
Abstract
Background and aims
Obesity is a major risk factor for the development of colon cancer. Secretion of the hormone leptin from adipocytes is increased in obesity, and serum levels are proportional to body fat mass. Serum leptin levels are an independent risk factor for colon cancer. Leptin receptors are expressed in normal, premalignant and malignant colonic epithelia. We have investigated the effects of leptin on proliferation and apoptosis of colonic cancer cells and the early signalling events involved.
Methods
Proliferation of HT-29 colon cancer cells in response to leptin was assessed by 3-[4, 5-dimethylthiazol-2-y-l]-2, 5-diphenyltetrazolium bromide (MTT) assay, and apoptosis was quantified by enzyme-linked immunosorbent assay (ELISA) for intracellular nucleosomes. Signalling pathways involved were determined by using specific inhibitors, quantification of phosphorylated active intermediates and ELISA of active nuclear-translocated transcription factors.
Results
Leptin stimulated HT-29 cell proliferation and inhibited both serum-starvation and celecoxib-induced apoptosis. The proliferative and anti-apoptotic effects of leptin were abolished by inhibition of JAK2 with AG490, phosphatidylinositol 3′-kinase (PI3 kinase) with LY294002 and c-Jun NH2-terminal kinase (JNK) with SP600125. Stimulation of HT-29 cells with leptin increased phosphorylation of JAK2, Akt and JNK. Activation of JAK2 was upstream of PI3 kinase/Akt but not of JNK. Activation of JAK2 was followed by activation and nuclear translocation of STAT3 and JNK activation led to increased activator protein 1 (AP-1) transcriptional activity.
Conclusions
Leptin stimulates proliferation and inhibits apoptosis in human colon cancer cells and may be an important factor in the increased incidence of colon cancer in obesity. This effect involves JAK2, PI3 kinase and JNK and activation of the oncogenic transcription factors signal transducer and activator of transcription (STAT)3 and AP-1.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Colorectal cancer is a major cause of mortality and morbidity worldwide, with an annual incidence of about a million and an annual mortality of about half a million [1]. There is a strong association between colon cancer and obesity [2, 3]. Many factors, including those related to lifestyle, may underlie this association, but factors produced by the increased fat mass may be involved. The adipokine leptin is elevated in the serum of obese people in direct correlation to fat mass [4] and may be an important biological link between obesity and colon cancer. Recent evidence suggests that leptin levels are an independent risk factor for colonic cancer [5].
The biological actions of leptin are best documented in relation to energy homeostasis, feeding and body weight, and this is dramatically demonstrated in the naturally occurring massively obese ob/ob mice that lack leptin or the db/db mice and fa/fa rats that lack functional leptin receptors. However, recent work has demonstrated that leptin has a wider spectrum of activity, including growth-promoting actions in various tissues. The presence of a leptin receptor (Ob-R) has been demonstrated on normal, precancerous and malignant human colonic epithelium as well as colon cancer cell lines [6, 7]. The leptin receptor is a member of the class I cytokine receptor family. The long receptor variant (Ob-Rb) is the main signal-transducing variant; it contains the amino acid residues crucial for receptor signalling [8]. Initial studies have shown that leptin induces proliferation and invasiveness of colon cancer cells both in vivo and in vitro [6, 7], although the cellular mechanisms responsible for these actions remain to be determined. Anti-apoptotic actions of leptin have been described [9], although pro-apoptotic effects were reported for leptin in human bone marrow stromal cells [10].
The intracellular signalling mechanisms which mediate cell proliferation, cell survival and apoptosis have attracted investigation because these mechanisms are crucial in the potential carcinogenic effects of leptin and important in any potential screening and therapeutic interventions. Activation of the leptin receptor has been reported to activate multiple signalling pathways in different systems, but the mechanisms and sequelae of those involved in the regulation of colonic epithelial function remain to be determined. Inhibition of apoptosis is an important promoter of epithelial carcinogenesis. We have hypothesized that leptin provides a link between obesity and colon cancer by promoting excessive proliferation and inhibiting apoptosis. In this study, we have explored the mechanisms involved in these effects.
Whilst a variety of transduction systems may promote proliferation and cell survival, in this study, we have investigated the involvement of the phosphatidylinositol-3′ kinase (PI3 kinase) /Akt, the c-Jun NH2-terminal kinase (JNK) and the JAK2/STAT3 pathways. Akt (protein kinase B), a serine/threonine protein kinase, is an important mediator of cell survival [11]. It is known to prevent apoptosis in various cells [12]. It is translocated to the cell membrane and subsequently activated by a process initiated by phosphorylation of membrane phospholipids by PI3 kinase present adjacent to the mammalian cell membrane [13]. The lipid kinase activity of PI3 kinase is activated by various cytokines, hormones and growth factors activating their respective receptors. The anti-apoptotic effect of leptin on hepatic stellate cells was mediated by phosphorylation of Akt [9], and in prostate cancer cells, leptin activated Akt [13].
