapy were evaluated together. Further, retinopathy in preterm infants tended to be lower.
In a multicenter study published after the meta-analysis, 807 children with a birth weight lower than 1 kg were given 1500 µg (5000 IU) of vitamin A intramuscularly in the form of retinyl palmitate for three times per week for 4 weeks [128]. There was a small but significant decrease (62% compared to 55% for non-treated controls) of the mortality rate or in the incidence of chronic lung diseases at the age of 36 weeks. However, the treatment did not achieve plasma retinol levels above the level indicative of biochemical deficiency (0.7 µmol/l) in all children.
The multicenter study showed that a 1500 µg (5000 IU) dose of vitamin A administered three times a week is necessary to maintain normal biochemical vitamin A status [128]. An expert panel of the Life Sciences Research Office (LSRO) of the American Society for Nutritional Sciences (ASNS) has recommended, however that preterm infants be given at least 204 µg of RE (retinol equivalents)/100 kcal (680 IU/100 kcal) and at most 380 µg of RE (1267 IU)/100 kcal for preterm infants [60].
In conclusion, high-dose vitamin A seems to play a role in the prevention of BPD. Further studies are needed to support the effects of high-dose vitamin A applied in conjunction with steroid therapy, as is commonly done today in clinical practice. When given to children with extremely low birth weights, steroids and in particular dexamethasone, induce temporary increases in plasma retinol and RBP presumably by stimulating their release from the liver and from stores in the lungs [43, 114]. The results of the meta-analysis demonstrate that compensating for vitamin A deficiencies of preterm infants and newborns is only partially successful. Much more important is ensuring sufficient vitamin A stores of the child. This occurs mainly during the last 90 days of pregnancy and can only be ensured by a diet supplying considerable amounts of β-carotene-rich fruit and vegetables or preformed vitamin A (for example 50–100 g of liver/week).
Weekly supplementation of women in Nepal with vitamin A (7 mg) or β-carotene (42 mg) reduced mortality during pregnancy by 40% and 49% [138]. The apparent reason for reduced mortality risk was less susceptibility to infection.
General considerations on β-carotene as a source of vitamin A
Bioavailability and bioequivalence
Bioavailability is defined as the fraction of a substance (e.g. carotenoids), which is available for normal physiological functions or for storage. Bioconversion refers to the amount of a carotenoid converted to retinol in the body.
Carotenoids are lipophilic substances absorbed from the small intestine along with other lipids and reappearing in the lipoprotein fractions of the plasma, as well as in erythrocytes and leucocytes [68, 71, 94]. The largest portion of the carotenoids in lipoproteins is found in LDL (75–80%) followed by HDL with 10–25% and in VLDL with 5–10% [72]. Increases in plasma concentration after carotenoid intake are subject to large inter-individual variation [34]. Typical serum levels of the most common carotenoid β-carotene range from 0.2–0.5 µmol/l [16]. Non-absorbed β-carotene is exclusively found in the feces while apocarotenals and not identified fragments have also been found in urine [46, 69].
Factors determining bioavailability and bioconversion are grouped under the mnemonic “SLAMENGHI” (Species of carotenoid, molecular Linkage, Amount of carotenoids consumed in a meal, Matrix in which the carotenoid is incorporated, Effectors of absorption and bioconversion, Nutrient status of the host, Genetic factors, Host-related factors and mathematical Interactions) [137]. The type and amount of carotenoids in plasma reflect those contained in the diet.
The bioavailability of carotenoids is influenced by the form in which they are present in the food matrix, especially if they are in crystalline form, esterified and/or emulsified in fat. Depending on the fat content of the meal, absorption from plant foods ranges between 30% and 60% and is subject to large inter-individual variation [35, 101]. The following Fig. 7 summarizes these effects using a number of examples.

Influences of the matrix on the bioavailability of beta-carotene [32]
Intake of β-carotene from supplements results in a larger plasma level increase compared to intakes of corresponding amounts from vegetables. Studies conducted by Micozzi et al. (1992) demonstrated that supplementation with a low dose of β-carotene (12 mg per day) leads to significantly greater increases in plasma concentrations (from 0.3 to 3.9 µmol/l) than can achieved by intake from foods [75]. These results were corroborated by findings from Le Marchand et al. (1994), who showed that doubling fruit and vegetable intake only increased the β-carotene plasma level by 40% [64].
In addition to fat, a number of other nutrients can impact carotenoids absorption, metabolism and bioconversion if consumed in conjunction with carotenoids. Proteins help to stabilize the fat emulsion in the small intestine and along with lecithin support micelle formation. The stomach pH can also considerably influence the absorption of carotenoids: After administrating a single dose of 120 mg β-carotene, its absorption at a pH value of 6.4 was only half as much at normal stomach pH (at about 2) [127]. Genetic factors that influence carotenoid absorption might be linked to fat absorption. Dividing a daily carotenoid dose into three single doses can improve absorption, as well as the presence of bile acids [91, 100].
The greatest absorption rate is achieved by carotenoids dissolved in oil, so that 3.3 µg of β-carotene can be converted into 1 µg of retinol [53]. Bioavailability of carotenoids from most other foods is much lower than from those prepared with oil. Therefore in 1967, “6” was established as the mean conversion factor for calculating retinol equivalents (RE) from carotenoids [37]. However, more recent studies showed that the bioavailability of β-carotene from vegetables is much lower than previously assumed, and that the so far accepted conversion factor most probably overestimates the contribution of carotenoids to the dietary vitamin A supply [29, 30]. De Pee (1995, 1998) now recommends a conversion factor of 12 for β-carotene from fruits and 26 for β-carotene from vegetables, instead of the current 6 [29, 30]. The following table shows the various conversion factors for different foods and the corresponding literature sources.
When evaluating studies that examined the bioavailability from foods versus dietary supplements, it is obvious how much bioavailability can vary, and how important β-carotene sources with high bioavailability and thus bioconversion, such as food supplements, can be to ensure sufficient vitamin A intakes.
Table 5 lists the most important studies regarding bioavailability. As shown in the table, subjects received either food supplements or a specific diet for a period of three to nine weeks. Following the supplementation period, plasma β-carotene levels were determined. The bioavailability and bioconversion of a given food and thus a food group can be estimated from the increases in plasma β-carotene concentrations upon supplementation or the specific diet. Thus, a conversion factor may be established to determine the amount of β-carotene required from a specific food source in order to provide 1 µg of RE. A pre-requisite for deriving this conversion factor from dietary supplementation studies, however, is that the bioefficacy of β-carotene dissolved in oil needs is known.
The efficacy of purified β-carotene dispersed in oil was first established in 1949 in a study involving two subjects with overt vitamin A deficiency [51]. After 18- and 22-month depletion the subjects were given β-carotene in oil for six months and five weeks. The impaired vision (reduced dark adaptation) that had occurred due to depletion was reversed by administering a dose of 2500 IU of β-carotene in oil or 1300 IU of vitamin A. Therefore an effect ratio of 2:1 was defined.