The mitogen-activated protein (MAP) kinase system also transduces proliferation and cell survival signals. The principal action of JNK is the regulation of activator protein 1 (AP-1) activity, and both pro- and anti-apoptotic actions of JNK have been reported. Activation of JNK is important in the proliferative actions of some growth factors in colon cancer [14], and we have recently shown that JNK mediates the anti-apoptotic effects of glycine-extended gastrin in HT-29 colon cancer cells [15]. Leptin has recently been shown to induce prostate cancer cell proliferation via activation of JNK [16].
When leptin binds to its signal-transducing long-chain receptor isoform (Ob-Rb), oligomerization of the receptor chains leads to activation of Janus tyrosine kinases (JAKs), which are closely associated with cytokine-type receptors. Activated JAKs, in turn, phosphorylate signal transducers and activators of transcription (STATs). One member of the JAK family, JAK2, has been shown to mediate anti-apoptosis in haematopoietic cells [17] and seems to be the JAK enzyme most involved in signalling via Ob-Rb [18]. The STAT proteins in turn control the transcription of a variety of genes; STAT3 is regarded as an oncogene, promoting proliferation and inhibiting apoptosis. Activation of STAT3 by leptin in the hypothalamus and in gastric cancer cells has been reported [19, 20]. Limited data suggest that STAT3 is upregulated in colonic neoplasia and preneoplasia [21].
In this study, we have investigated the effect of leptin on proliferation and apoptosis in the HT-29 colon cancer cell line because this has been shown to express the fully functional long leptin receptor [7]. We have used celecoxib to induce apoptosis because of the direct relation to clinical practice. The cyclo-oxygenase 2 (COX-2) inhibitor celecoxib reduces the incidence of adenomatous polyps in high-risk patients [22] and animal models [23], and has been approved by the Food and Drug Administration in the USA for chemoprevention in familial adenomatous polyposis. Celecoxib induces apoptosis in cancer cell lines that do not express COX-2, consistent with data showing that non-steroidal anti-inflammatory drugs (NSAIDs) and COX-2 inhibitors have beneficial actions in chemoprevention by targeting multiple pathways [24].
Methods
Cell culture
HT-29 human colon cancer cells were grown in Dulbecco’s modified Eagle medium (DMEM) containing 4,500 mg/l glucose, 100 mg/l penicillin, 100 mg/l streptomycin and 2 mM l-glutamine, and supplemented with 10% foetal bovine serum (FBS) in 5% CO2/95% air at 37°C. Cells were subcultured following enzymatic digestion using trypsin/EDTA solution.
Assessment of cell proliferation and apoptosis
In 48-well plates, 5×104 cells per well were cultured in 10% FBS containing medium for 24 h and then serum-starved for 24 h. The medium was subsequently changed to fresh serum-free medium, inhibitors were added as appropriate and cells were treated with leptin and/or celecoxib. After a further 24 h in culture, cell proliferation was assessed using the MTT [3-(4, 5-dimethylthiazol-2-y-l)-2, 5-diphenyltetrazolium bromide] colorimetric assay as previously described [25, 26], and apoptosis was quantified by measuring intracellular nucleosome concentration in cell lysates [27, 28] using the cell-death enzyme-linked immunosorbent assay (ELISA) kit (Roche, Germany) according to the manufacturer’s instructions.
Nuclear extraction and transcription factor assays
Confluent HT-29 cells in 6-well plates were serum-starved for 24 h. After changing to fresh serum-free medium and treatment with kinase inhibitors for 1 h, cells were treated with leptin. After 30 min incubation, nuclear extraction was performed using a nuclear extraction kit (Active Motif, Belgium). Protein concentration was quantified using a Bio-Rad protein assay (Bio-Rad, UK) with bovine serum albumin as a standard. Yield was 0.15±0.005 mg/well. Nuclear translocation of active STAT3 was assessed using the STAT3 TransAM ELISA, and nuclear levels of active AP-1 were quantified using the AP-1 c-Jun TransAm ELISA (both from Active Motif) according to the manufacturer’s instructions. For the AP-1 assay, 8 μg of nuclear extract protein per well was used, and 12 μg was used for the STAT3 assay. These assays utilise oligonucleotides containing appropriate consensus binding sites immobilised on the wells of a 96-well plate to capture active AP-1 or STAT3 contained in the nuclear extracts. After binding of a primary antibody directed against the transcription factor, quantitation was performed using a horseradish-peroxidase-conjugated secondary antibody and substrate solution.