The basis of these data from 1949, however, is now regarded as highly questionable: Today, it is known that the correction of dark adaptation via β-carotene occurs through via the action of the enzyme 15, 15′-dioxygenase that is found in the pigment epithelium, but not, as was assumed at that time, only by formation of vitamin A from β-carotene in the intestine. Therefore, even low amounts of β-carotene may produce that effect.
To calculate the bioefficacy of individual foods in the following Table 5, the conversion factor defined by the International Union of Pure and Applied Chemistry (IUPAC) in 1959, was used: 3.3 µg of β-carotene in oil correspond to 1 µg of retinol [53].
In 2001 the Food and Nutrition Board (FNB) published a new conversion factor in the USA, in addition to new recommendations for vitamin A intakes [31]. The long used unit for vitamin A intake (Retinol Equivalent, RE) was replaced by a new unit named the retinol activity equivalent (RAE). According to this new unit, the vitamin A activity of carotenoids absorbed from foods as provitamin is now half its former value. The RAE of β-carotene was defined as 12 µg of carotenoids per 1 µg of retinol [31]. Therefore, the new values show a provitamin activity of β-carotene from fruit and vegetables, which is only half of the previous RDAs (Recommended Dietary Allowances) [103].
The application of a conversion factor of 12, instead of 6, to VERA data would result in a more than 15% reduction of the total vitamin A intakes in Germany. The percentage of people achieving just 50% of the recommended intakes would increase considerably. Previously, an average of 87% of the total vitamin A requirement was met by children and adolescents (females, 4 ≤ 19 years). Using the conversion factor of 12 instead, the requirements are met by on average less than 70% in these population subgroups.
Daily carotene intake of women was calculated as 1.64 mg a day by Schneider et al. (1995), which is only 82% of the recommended intake considering a recommended daily intake of 2 mg [107]. According to data from the VERA study, 52% of these come from fresh vegetables and about 10% from juices and non-alcoholic drinks (Fig. 8).

Percentage of different food groups of total intake of β-carotene in comparison with recommended intake in % (women with a mean intake of 1.64 mg/day) [107]
These calculations, however, are purely based on consumption data and do not account for differences in bioavailability from different foods. Taking into account differences in bioavailabilty using conversion factors as stated in Tables 4 and 5, weighing of the food groups shifts in favor of juices and non-alcoholic drinks. Figure 9 shows the intake of β-carotene in RE calculated using conversion factors proposed by De Pee et al. (1998) (fresh vegetables: 26:1, vegetarian products and fruits: 12:1 and other foods: 6:1) in comparison with a calculation using the conversion factors as recommended by the FNB; naturally occurring β-carotene: 12:1) [30, 31]. Juices and non-alcoholic drinks are considered in both cases with the factor 4:1 because β-carotene is more readily bioavailable from juices and juices and non-alcoholic drinks are frequently fortified with β-carotene.

Using the conversion factors proposed by De Pee et al. (1998) results in higher intakes in terms of RE from juices and non-alcoholic drinks compared with intakes from fresh vegetables [30]. If fortification of juices and non-alcoholic drinks with β-carotene is reduced, or consumption of such products is decreased, consumers lose an important source of vitamin A, and certain risk groups, will have insufficient intakes.
Even when using the conversion factor of the FNB (12), beverages remain an important source of β-carotene that should not to be ignored.
Plasma and tissue concentrations in the German population
Mean β-carotene plasma levels in Germany are 0.3–0.6 µmol/l. Studies from other countries indicate differences in β-carotene levels of 20–100% between men and women [5, 6, 19, 26, 55, 85]. Additional factors that can influence carotenoid status are age, disease and/or diet. Even short-term supplementation of physiological amounts of β-carotene results in considerable changes in plasma concentrations. As Johnson et al. (1995) observed a short-term increase of 127% (0.54 ± 0.11 µmol/l) occurred after a single β-carotene administration of 120 mg [58].
Long-term supplementation of carotenoids leads to sustained increases in serum concentration with a plateau being reached after 4 weeks on average, and plateau concentrations varying from individual to individual [57, 70, 134].
Which plasma and tissue concentrations may be related to good health, or may be critical, and the means of achieving these are the subject of numerous discussions. The Consensus Conference on antioxidants concluded that a plasma level greater than 0.4 µmol/l, as achieved from a daily intake of 2–4 mg of β-carotene, can be regarded as desirable regarding disease prevention [17]. However, based on the findings of two intervention studies (ATBC, CARET), a value for plasma levels must be determined corresponding to a limit value for a tolerable upper intake level (UL). Because the Food and Nutrition Board (FNB) did not define a tolerable upper intake level (UL) for β-carotene, the plasma levels achievable from supplementation need to be taken as the limit value to assess safety. A key question to address is whether a critical plasma level will be exceeded by a defined β-carotene dosage. According to the definition given by the Consensus Conference on Antioxidants, the “critical plasma level” is a level greater than 3 µmol/l [17]. Thus, it must be determined whether a specific amount of β-carotene consumed for a certain period of time will result in plasma levels exceeding this “critical plasma level”, taking into account individual variability. Unfortunately, there is very few scientific data to answer this question, and the data available is of very variable quality due to the large number and not standardized methods of measurement used, but it also reflects the high degree of variability of individual β-carotene plasma levels.
Inter-individual variability
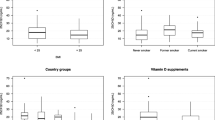
The plasma levels of β-carotene are on average higher in women than in men with differences of between 20 and 100%, depending on the study [5, 6, 19, 26, 55, 85]. Markedly reduced plasma levels of β-carotene are observed in smokers and alcoholics [1, 24, 54, 63, 66, 105, 123, 129]. Even low daily alcohol consumption (0.5 g/kg body weight/day) can lead to clear reductions in β-carotene plasma levels [125]. Further factors affecting plasma levels are age, illness and/or the type of diet. In addition to these more obvious parameters, there are others requiring more study such as responder type and interaction with other micronutrients (tocopherols, retinol, other carotenoids) [74]. These interrelationships are often unclear and even partly contradictory [3, 45].
The rise in plasma concentrations, which can be achieved through a specific dose, cannot be clearly predicted on the basis of the studies conducted during the previous 12 years (Fig. 10). The average plasma levels of healthy humans vary between 0.2 and 0.6 µmol/l, taking into account the variations discussed above. With daily dosages ranging from 12 to about 15 mg, plasma levels between 3 and 4 µmol/l are reached, often even after just a few (4–6) weeks of supplementation. With a few exceptions, markedly greater increases were observed only with significantly higher doses (100–580 mg/day). Here plasma levels of 7 to higher than 12 µmol/l were reached. Lower dosages (20 mg) can lead to higher levels (7 µmol/l) if daily supplementation is continued over a number of years [2]. Such high dosages cannot be considered as “normal” supplementation level for the general population, but may well be of importance in specific circumstances.