Detection of activated Akt and JNK
In 96-well plates, 104 cells per well were subcultured for 72 h in serum-containing medium and then in serum-free medium for a further 24 h. Cells were then treated with inhibitors for 1 h followed by leptin. After 15 min incubation, cells were fixed with 4% formaldehyde. Non-phosphorylated and active phosphorylated Akt and JNK were quantified using specific FACE ELISA kits (Active Motif) according to the manufacturer’s instructions.
Detection of activated JAK2
Tyrosine-phosphorylated JAK2 was quantified using a cell-based ELISA. Cells were treated, prepared and fixed as for the Akt and JNK assays. After three washes (5 min each time with agitation at 100 rpm on a plate shaker) with wash buffer [phosphate-buffered saline (PBS) containing 0.1% Triton X-100], endogenous peroxidase was then quenched by incubating for 30 min in wash buffer containing 1% hydrogen peroxide and 0.1% sodium azide. After two further washes with wash buffer, fixed cells were incubated for 1 h with 3% non-fat dried milk to block non-specific binding. After two further washes in wash buffer, primary antibodies diluted 1:250 in PBS/0.05% Tween 20 were added and incubated overnight at 4°C. The antibodies used were rabbit polyclonal antibodies; anti-phospho-JAK2 (Tyr 1007/Tyr 1008) (Santa Cruz Biotechnology, sc-16566-R) and anti-JAK2 (Santa Cruz, sc-278). The specific blocking peptides (Santa Cruz) were added at 10 μg/ml final concentration to confirm specificity. After removal, the primary antibody cells were washed three times for 5 min each with wash buffer and then incubated for 1 h with horseradish-peroxidase-conjugated goat anti-rabbit immunoglobulin (Ig)G (Sigma) diluted 1:1,000 in PBS/0.05% Tween 20. Wells were then washed three times with wash buffer and twice with PBS, each for 5 min. Substrate solution (o-phenylenediamine dihydrochloride, Sigma FAST OPD) was used for quantification using an optical density of 450 nm (OD450 nm) with a reference wavelength of 650 nm. Subsequently, ELISA readings were corrected for the actual cell numbers in each well by performing crystal violet staining. Wells were washed twice with wash buffer and twice with PBS and air-dried for 5 min. After staining with crystal violet for 30 min, the excess stain was removed by washing in PBS. Incubation in 1% sodium dodecyl sulphate for 60 min was performed to extract the stain, and the OD595 nm was determined. Corrected JAK/phospho-JAK2 levels were determined by dividing the OD450 by OD595.
Chemicals and reagents
Recombinant human leptin SP600125 and AG490 were purchased from Merck (Nottingham, UK). LY294002 and celecoxib were from Alexis Biochemicals (Nottingham, UK). MTT was purchased from Sigma (Poole, UK). All cell culture media and supplements were from Invitrogen (UK). When appropriate, stock solutions of inhibitors were dissolved in dimethyl sulphoxide (DMSO). The final concentration of DMSO in experiments was always less than 0.1%, and control wells contained an equivalent concentration. Inhibitors were added 1 h prior to the addition of leptin in all experiments.
Statistical tests
Proliferation and cell toxicity experiments were performed in triplicate wells. Nuclear extracts and transcription factor analyses in duplicate and phosphorylated kinase assays in quadruplicate wells. Each experiment was repeated three to six times. Results are expressed as mean±SEM. One-way analysis of variance was used for dose–response curves, and the paired t tests were used to analyse the effect of inhibitors. A P value less than 0.05 was regarded as significant.
Results
Leptin stimulates proliferation of HT-29 cells
Leptin stimulated an increase in HT-29 cell numbers after 24 h incubation. A dose-dependent increase in proliferation was seen over the concentration range 0.1–10 nM (Fig. 1). The maximal effect was 39±8% above basal with 10 nM leptin, and this concentration of leptin was used for further investigation of the signalling pathways.

Effect of leptin on HT-29 cell proliferation. Serum-starved HT-29 cells were treated for 24 h with increasing concentrations of leptin. Cell proliferation after 24 h was quantified by MTT assay. Results expressed as percentage above untreated control cells, mean±SEM, N=6, **P<0.01 vs basal
Leptin inhibits apoptosis in HT-29 cells
Leptin significantly reduced apoptosis produced either by serum starvation or celecoxib treatment. Preliminary experiments confirmed a potent pro-apoptotic effect of celecoxib. A fourfold increase in apoptosis was produced by a concentration of 25 μM, and this was used for further characterisation. Lower concentrations of celecoxib (<10 μM) did not produce appreciable apoptosis despite being as effective as higher doses in suppressing COX-2-derived PGE2 production [28]. Leptin (10 nM) reduced serum-starvation-induced apoptosis by 48% and celecoxib-induced apoptosis by 65% (Fig. 2). Leptin alone was as effective as culture medium containing 10% FBS in reducing celecoxib-induced apoptosis (data not shown).