β-carotene plasma values from supplementation of different daily doses (the numbers in the graphic show the individual daily doses of β-carotene in mg)
Very little is known about the time-dependent course of plasma β-carotene during a dietary supplementation regimen because most studies have only determined an end value. According to Prince and Frisoli (1993) the half-life of β-carotene in plasma is independent on the daily dosage and is approximately 9–10 days. In their study, the steady state was reached after about 30 days (Fig. 11), and was depended on the daily dose [100]. Once supplementation was stopped, plasma concentrations dropped exponentially (half-life about 10 days). Comparable results had already been presented previously by Ringer et al. (1991) [106]. At daily doses of β-carotene between 15 mg and 180 mg a plateau was reached in all groups after 2–4 weeks, with the plateau concentrations depending on the given dose. However, it is unclear if and how plateau concentrations can be modified by other parameters. According to animal experiments, the activity of 15,15′-dioxygenase, the enzyme that cleaves β-carotene to form vitamin A in the intestinal mucosa, depends upon the retinol status of the animal [133]. Therefore, in the case of suppressed dioxygenase activity (for example during simultaneous intake of vitamin A), the bioavailability of β-carotene may be increased.

Temporary course of the plasma concentrations during a supplementation of two different daily doses (source [100])
Despite of the importance to answer such questions, interactions with other nutrients in humans have only been investigated in few studies. The results were often contradictory, for example with regard to vitamin E. While some researchers observed a decrease in vitamin E plasma levels upon β-carotene supplementation others found no such decrease or even observed a slight increase [3, 18, 45, 79, 86, 139, 142]. Provided that there is no deficiency retinol plasma levels are not affected by β-carotene supplementation as has been reported consistently [3, 99, 139]. Synergistic effects of simultaneous administration of β-carotene and vitamin A have not been investigated yet. However, data from animals suggest that such an interaction may occur, modulated via regulation of the 15, 15′-dioxygenase activity.
In conclusion, β-carotene plasma levels are subject to strong inter-individual variations, which are partly due to factors such as gender, age, diet, nicotine and alcohol consumption. Supplementation of β-carotene leads to a marked increase in plasma levels, on average by factor 10. After about 4 weeks, a plateau is reached. The plateau level achieved is not only dependent on the daily dose but also on the food (or supplement) matrix, daily dose frequency, simultaneous intake of fat, as well as the baseline levels of the individual. Once supplementation is discontinued, plasma concentrations return to baseline levels within a few weeks. It is not yet well understood if and to what extent interactions with other micronutrients affect the absorption of β-carotene.
Conclusions
β-carotene is a substance widely distributed in nature and found in foods of plant origin. β-carotene concentrations in these foods are, however, subject to considerable variation. β-Carotene is an important nutrient due to the many functions in the human body, depending on its metabolization: β-carotene acts as antioxidant, by quenching singlet oxygen and scavenging peroxyl radicals. Further, the apocarotenals obtained from eccentric cleavage may exert their own effects, as is currently addressed in many studies.
Long before the importance of β-carotene as antioxidant was established, β-carotene was known as provitamin A and thus as a precursor for an essential nutrient. This function became a “side issue” during the period of high interest in the antioxidant effects of β-carotene, and was re-considered recently when studies re-examined if and how much β-carotene is needed to meet the vitamin A requirements of humans by β-carotene alone. This was especially important for regions of the world where the provitamin represented the major source of vitamin A, as diets lack preformed vitamin A. A pre-requisite for β-carotene being used as precursor for vitamin A is that the provitamin present in foods is bioavailable and converted to vitamin A. Established β-carotene conversion factors vary between 4:1 (fortified juices) and 26:1 (fresh spinach) depending on the individual food. The conversion of β-carotene dispersed in oil, the form found in food supplements, is assumed to be 3.3 while for fortified juices the conversion rate is 4:1–6:1, for fruit 12:1 and for vegetables 26:1. The Food and Nutrition Board of the USA 31 cited 12:1 as the general conversion factor for typical plant based foods, i.e., 12 mg of β-carotene needs to be consumed in order to produce 1 mg of vitamin A; the daily recommended dose. The β-carotene intake in Germany is approximately 2 mg per day, and usually lower, according to the four studies available so far. This means that β-carotene from food sources contributes at the most to 10–15% of the recommended vitamin A intakes. For calculating the vitamin A intake, which amounts to on average about 1 mg/day in Germany and thus meets the current recommendations (1 mg), the contribution of β-carotene is included in the calculation using a conversion factor of 6. This means, unfortunately, that the actual contribution of β-carotene to the vitamin A intake is overestimated, due to the poorer conversation rates of β-carotene known today. Children and adolescents, for example, hardly meet the recommended vitamin A intakes for their age groups. Even more critical, for pregnant and breastfeeding women it is nearly impossible to meet the higher intakes recommended for their special situation. If the more recent, now accepted conversion factors were applied, the vitamin A intakes of these risk groups would be even worse. Adolescents would only consume about 70% of the recommendations for vitamin A intakes, and pregnant and breastfeeding women even less, especially if they do not consume liver. NHANES III showed that 16–33% of all children examined (mainly from lower income families) aged between 4 and 8 years had critically low vitamin A blood levels [9]. This is a serious condition as retinol levels only decrease if the liver stores are almost depleted. According to the third nutrition report of the FASEB, too low vitamin A intakes have to be considered a frequent and serious health problem [39].
Considering the improved bioavailability of β-carotene from food supplements (conversion rate 3.3:1) and the fortification of beverages and other foods, specifically those with a lipid matrix, fortified foods and food supplements potentially play an important role in supplying vitamin A to the population. Studies conducted in Germany show that β-carotene accounts for up 25–30% of vitamin A intakes. Considering a conversion factor of 12:1, 30% of the total β-carotene intake is derived from fortified juices and the so-called “ACE drinks”, implying that about 10% of the total vitamin A supply comes from juices and other beverages. This is especially important for young women who are known to avoid meat and innards. A restriction of, or a warning label on such fortified foods or food supplements, cautioning against β-carotene, would cause young women, especially those considering pregnancy, to avoid those foods completely. This may create a difficult situation, as indicated already by a number of recent studies: insufficient vitamin A intakes in spite of ample possibilities to ensure adequate intakes.