Effect of leptin on HT-29 cell apoptosis. Serum-starved HT-29 cells were incubated with leptin (10 nM) and/or celecoxib (25 μM) for 24 h. Intracellular nucleosomes were quantified by ELISA as an assessment of apoptosis. A405 nm–A490 nm is proportional to nucleosome concentration. Values were corrected for cell numbers quantified using parallel MTT assays. Mean±SEM, N=3–4, *P<0.05 vs celecoxib-treated, **P<0.01 vs control cells in serum-free medium
Leptin promotes cell survival in celecoxib-treated HT-29 cells
The inhibition of apoptosis in celecoxib-treated cells by leptin was translated into an increase in overall viable cell numbers in celecoxib-treated cells. Celecoxib produced a dose-dependent reduction in cell numbers, consistent with the increased induction of apoptosis. Celecoxib (25 μM) reduced viable cell numbers by 55±7%, and leptin ameliorated this reduction in cell numbers by 50% (Fig. 3). Leptin did not reverse the reduction in viable cell numbers produced by the highest concentrations of celecoxib.

Effect of leptin on cell numbers in celecoxib-treated cells. Serum-starved HT-29 cells were incubated with celecoxib±leptin (10 nM) for 24 h. Cell number after 24 h was quantified by MTT assay. Results expressed as percentage of untreated cells in serum-free medium. Mean±SEM, N=4, *P<0.05 vs control, **P<0.01 vs control, ‡ P<0.05 vs celecoxib-treated cells
The growth-promoting and cell survival effects of leptin are mediated by Akt, JNK and JAK2
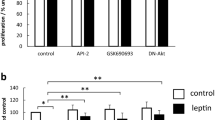
Inhibition of either the PI3 kinase/Akt pathway with the PI3 kinase inhibitor LY294002 or JNK with SP600125 abolished both the proliferative and cell survival effects of leptin (Fig. 4). Neither of the inhibitors alone had a significant effect on either basal or celecoxib-treated cell numbers. However, in the presence of LY294002 or SP600125, leptin failed to increase cell numbers in either control or celecoxib-treated cells. The specific JAK2 inhibitor AG490 also abolished the proliferative and cell survival effects of leptin (Fig. 4).

Effect of inhibition of JNK, Akt and JAK2 on proliferative and cell survival effects of leptin. Serum-starved HT-29 cells were pretreated for 60 min with inhibitors of JNK (SP600126; 10 μM,), PI3 kinase/Akt (LY294002; 10 μM) and JAK2 (AG490; 25 μM) before 24 h treatment with leptin (10 nM)±celecoxib (25 μM). a Cell proliferation after 24 h was quantified by MTT assay. Results expressed as percentage of control untreated cells, mean±SEM, N=4, **P<0.01 vs control (top). b Cell survival in celecoxib-treated cells was assessed after 24 h by MTT assay. Results expressed as percentage of control untreated cells, mean±SEM, N=5, ‡ P<0.05 vs celecoxib-treated cells (bottom)
Inhibition of apoptosis by leptin requires signalling via Akt, JNK and JAK2
To confirm that the effects of the kinase inhibitors on increased cell survival were due to preventing the anti-apoptotic effect of leptin, the effect of pharmacological inhibition of JNK, Akt and JAK2 on apoptosis, as measured by intracellular nucleosome release, was assessed. Inhibition of JAK2 with AG490 produced a significant increase (400%) in apoptosis, LY294002 produced a smaller, although significant increase in apoptosis (190%), but SP600125 had no effect on basal apoptosis. No anti-apoptotic effect of leptin was seen in the presence of inhibition of either JNK with SP600125, Akt with LY294002 or JAK2 with AG490 (Fig. 5).

Effect of inhibition of JNK, Akt and JAK2 on the anti-apoptotic effect of leptin. Serum-starved HT-29 cells were pretreated for 60 min with inhibitors of JNK (SP600126; 10 μM,), PI3 kinase/Akt (LY294002; 10 μM) and JAK2 (AG490; 25 μM) before 24 h treatment with leptin (10 nM). Apoptosis was quantified with ELISA for intracellular nuclosomes, A405 nm–A490 nm is proportional to nucleosome concentration. Mean±SEM, N=3, **P<0.01 vs serum-free control
Leptin increases phosphorylation of JNK, Akt and JAK2
To confirm the results with the pharmacological inhibitors, we examined whether leptin increased phosphorylation of the implicated signalling intermediates. Leptin increased Akt phosphorylation by 213% and JNK phosphorylation by 228% after 15 min stimulation (Fig. 6). SP600125 abolished JNK phosphorylation, and LY294002 abolished Akt phosphorylation, but SP600125 did not affect Akt activation, and LY294002 failed to reduce JNK phosphorylation. Celecoxib reduced leptin-stimulated JNK phosphorylation by 51% and Akt phosphorylation by 55%, but in both cases, the degree of inhibition was markedly less than that seen with the specific kinase inhibitors LY294002 and SP600125 (Fig. 6). Inhibition of JAK2 with AG490 had no effect on leptin-stimulated JNK phosphorylation but abolished Akt phosphorylation induced by leptin (Fig. 6).

Effect of leptin on phosphorylation of Akt (a; top) and JNK (b; bottom). Serum-starved HT-29 cells were pretreated with inhibitors of JAK2 (AG490; 25 μM), PI3 kinase [LY294002; 10 μM (LY)], JNK [SP600125; 10 μM (SP)] or celecoxib [25 μM (cele)] for 60 min before stimulation with leptin (10 nM) for 15 min. Phosphorylated and total Akt and JNK were assessed by ELISA. Results expressed as phosphorylated/total enzyme corrected for cell number as assessed by subsequent crystal violet staining. Mean±SEM, N=3, **P<0.01 vs control, ‡ P<0.05 vs leptin, ‡‡ P<0.05 vs leptin
Leptin increased JAK2 tyrosine phosphorylation by 1.93-fold compared with basal but had no significant effect on total JAK2 as determined with the ELISA. The specificity of the assays was demonstrated by the effective competition by the specific blocking peptides without any reciprocal inhibition (Fig. 7). Leptin stimulation of JAK2 phosphorylation was abolished by AG490 (Fig. 7).

Effect of leptin on JAK2 phosphorylation. Serum-starved HT-29 cells were pretreated with AG490 (25 μM) for 60 min before stimulation with leptin (10 nM) for 10 min. Phosphorylated and total JAK2 were assessed by cell-based ELISA in formalin-fixed cells. a Effect of leptin on total JAK2 levels and effect of JAK2 and phospho-JAK2 blocking peptides on detection of JAK2. b Effect of leptin on phospho-JAK2 levels and effect of JAK2 and phospho-JAK2 blocking peptides on detection of phospho-JAK2. c Effect of leptin on phospho-JAK2/JAK2 ratio. Mean±SEM, N= 3, *P<0.05 vs basal, ‡‡ P< 0.01 vs leptin
Leptin increases activation of the transcription factors AP-1 and STAT3
To further determine the significance of the signalling pathways implicated in the anti-apoptotic effects of leptin, we examined activation of the transcription factors AP-1 and STAT3. Leptin increased AP-1 activation 1.9-fold above basal, and this was abolished by the JNK inhibitor SP600125 but not inhibited by either the PI3 kinase inhibitor LY294002 or the JAK2 inhibitor AG490. Celecoxib reduced AP-1 activation by 48% (Fig. 8). Leptin was also a potent stimulus to nuclear translocation of active STAT3 (Fig. 8): 30 min after leptin stimulation, nuclear translocation of STAT3 had increased 2.2-fold. This stimulation was blocked by AG49 but was unaffected by inhibition of JNK with SP600125, Akt with LY294004 or celecoxib (Fig. 8).

Effect of leptin on activation of AP-1 (a; top) and STAT3 (b; bottom). Serum-starved HT-29 cells were incubated with inhibitors AG490 (25 μM), LY294002 (10 μM), SP600125 (10 μM) or celecoxib (25 μM) for 60 min before incubation with leptin (10 nM) for 30 min. a Active AP-1 and b STAT3 in nuclear extracts were quantified by ELISA. OD450 nm is proportional to transcription factor concentration. Mean±SEM, N=4, **P< 0.01 vs basal, ‡ P<0.05 vs leptin, ‡‡ P<0.01 vs leptin
Discussion and conclusions
Abnormal apoptosis and cell proliferation characterize the aetiology and progression of cancer. Leptin, an adipocyte-secreted hormone elevated in the blood of obese people, is a direct risk factor for colon cancer [5]. Consistent with these conclusions from epidemiological studies, we have confirmed a pro-proliferative effect of leptin on colon cancer cells. This finding is in agreement with previous reports in the literature [6, 7]. The anti-apoptotic effect of leptin demonstrated here suggests strongly that the overall increase in colonic epithelial cell number as a result of leptin treatment is due to a combination of stimulation of cell proliferation and inhibition of apoptosis by leptin. The concentrations of leptin used in the study correspond to those found circulating in normal and obese subjects, suggesting that these effects of leptin are physiologically relevant [29, 30].