There are two main causes for that: Young women and those considering pregnancy have been repeatedly advised to avoid the consumption of liver because of the claimed risk for very high vitamin A levels in liver [14]. However, a risk for too high vitamin A intake in form of preformed vitamin A exists, if at all, only during the first 4 weeks of pregnancy, but not later. Furthermore, assuming an absorption rate of 40%, it is hardly possible to consume critical amounts of vitamin A from 100 g of liver. In addition, the actual teratogenic substance is not vitamin A (retinol), but its metabolite retinoic acid, which does not occur in foods, but will only be synthesized from retinol in the body. Since the synthesis of retinoic acid from retinol is strictly controlled, even excessive retinol intakes will not result in supra-physiological levels of retinoic acid. Thus, the warning against liver consumption is not based on scientific evidence, and may have caused the low consumption of liver to decrease even further, especially among young women. The amount of liver consumed in Germany is approximately 500 g/capita/year and has probably ceased completely for young women, as confirmed by data from Austria and the UK [40].
β-carotene in plant foods as previously discussed, is only a limited vitamin A source. Nevertheless, it may be an important vitamin A source if provided in sufficient amounts. In Germany the recommendations are just about reached even though food supplements and fortified food have already been considered. Not only the health of the mother may be at risk if vitamin A intakes are insufficient, but also and even more so the development of the child. Studies from the UK demonstrated that decreasing energy intakes (<1800 kcal) results in reduced intakes of various micronutrients, particularly folic acid, iron and vitamin A, and is associated with low birth weight. The overall development of the baby, and especially lung development and maturation of the embryo, is essentially depending on a sufficient vitamin A supply. If vitamin A supply is low, vitamin A stores in the lung especially of preterm babies are low. Vitamin A stores are, however, necessary to ensure proper lung development, including expression of surfactant proteins and a number of factors for growths and differentiation. In preterm infants the vitamin A blood levels are usually too low because the liver cannot yet produce sufficient retinol binding protein in order to secrete vitamin A. Therefore it is critical to develop sufficient vitamin A stores in the lung, which happens in the third trimester of pregnancy. If not, these children will be at increased risk for bronchopulmonary dysplasia (BPD), one of the most frequent and threatening respiratory diseases in preterm infants. Postnatal treatment with vitamin A was only moderately successful so far, strongly suggesting that maternal vitamin A supply in the third trimester of pregnancy is essential for the development of sufficient vitamin A stores for proper lung development. The same applies to babies, which are breastfed and thus get their vitamin A via their mother’s milk. If the vitamin A status of the mother is low, an adequate supply to the baby cannot be ensured, which has been associated with increased sensitivity to respiratory diseases caused by the respiratory syncytial virus (RSV). A significant relation between RSV-associated lung disease and previous BPD has been demonstrated.
A further scientifically not justified restriction of vitamin A sources initiated by authorities places the safe and adequate vitamin A supply at risk. Due to better bioavailablity particularly β-carotene fortified foods contribute significantly to vitamin A intakes. Restrictions, which are largely relevant to smokers should be considered very carefully: The application of warning labels or other measures on products may avoid those who depend on such products (e.g. young women considering pregnancy) consuming them. Scientific evidence on the risks of an insufficient vitamin A status for lung maturation and development of the fetus and the baby are much more convincing and accepted by the scientific community than the evidence for developing lung cancer upon high β-carotene intakes. Furthermore, the dosages used in the studies on lung cancer can not be achieved in Germany from foods, even if fortified foods are included, even less so on a long term basis. Until the contribution of β-carotene containing food supplements and fortified foods to vitamin A intakes has been fully clarified, any restrictive limitations should be avoided. This does not mean that label indications specific for smokers might not be useful, or that higher β-carotene dosages (>5 mg per day) might be refined to licensed medicines. The main focus should however be on diet and health education measures, which communicate the risk for micronutrient deficiencies especially in young women, and offer strategies for avoiding such risks.
References
Adoncecchi L, Marrocco W, Suraci C, Pecora P, Gallinella B, Porra R, Cavina G (1984) Effect of renal and liver failure on blood levels of vitamin A, its precursor (beta-carotene) and its carrier proteins (prealbumin and retinol binding protein) Influenza dell’ insufficienza renale ed epatica sul tasso ematico della vitamina A, dei suoi precursori (beta-carotene) e dei suoi vettori proteici (PA e RBP). Boll Soc Ital Biol Sper 60:881–886
Age-Related Eye Disease Study (AREDS) Research Group (2001) A randomized, placebo-controlled, clinical trial of high-dose supplementation with vitamins C and E, beta carotene, and zinc for age-related macular degeneration and vision loss: AREDS report no. 8. Arch Ophthalmol 119:1417–1436
Albanes D, Virtamo J, Rautalahti M, Haukka J, Palmgren J, Gref CG, Heinonen OP (1992) Serum beta-carotene before and after beta-carotene supplementation. Eur J Clin Nutr 46(1):15–24
Allen LH (2005) Multiple micronutrients in pregnancy and lactation: an overview. Am J Clin Nutr 81(suppl):1206S–1212S
Aoki K, Ito Y, Sasaki R, Ohtani M, Hamajima N, Asano A (1987) Smoking, alcohol drinking and serum carotenoid levels. Jpn J Cancer Res 78(10):1049–1056
Ascherio A, Stampfer MJ, Colditz GA, Rimm EB, Litin L, Willett WC (1992) Correlations of vitamin A and E intakes with the plasma concentrations of carotenoids and tocopherols among American men and women. J Nutr 122(9):1792–1801
Atkinson SA (2001) Special nutritional needs of infants for prevention of and recovery from bronchopulmonary dysplasia. J Nutr 131(3):942S–946S
Azais-Braesco V, Pascal G (2000) Vitamin A in pregnancy: requirements and safety limits. Am J Clin Nutr 71(suppl):1325S–133S
Ballew C, Bowmann BA, Sowell AL, Gillespie C (2001) Serum retinol distributions in residents of the United States: Third National Health and Nutrition Examination Survey, 1988–1994. Am J Clin Nutr 73:586–593
Barreto ML, Santos LMP, Assis AM, Araujo MP, Farenzena GG, Santos PA, Fiaccone RL (1994) Effect of vitamin A supplementation on diarrhoea and acute lower-respiratory-tract infections in young children in Brazil. Lancet 344:228–231
Basu S, Sengupta B, Roy Paladhi PK (2003) Single megadose vitamin A supplementation of Indian mothers and morbidity in breastfed young infants. Postgrad Med J 79:397–402
Bates CJ (1983) Vitamin A in pregnancy and lactation. Proc Nutr Soc 42:65–79
Bental RY, Cooper PA, Cummins RR (1994) Vitamin A therapy—effects on the incidence of bronchopulmonary dysplasia. S Afr J Food Sci Nutr 6:141–145
BgVV (Bundesinstitut für gesundheitlichen Verbraucherschutz und Veterinärmedizin) Pressedienst. Schwangere sollten weiterhin auf den Verzehr von Leber verzichten. 20/1995, 23.10.1995. http://www.bfr. bund.de/cms5w/sixcms/detail.php/775
Biesalski HK (1990) Separation of retinyl esters and their geometric isomers by isocratic adsorption high-performance liquid chromatography. Methods Enzymol 189:181–189
Biesalski HK (1995) Antioxidant vitamins in prevention. Dt. Ärzteblatt 92:1316–1321
Biesalski HK, Böhles H, Esterbauer H, Fürst P, Gey F, Hundsdörfer G, Kasper H, Sies H, Weisburger J (1997) Antioxidant vitamins in prevention. Clin Nutr 16:151–155
Biesalski HK, Hemmes C, Hopfenmuller W, Schmid C, Gollnick HP (1996) Effects of controlled exposure of sunlight on plasma and skin levels of beta-carotene. Free Radic Res 24(3):215–224
Brandt R, Kaugars GE, Riley WT, Dao Q, Silverman S Jr, Lovas JG, Dezzutti BP (1994) Evaluation of serum and tissue levels of beta-carotene. Biochem Med Metab Biol 51(1):55–60
Brandt RB, Müller DG, Schröder JR, Guyer KE, Kirkpatrick BV, Hutcher NE, Ehrlich FE (1978) Serum vitamin A in premature and term infants. J Pediatr 92:101–104
Bundesinstitut für Risikobewertung (BfR) Domke A, Großklaus R, Niemann B, Przyrembel H, Richter K, Schmidt E, Weißenborn A, Wörner B, Ziegenhagen R (2004) Verwendung von Vitaminen in Lebensmitteln - Toxikologische und ernährungsphysiologische Aspekte. BfR-Wissenschaft 03/2004
Castenmiller JJM, West CE, Linssen JPH, van het Hof KH, Voragen AGJ (1999) The food matrix of spinach is a limiting factor in determining the bioavailability of beta-carotene and to a lesser extent of lutein in humans. J Nutr 129:349–355
Chappell JE, Francis T, Clandinin MT (1985) Vitamin A and E content of human milk at early stages of lactation. Early Hum Dev 11:157–167
Chow CK, Thacker RR, Changchit C, Bridges RB, Rehm SR, Humble J, Turbek J (1986) Lower levels of vitamin C and carotenes in plasma of cigarette smokers. J Am Coll Nutr 5:305–312
Chytil F (1992) The lungs and vitamin A. Am J Physiol 262:L517–L527
Comstock GW, Menkes MS, Schober SE, Vuilleumier JP, Helsing KJ (1988) Serum levels of retinol, beta-carotene, and alpha-tocopherol in older adults. Am J Epidemiol 127(1):114–123
DACH (2000) Deutsche Gesellschaft für Ernährung, Österreichische Gesellschaft für Ernährung, Schweizerische Gesellschaft für Ernährungsforschung, Schweizerische Vereinigung für Ernährung. Referenzwerte für die Nährstoffzufuhr. Umschau/Braus, Frankfurt am Main
Darlow BA, Graham PJ (1999) Vitamin A supplementation for preventing morbidity and mortality in very low birth weight infants. The Cochrane Library 3:1–13
De Pee S, West CE, Muhilal, Karyadi D, Hautvast JG (1995) Lack of improvement in vitamin A status with increased consumption of dark-green leafy vegetables. Lancet 346:75–81
De Pee S, West CE, Permaesih D, Martuti S, Muhilal, Hautvast JG (1998) Orange fruit is more effective than are dark-green, leafy vegetables in increasing serum concentrations of retinol and beta-carotene in schoolchildren in Indonesia. Am J Clin Nutr 68:1058–1067
Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium and Zinc (2001) Institute of Medicine, Food and Nutrition Board, National Academy Press, Washington, DC
Dietary Reference Intakes for Vitamin C, Vitamin E, Selenium and Carotenoids (2000) Institute of Medicine, Food and Nutrition Board. National Academy Press, Washington, DC
Dijkhuizen MA, Wieringa FT, West CE, Muhilal (2004) Zinc plus β-carotene supplementation of pregnant women is superior to β-carotene supplementation alone in improving vitamin A status in both mothers and infants. Am J Clin Nutr 80:1299–1307
Dimitrov NV, Boone CW, Hay MB, Whetter P, Pins M, Kelloff GJ, Malone W (1986) Plasma beta carotene levels/kinetic patterns during administration of various doses of beta-carotene. J Nutr 3:227–237
Dimitrov NV, Meyer C, Ullrey DE, Chenoweth W, Michelakis A, Malone W, Boone C, Fink G (1988) Bioavailability of beta-carotene in humans. Am J Clin Nutr 48(2):298–304
Doyle W, Srivastava A, Crawford MA, Bhatti R, Brookee Z, Costeloe L (2001) Inter-pregnancy folate and iron status of women in an inner-city population Br J Nutr 86:81–87
FAO and WHO (1967) Requirements of vitamin A, thiamine, riboflavin and niacin, vol. 8. FAO, Food and Nutrition Series
FAO and WHO (1988) Requirements of vitamin A, iron, folate and vitamin B12, vol 23. FAO Food and Nutrition Series
FASEB (1995) Life Science Research Office. Third report on nutrition monitoring in the United States, vol 2. US Government Printing Office, Washington, DC
Federal Market Association for Livestock and Meat (2000)
Filteau SM, Morris SS, Abbott RA, Krkwood BR, Arthur P, Ross DA, Gyapong JO, Raynes JG (1993) Influence of morbidity on serum retinol of children in a community-based study in northern Ghana. Am J Clin Nutr 58:192–197
Geevarghese SK, Chytil F (1994) Depletion of retinyl esters in the lungs coincides with lung prenatal morphological maturation. Biochem Biophys Res Commun 200:529–535