The main aim of this study was to further elucidate the mechanisms of leptin actions on colonic epithelial cells. Consistent with this, we examined the involvement of PI3 kinase/Akt, JNK and JAK2 in colonic epithelial leptin signalling. Pharmacological inhibition and direct detection of phosphorylated intermediates showed that these kinases are activated by leptin upon binding to its receptor, and they consequently mediate the proliferative and anti-apoptotic effects. The effects of the pharmacological inhibitors showed that activation of Akt is independent of activation of JNK. As expected, the specific pharmacological inhibitor of PI3 kinase LY294002 inhibited Akt phosphorylation. This confirms that PI3 kinase is upstream of, and activates, Akt [31]. Interestingly, inhibition of JAK2 abolished Akt phosphorylation but not JNK phosphorylation, showing that JAK2 is upstream of Akt in colonic epithelial leptin signalling, and that the JNK pathway is apparently activated separately. Our finding of a separate and PI3-kinase-independent activation of JNK by leptin in colonic epithelial cells contrasts with the report of PI3-kinase-induced activation of JNK in human T cells [32]. The human Ob-Rb contains a JAK-binding/activation site and five potential tyrosine phosphorylation sites in the intracellular portion [33]; whether activation of PI3 kinase and signalling in colon cancer occurs via docking on one specific receptor phosphotyrosine site or via a downstream-associated docking protein such as IRS-1 remains to be identified [33]. Leptin stimulated JNK phosphorylation, and inhibition of JNK with SP600125 abolished the proliferative and anti-apoptotic effects of leptin. Further studies will be required to determine the processes linking the leptin receptor to JNK activation. Other JAK proteins, particularly JAK1, have been shown to be able to mediate Ob-R signalling [33], and further studies are underway to elucidate the role of other kinases and guanosine triphosphate (GTP)-binding proteins in mediating coupling to JNK.
We also demonstrated that celecoxib inhibited leptin-induced activation of JNK and Akt. Concentrations of celecoxib that significantly blocked proliferation and induced apoptosis reduced Akt and JNK activation by approximately 50%. We and others [24, 28] have shown that celecoxib induces apoptosis and reduces cell numbers in a COX-2-independent manner. Previous studies have suggested that celecoxib can inhibit activation of the PI3 kinase/Akt pathway by inhibiting PDK1, which is the enzyme most commonly responsible for phosphorylating the membrane-directed Akt [34], and our data would be consistent with this finding. However, the inhibition of JNK phosphorylation by celecoxib is a novel finding which adds further complexity to the multiple sites of chemopreventative actions of celecoxib. Further studies will be required to determine the specific site in the signalling pathway at which celecoxib inhibits JNK activation. It is noteworthy that despite the potent pro-apoptotic effect of celecoxib, leptin still produced a major inhibition of apoptosis; perhaps, the multiplicity of anti-apoptotic signals generated by leptin overcame the multiple actions of celecoxib.
Leptin increased activation of the transcription factor AP-1, which regulates various cell processes, including apoptosis and cell cycle progression, and proliferation by regulating gene transcription and AP-1 sites have been found in many genes involved in tumour transformation [35]. AP-1 activation and subsequent transcriptional activities are regulated by the MAP kinases [p38 MAP kinase, JNK and extracellular-signal-related kinase (ERK)] [35, 36]. Consistent with this, we found that leptin-induced activation of AP-1 was inhibited by specific inhibition of JNK but not PI3 kinase or JAK2. Consistent with the important role of JNK in phosphorylating and activating the component parts of the AP-1 complex, we have shown that the inhibition of JNK activity celecoxib was associated with a comparable reduction in AP-1 activation. Consequently, we conclude that AP-1 is a likely downstream mediator of JNK-dependent leptin anti-apoptotic and pro-proliferative effects in colon cancer. This finding is consistent with previously published reports that AP-1 is induced by mitogenic signals, and that it mediates tumour promotion and cell proliferation in intestinal epithelial cells and plays an important role in colon cancer [37, 38].