Georgieff MK, Mammel MC, Mills MM, Gunter EW, Johnson DE, Thompson TR (1989) Effect of postnatal steroid administration on serum vitamin A concentrations in newborn infants with respiratory compromise. J Pediatr 114:301–304
Goldin GV, Opperman LA (1980) Induction of supernumerary tracheal buds and the stimulation of DNA synthesis in the embryonic chick lung and trachea by epidermal growth factor. J Embryol Exp Morphol 60:135–243
Goodman GE, Metch BJ, Omenn GS (1994) The effect of long-term beta-carotene and vitamin A administration on serum concentrations of alpha-tocopherol. Cancer Epidemiol Biomarkers Prev 3(5):429–432
Goodwin TW (1986) Metabolism, nutrition, and function of carotenoids. Annu Rev Nutr 6:273–297
Green HL, Phillips BL, Franck L, Filtmore CM, Said HM, Murrell JE, Moore ME, Briggs R (1987) Persistently low blood retinal levels during and after parenteral feeding of very low birth weight infants: examination of losses into intravenous administration sets and a method of prevention by addition to a lipid emulsion. Pediatrics 79:894–900
Gutcher GR, Lax AA, Farrell PM (1984) Vitamin A losses to plastic intravenous infusion devices and an improved method of delivery. Am J Clin Nutr 40:8–13
Haigh R, D‘Souza SW, Micklewright L, Gregory H, Butler SJ, Hollingsworth M, Donnai P, Boyd RD (1989) Human amniotic fluid urogastrone (epidermal growth factor) and fetal lung phospholipids. Br J Obstet Gynaecol 96:171–178
Hazinski TA (1990) Bronchopulmonary dysplasia. In: Chernick V (ed) Disorders of the respiratory tract in children. WB Saunders, Philadelphia, pp 300–320
Hume EM, Krebs HA (1949) Vitamin A requirements of human adults. Medical Research Council Special Report Series, His Majesty’s Stationary Office, London
Hustead VA, Gutcher GR, Anderson AA, Zachman RD (1984) Relationship of vitamin A (retinol) status to lung disease in the preterm infant. J Pediatr 105:610–615
IUPAC (1959) The vitamin A potency of beta-carotene. International Union of Pure and Applied Chemistry, London
Ito Y, Minohara M, Otani M, Sarai S, Niiya Y (1989) Effects of alcohol drinking and cigarette smoking on serum alpha- and beta-carotene concentrations in healthy adults. Nippon Eiseigaku Zasshi 44:607–614
Ito Y, Sasaki R, Minohara M, Otani M, Aoki K (1987) Quantitation of serum carotenoid concentrations in healthy inhabitants by high-performance liquid chromatography. Clin Chim Acta 169(2–3):197–207
Iyengar L, Apge SV (1972) Nutrient stores in human fetal livers. Br J Nutr 27:313–317
Jarvinen R, Knekt P, Seppanen R, Heinonen M, Aaran RK (1993) Dietary determinants of serum beta-carotene and serum retinol. Eur J Clin Nutr 47(1):31–41
Johnson EJ, Suter PM, Sahyoun N, Ribaya-Mercado JD, Russell RM (1995) Relation between beta-carotene intake and plasma and adipose tissue concentrations of carotenoids and retinoids. Am J Clin Nutr 62(3):598–603
Kitterman JA, Liggins GC, Clements JA, Campos G, Lee CH, Ballard PL (1981) Inhibitors of prostaglandin synthesis, tracheal fluid, and surfactant in fetal lambs. J Appl Physiol 51:1562–1567
Klein CJ (2002) Nutrient requirements for preterm infant formulas. J Nutr (suppl):132
Kohlhaufl M, Haussinger K, Stanzel F, Markus A, Tritschler J, Mühlhöfer A, Morresi-Hauf A, Golly I, Scheuch G, Jany BH, Biesalski HK (2002) Inhalation of aerosolized vitamin a: reversibility of metaplasia and dysplasia of human respiratory epithelia—a prospective pilot study. Eur J Med Res 7(2):72–78
Koo WWK, Krug-Wispe S, Succop P, Tsang RC, Neylan M (1995) Effects of different vitamin A intakes on very-low-birth-weight infants. Am J Clin Nutr 62:1216–1220
Kune GA, Bannerman S, Field B, Watson LF, Cleland H, Merenstein D, Vitetta L (1992) Diet, alcohol, smoking, serum beta-carotene, and vitamin A in male non-melanocytic skin cancer patients and controls. Nutr Cancer 18:237–244
Le Marchand L, Hankin JH, Carter FS, Essling C, Luffey D, Franke AA, Wilkens LR, Cooney RV, Kolonel LN (1994) A pilot study on the use of plasma carotenoids and ascorbic acid as markers of compliance to a high fruit and vegetable dietary intervention. Cancer Epidemiol Biomarkers Prev 3(3):245–251
Lorenzo JA, Quinton J, Sousa S, Raisz LG (1986) Effects of DNA and prostaglandin synthesis inhibitors on the stimulation of bone resorption by epidermal growth factor in fetal rat long-bone cultures. J Clin Invest 77:1897–1902
Majumdar SK, Shaw GK, Thomson AD (1983) Blood beta-carotene status in chronic alcoholics––a good biochemical marker for malnutrition. Drug Alcohol Depend 12:111–113
Marino PA, Rooney SA (1980) Surfactant secretion in a newborn rabbit lung slice model. Biochim Biophys Acta 620:509–519
Mathews-Roth MM (1975) Presence of carotenoids in the erythrocyte membranes of carotenemic and noncarotenemic individuals. Clin Chem 21(2):258–259
Mathews-Roth MM (1981) Carotenoids in medical applications. In: Bauernfeind JC (ed) Carotenoids as colorants and vitamin A precursors. Academic Press, New York, pp 755–785
Mathews-Roth MM (1990) Plasma concentrations of carotenoids after large doses of beta-carotene. Am J Clin Nutr 52(3):500–501
Mathews-Roth MM, Abraham AA, Gabuzda TG (1976) Beta-carotene content of certain organs from two patients receiving high doses of beta-carotene. Clin Chem 22(6):922–924
Mathews-Roth MM, Gulbrandsen CL (1974) Transport of beta-carotene in serum of individuals with carotenemia. Clin Chem 20:1578–1579
Metzler MD, Snyder JM (1993) Retinoic acid differentially regulates expression of surfactant-associated proteins in human fetal lung. Endocrinology 133:1990–1998
Meyer JC, Grundmann HP, Seeger B, Schnyder UW (1985) Plasma concentrations of beta-carotene and canthaxanthin during and after stopping intake of a combined preparation. Dermatologica 171:76–81
Micozzi MS, Brown ED, Edwards BK, Bieri JG, Taylor PR, Khachik F, Beecher GR, Smith JC Jr (1992) Plasma carotenoid response to chronic intake of selected foods and beta-carotene supplements in men. Am J Clin Nutr 55(6):1120–1125
Montreevasuat N, Olson JA (1979) Serum and liver concentration of vitamin A in Thai fetuses as a function of gestational age. Am J Clin Nutr 32:601–606
Mupanemunda RH, Lee DSC, Fraher LJ, Koura IR, Chance GW (1994) Postnatal changes in serum retinol status in very low birth weight infants. Early Hum Dev 38:45–54
Müller H (1996) Daily intake of carotenoids (carotenes and xanthophylls) from total diet and the carotenoid content of selected vegetables and fruit. Z Ernährungswiss 35:45–50
Nair PP, Judd JT, Berlin E, Taylor PR, Shami S, Sainz E, Bhagavan HN (1993) Dietary fish oil-induced changes in the distribution of alpha-tocopherol, retinol, and beta-carotene in plasma, red blood cells, and platelets: modulation by vitamin E. Am J Clin Nutr 58(1):98–102
Nationale Verzehrsstudie. Ergebnisse der Basisauswertung. Materialien zur Gesundheitsforschung. Schriftenreihe zum Programm der Bundesregierung Forschung und Entwicklung im Dienste der Gesundheit. Wirtschaftsverlag NW, Bonn 1991
Navarro J, Causse MB, Desquibet N, Herve F, Lallemand D (1984) The vitamin A status of low birth weight infants and their mothers. J Pediatr Gastroenterol Nutr 3:744–748
Neuzil KM, Gruber WC, Chytil F, Stahlman MT, Engelhardt B, Graham BS (1994) Serum vitamin A levels in respiratory syncytial virus infection. J Pediatr 124:433–436
Newman V (1993) Vitamin A and breast-feeding: a comparison of data from developed and developing countries. Wellstart International, San Diego (CA)
Newman V (1994) Vitamin A and breast-feeding: a comparison of data from developed and developing countries. Food Nutr Bull 15:161–176
Nierenberg DW, Stukel TA, Baron JA, Dain BJ, Greenberg ER (1989) Determinants of plasma levels of beta-carotene and retinal. Skin Cancer Prevention Study Group. Am J Epidemiol 130(3):511–521
Nierenberg DW, Stukel TA, Mott LA, Greenberg ER (1994) Steady-state serum concentration of alpha tocopherol not altered by supplementation with oral beta-carotene. The Polyp Prevention Study 1 Group. J Natl Cancer Inst 86(2):117–120
Nogee LM, Wispe JR, Clark JC, Weaver TE, Whitsett JA (1991) Increased expression of pulmonary surfactant proteins in oxygen-exposed rats. Am J Respir Cell Mol Biol 4:102–107
Oberg KC, Carpenter G (1989) EGF-induced PGE2 release is synergistically enhanced in retinoic acid treated fetal rat lung cells. Biochem Biophys Res Commun 162:1515–1521
Odom MJ, Snyder JM, Boggaram V, Mendelson CR (1988) Glucocorticoid regulation of the major surfactant associated protein (SP-A) and its messenger ribonucleic acid and of morphological development of human fetal lung in vitro. Endocrinology 123:1712–1720
Okabe T, Yorifuji H, Yamada E, Takaku F (1984) Isolation and characterization of vitamin-A-storing lung cells. Exp Cell Res 154:125–135
Olson JA (1994) Needs and sources of carotenoids and vitamin A. Nutr Rev 52(2 Pt 2):S67–S73
Olson JA, Gunning DB, Tilton RA (1984) Liver concentrations of vitamin A and carotenoids as a function of age and other parameters, of American children who died of various causes. Am J Clin Nutr 39:903–910
Papagaroufalis C, Cairis M, Pantazatou E, Megreli C, Xanthou M (1988) A trial of vitamin A supplementation in infants susceptible to bronchopulmonary dysplasia (BPD) in very-low-birth-weight (VLBW) infants. Pediatr Res 23:518A
Parker RS (1996) Absorption, metabolism, and transport of carotenoids. FASEB J 10(5):542–551
Pearson E, Bose C, Snidow T, Ransom L, Young T, Bose G, Stiles A (1992) Trial of vitamin A supplementation in very low birth weight infants at risk of bronchopulmonary dysplasia. J Pediatr 121:420–427
Peeples JM, Carlson SE, Werkman SH, Cooke RJ (1991) Vitamin A status of preterm infants during infancy. Am J Clin Nutr 53:1455–1459
Pelz R, Schmidt-Faber B, Heseker H (1998) Carotenoid intake in the German National Food Consumption Survey Z Ernährungswiss 37:319–327
Pinnock CB, Douglas RM, Badcock NR (1986) Vitamin A status in children who are prone to respiratory tract infections. Aust Paediatr J 22:95–99
Postaire E, Kouyate D, Rousset G, Regnault C, Lati E, Bejot M, Gossioux P, Hazebroucq G (1993) Plasma concentrations of beta-carotene, vitamin A and vitamin E after beta-carotene and vitamin E intake. Biomed Chromatogr 7(3):136–138
Prince MR, Frisoli JK (1993) Beta-carotene accumulation in serum and skin. Am J Clin Nutr 57(2):175–181
Rao CN, Rao BS (1970) Absorption of dietary carotenes in human subjects. Am J Clin Nutr 23(1):105–109
Rasmussen KM (1998) Vitamin A needs during pregnancy and lactation for the health of the mother and her fetus or infant. World Health Organization. Safe vitamin A dosage during pregnancy and lactation. Recommendations and report of a consultation
Recommended Dietary Allowances (1989) Food and Nutrition Board, Academy of Sciences, 10th edn. National Academy Press, Washington, DC
Riedl J, Linseisen J, Wolfram G (1997) Carotinoid-Aufnahme junger Erwachsener in Bayern. Z Ernährungswiss 36:61–62
Riemersma RA, Wood DA, Macintyre CC, Elton RA, Gey KF, Oliver MF (1991) Risk of angina pectoris and plasma concentrations of vitamins A, C, and E and carotene. Lancet 337:1–5
Ringer TV, DeLoof MJ, Winterrowd GE, Francom SF, Gaylor SK, Ryan JA, Sanders ME, Hughes GS (1991) Beta-carotene’s effects on serum lipoproteins and immunologic indices in humans. Am J Clin Nutr 53:688–694
Schneider R, Eberhardt W, Heseker H, Kübler W (1995) Vitamin intake and vitamin status in Germany. Scientific basis for vitamin intake in human nutrition. Bibl Nutr Dieta 52:116–127
Schulz C, Engel U, Kreienberg L, Biesalski HK (2006) Vitamin A and beta-carotene supply of women with gemini or short birth intervals: a pilot study. Eur J Nutr 46:12–20
Shah RS, Rajalekshmi R (1984) Vitamin A status of the newborn in relation to gestational age, body weight, and maternal nutritional status. Am J Clin Nutr 40:794–800
Shah RS, Rajalekshmi R (1987) Liver stores of vitamin A in human fetuses in relation to gestational age, fetal size and maternal nutritional status. British J Nutr 58:181–189
Shenai JP, Chytil F, Jhaveri A, Stahlman MT (1981) Plasma vitamin A and retinol-binding protein in premature and term neonates. J Pediatr 99:302–305
Shenai JP, Chytil F, Stahlman MT (1985) Vitamin A status of neonates with bronchopulmonary dysplasia. Pediatr Res 19:185–189
Shenai JP, Kennedy KA, Chytil F, Stahlman MT (1987) Clinical trial of vitamin A supplementation in infants susceptible to bronchopulmonary dysplasia. J Pediatr 111:269–277
Shenai JP, Mellen BG, Chytil F (2000) Vitamin A status and dexamethasone treatment in bronchopulmonary dysplasia. Pediatrics 106:547–553