Our study also showed that leptin induced nuclear translocation of activated STAT3, and that phosphorylation and activation of STAT3 were dependent on JAK2 activation. STAT3 is regarded as an oncogene; increased activation is associated with cell transformation, increased proliferation and inhibition of apoptosis. Increased STAT3 activation has been reported in the polyp-cancer sequence in colon cancer [21], and this enhanced activation seems driven by increased growth factor stimulation rather than activating STAT3 mutations [39]. It is possible that leptin drives in vivo STAT3 activation. This hypothesis requires further examination. Inhibition of JAK2 blocked the anti-apoptotic effect of leptin and STAT3 activation, and it is likely that STAT3 may be an important cell proliferative mechanism for leptin in the colon; leptin-induced STAT3 activation was recently reported in gastric cancer cells [19]. The exact STAT3 target genes involved require further investigation, but in other systems, STAT3 activates c-fos and the anti-apoptotic genes survivin and Bcl-XL [40].
Several studies have illustrated the complexity of signalling events initiated by leptin: in kidney cells, leptin stimulated invasion via pathways involving Rac1, RhoA, protein kinase C, Akt, mammalian target of rapamycin (mTOR) and JAK2, although it is not yet clear either how these pathways interact with each other or what the downstream targets are [6]. In gastric cancer cells leptin stimulated separate activation of the ERK and STAT3 pathways [19]. One recent study has shown that leptin ameliorates the apoptotic action of butyrate in colon cancer cells: this effect was associated with activation of ERK and nuclear translocation of nuclear factor kappa B (NF-κB) [41].
In conclusion, we report anti-apoptotic and pro-proliferative effects for leptin in colon cancer cells. These actions of leptin suggest that it plays a role in the aetiology and progression of colon cancer, and contributes to the increased risk for colon cancer in obesity. Leptin activates at least three pathways essential to proliferation and inhibition of apoptosis: PI3-kinase-dependent activation of Akt, JNK-dependent activation of AP-1 and JAK2-dependent activation of STAT3. Furthermore, JAK2 is upstream of Akt in leptin signalling in colonic epithelial cells. Leptin ameliorated the potentially beneficial growth-inhibitory and pro-apoptotic actions of celecoxib, and the hyperleptinaemia characteristic of obesity may render such chemopreventative strategies less effective. Further studies exploring the actions of leptin on colonic epithelial dynamics in vitro and in vivo are warranted to define chemopreventative and therapeutic strategies, but inhibition of signalling from the leptin receptor may have potential for beneficial actions in treating and preventing colon cancer.
Abbreviations
- AP-1:
-
activator protein 1
- COX:
-
cyclo-oxygenase
- DMEM:
-
Dulbecco’s modified Eagle medium
- DMSO:
-
dimethyl sulphoxide
- ELISA:
-
enzyme-linked immunosorbent assay
- FBS:
-
foetal bovine serum
- JAK:
-
Janus tyrosine kinase
- JNK:
-
c-Jun NH2-terminal kinase
- MAP:
-
mitogen-activated protein kinase
- MTT:
-
3-[4, 5-dimethylthiazol-2-y-l]-2, 5-diphenyltetrazolium bromide
- PBS:
-
phosphate-buffered saline (pH 7.4)
- PI3 kinase:
-
phosphatidylinositol 3′-kinase
- STAT:
-
signal transducer and activator of transcription
References
Tuynman JB, Peppelenbosch MP, Richel DJ (2004) COX-2 inhibition as a tool to treat and prevent colorectal cancer. Crit Rev Oncol Hematol 52:81–101
Giovannucci E, Ascherio A, Rimm EB, Colditz GA, Stampfer MJ, Willett WC (1995) Physical activity, obesity, and risk for colon cancer and adenoma in men. Ann Intern Med 122:327–334
Pan SY, Johnson KC, Ugnat AM, Wen SW, Mao Y (2004) Association of obesity and cancer risk in Canada. Am J Epidemiol 159: 259–268
Considine RV, Sinha MK, Heiman ML, Kriauciunas A, Stephens TW, Nyce MR, Ohannesian JP, Marco CC, McKee LJ, Bauer TL et al (1996) Serum immunoreactive-leptin concentrations in normal-weight and obese humans. N Engl J Med 334:292–295
Stattin P, Lukanova A, Biessy C, Soderberg S, Palmqvist R, Kaaks R, Olsson T, Jellum E (2004) Obesity and colon cancer: does leptin provide a link? Int J Cancer 109:149–152