Shenai JP, Rush MG, Stahlman MT, Chytil F (1990) Plasma retinol-binding protein response to vitamin A administration in infants susceptible to bronchopulmonary dysplasia. J Pediatr 116:607–614
Shensi JP, Chytil F, Stahlman MT (1985) Liver vitamin A reserves of very low birth weight neonates. Pediatr Res 19:892–893
Shirali GS, Oelberg DG, Mehta KP (1989) Maternal-neonatal serum vitamin A concentrations. J Pediatr Gastroenterol Nutr 9:62–66
Sichert-Hellert W, Kersting M, Manz F (2001) Changes in time-trends of nutrient intake from fortified and non-fortified food in German children and adolescents—15 year results of the DONALD study. Dortmund Nutritional and Anthropometric Longitudinally Designed Study. Eur J Nutr 40(2):49–55
Snyder JM (1991) The biology of the surfactant-associated proteins. In: Bourbon JR (ed) Pulmonary surfactant: biochemical, functional, regulatory, and clinical concepts. CRC Press, Boca Raton, pp 105–126
Stahlman MT (1984) Chronic lung disease following hyaline membrane disease. In: Stern L, Vert P (eds) Neonatal medicine. Masson, New York, pp 454–473
Stephensen CB, Alvarez JO, Kohatsu J, Hardmeier R, Kennedy JI Jr, Gammon RB Jr (1994) Vitamin A is excreted in the urine during acute infection. Am J Clin Nutr 60:388–392
Stofft E, Biesalski HK, Zschaebitz A, Weiser H (1992) Morphological changes in the tracheal epithelium of guinea pigs in conditions of “marginal” vitamin A deficiency. Int J Vit Nutr Res 62:134–142
Stryker WS, Kaplan LA, Stein EA, Stampfer MJ, Sober A, Willett WC (1988) The relation of diet, cigarette smoking, and alcohol consumption to plasma beta-carotene and alpha-tocopherol levels. Am J Epidemiol 127:283–296
Sundell HW, Gray ME, Serenius FS, Escobedo MB, Stahlman MT (1980) Effects of epidermal growth factor on lung maturation in fetal lambs. Am J Pathol 100:707–725
Suzukawa M, Ishikawa T, Yoshida H, Hosoai K, Nishio E, Yamashita T, Nakamura H, Hashizume N, Suzuki K (1994) Effects of alcohol consumption on antioxidant content and susceptibility of low-density lipoprotein to oxidative modification. J Am Coll Nutr 13:237–242
Takusa N, Sato S, Yamada T, Shimizu Y (1987) Epidermal growth factor (EGF) and tumor promoter 12-O-tetradecanoylphorbol 13-acetate (TPA) stimulate PG synthesis and thymidine incorporation in cultured porcine thyroid cells. Biochem Biophys Res Commun 143:880–884
Tang G, Serfaty-Lacrosniere C, Camilo ME, Russell RM (1996) Gastric acidity influences the blood response to a beta-carotene dose in humans. Am J Clin Nutr 64(4):622–626
Tyson JE, Wright LL, Oh W (1999) Vitamin A supplementation for extremely-low-birth-weight infants. N Engl J Med 340:1962–1968
Törrönen R, Lehmusaho M, Häkkinen S, Hänninen O, Mykkänen H (1996) Serum beta-carotene response to supplementation with raw carrots, carrot juice or purified beta-carotene in healthy non-smoking women. Nutr Res 16:565–575
Underwood BA (1994) Maternal vitamin A status and its importance in infancy and early childhood. Am J Clin Nutr 59:517S–522S
Van Het Hof KH, Brouwer I, West CE, Haddeman E, Steegers-Theunissen RPM, Van Dusseldorp M, Weststrate JA, Eskes TKAB, Hautvast JGAJ (1999) Bioavailability of lutein from vegetables is 5 times higher than that of beta-carotene. Am J Clin Nutr 70:261–268
Verma RP, McCulloch KM, Worrell L, Vidyasagar D (1996) Vitamin A deficiency and severe bronchopulmonary dysplasia in very low birth weight infants. Am J Perinatol 13:389–393
Villard L, Bates CJ (1986) Carotene dioxygenase (EC 1.13.11.21) activity in rat intestine: effects of vitamin A deficiency and of pregnancy. Br J Nutr 56(1):115–122
Von Laar J, Stahl W, Bolsen K, Goerz G, Sies H (1996) Beta-carotene serum levels in patients with erythropoietic protoporphyria on treatment with the synthetic all-trans isomer or a natural isomeric mixture of beta-carotene. J Photochem Photobiol B 33(2):157–162
Weaver TE, Whitsett JA (1991) Function and regulation of expression of pulmonary surfactant-associated proteins. Biochem J 273:249–264
Werkman SH, Peeples JM, Cooke RJ, Tolley EA, Carlson SE (1994) Effect of vitamin A supplementation of intravenous lipids on early vitamin A intake and status of premature infants. Am J Clin Nutr 59:586–595
West CE, Castenmiller JJ (1998) Quantification of the “SLAMENGHI” factors for carotenoid bioavailability and bioconversion. Int J Vitam Nutr Res 68(6):371–377
West KP, Katz J, Khatry SK, LeClerq SC, Pradhan EK, SR Shrestha, Connor PB, Dali SM, Christian P, Pokhrel RP, Sommer A (1999) Double blind, cluster randomised trial of low dose supplementation with vitamin A or β carotene on mortality related to pregnancy in Nepal. BMJ 318:570–575
Willett WC, Stampfer MJ, Underwood BA, Taylor JO, Hennekens CH (1983) Vitamins A, E, and carotene: effects of supplementation on their plasma levels. Am J Clin Nutr 38(4):559–566
Woodruff CW, Latham CB, Bactier H, Hewett JE (1987) Vitamin A status of preterm infants: correlation between plasma retinol concentration and retinol dose response. Am J Clin Nutr 46:985–988
Woodruff CW, Latham CB, James EP, Hewett JE (1986) Vitamin A status of preterm infants: the influence of feeding and vitamin A supplements. Am J Clin Nutr 44:384–389
Xu MJ, Plezia PM, Alberts DS, Emerson SS, Peng YM, Sayers SM, Liu Y, Ritenbaugh C, Gensler HL (1992) Reduction in plasma or skin alpha-tocopherol concentration with long-term oral administration of beta-carotene in humans and mice. J Natl Cancer Inst 84(20):1559–1565
Zachman RD (1989) Retinol (vitamin A) and the neonate: special problems of the human premature infant. Am J Clin Nutr 50:413–424
Zachman RD, Tsao FHC (1988) Retinyl ester synthesis by isolated adult rabbit lung-type II cells. Int J Vit Nutr Res 58:161–165
American Academy of Pediatrics, Committee on Nutrition (1998) Nutritional needs of pretern infants. Pediatric Nutrition Handbook (Kleinman RE, ed.), American Academy of Pediatrics, EIK Grove Village (IL), 55–87
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Strobel, M., Tinz, J. & Biesalski, HK. The importance of β-carotene as a source of vitamin A with special regard to pregnant and breastfeeding women. Eur J Nutr 46 (Suppl 1), 1–20 (2007). https://doi.org/10.1007/s00394-007-1001-z
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00394-007-1001-z