Attoub S, Noe V, Pirola L, Bruyneel E, Chastre E, Mareel M, Wymann MP, Gespach C (2000) Leptin promotes invasiveness of kidney and colonic epithelial cells via phosphoinositide 3-kinase-, rho-, and rac-dependent signaling pathways. FASEB J 14:2329–2338
Hardwick JC, van Den Brink GR, Offerhaus GJ, van Deventer SJ, Peppelenbosch MP (2001) Leptin is a growth factor for colonic epithelial cells. Gastroenterology 121:79–90
Myers MG Jr (2004) Leptin receptor signaling and the regulation of mammalian physiology. Recent Prog Horm Res 59:287–304
Saxena NK, Titus MA, Ding X, Floyd J, Srinivasan S, Sitaraman SV, Anania FA (2004) Leptin as a novel profibrogenic cytokine in hepatic stellate cells: mitogenesis and inhibition of apoptosis mediated by extracellular regulated kinase (Erk) and Akt phosphorylation. FASEB J 18:1612–1614
Kim GS, Hong JS, Kim SW, Koh JM, An CS, Choi JY, Cheng SL (2003) Leptin induces apoptosis via ERK/cPLA2/cytochrome c pathway in human bone marrow stromal cells. J Biol Chem 278:21920–21929
Brunet A, Bonni A, Zigmond MJ, Lin MZ, Juo P, Hu LS, Anderson MJ, Arden KC, Blenis J, Greenberg ME (1999) Akt promotes cell survival by phosphorylating and inhibiting a Forkhead transcription factor. Cell 96:857–868
Thompson JE, Thompson CB (2004) Putting the rap on Akt. J Clin Oncol 22:4217–4226
Somasundar P, Frankenberry KA, Skinner H, Vedula G, McFadden DW, Riggs D, Jackson B, Vangilder R, Hileman SM, Vona-Davis LC (2004) Prostate cancer cell proliferation is influenced by leptin. J Surg Res 118:71–82
Stepan VM, Sawada M, Todisco A, Dickinson CJ (1999) Glycine-extended gastrin exerts growth-promoting effects on human colon cancer cells. Mol Med 5:147–159
Ogunwobi O, Beales I (2005) Glycine-extended gastrin stimulates proliferation and inhibits apoptosis in colon cancer cells via multiple pathways (abstract). Gut 54(Suppl 11):A57
Onuma M, Bub JD, Rummel TL, Iwamoto Y (2003) Prostate cancer cell–adipocyte interaction: leptin mediates androgen-independent prostate cancer cell proliferation through c-Jun NH2-terminal kinase. J Biol Chem 278:42660–42667
Sakai I, Kraft AS (1997) The kinase domain of Jak2 mediates induction of bcl-2 and delays cell death in hematopoietic cells. J Biol Chem 272:12350–12358
Kloek C, Haq AK, Dunn SL, Lavery HJ, Banks AS, Myers MG Jr (2002) Regulation of Jak kinases by intracellular leptin receptor sequences. J Biol Chem 277:41547–41555
Pai R, Lin C, Tran T, Tarnawski A (2005) Leptin activates STAT and ERK2 pathways and induces gastric cancer cell proliferation. Biochem Biophys Res Commun 331:984–992
Vaisse C, Halaas JL, Horvath CM, Darnell JE Jr, Stoffel M, Friedman JM (1996) Leptin activation of Stat3 in the hypothalamus of wild-type and ob/ob mice but not db/db mice. Nat Genet 14:95–97
Ma XT, Wang S, Ye YJ, Du RY, Cui ZR, Somsouk M (2004) Constitutive activation of Stat3 signaling pathway in human colorectal carcinoma. World J Gastroenterol 10:1569–1573
Steinbach G, Lynch PM, Phillips RK, Wallace MH, Hawk E, Gordon GB, Wakabayashi N, Saunders B, Shen Y, Fujimura T et al (2000) The effect of celecoxib, a cyclooxygenase-2 inhibitor, in familial adenomatous polyposis. N Engl J Med 342:1946–1952
Corpet DE, Pierre F (2005) How good are rodent models of carcinogenesis in predicting efficacy in humans? A systematic review and meta-analysis of colon chemoprevention in rats, mice and men. Eur J Cancer 41:1911–1922
Maier TJ, Schilling K, Schmidt R, Geisslinger G, Grosch S (2004) Cyclooxygenase-2 (COX-2)-dependent and -independent anticarcinogenic effects of celecoxib in human colon carcinoma cells. Biochem Pharmacol 67:1469–1478
Beales I, Calam J (1997) Stimulation of IL-8 production in human gastric epithelial cells by Helicobacter pylori, IL-1β and TNF-α requires tyrosine kinase activity, but not protein kinase C. Cytokine 9:514–520
Beales IL (2004) Gastrin and interleukin-1beta stimulate growth factor secretion from cultured rabbit gastric parietal cells. Life Sci 75:2983–2995
Micoud F, Mandrand B, Malcus-Vocanson C (2001) Comparison of several techniques for the detection of apoptotic astrocytes in vitro. Cell Prolif 34:99–113
Ogunwobi OO, Beales IL (2006) Glycine-extended gastrin stimulates proliferation and inhibits apoptosis in colon cancer cells via cyclo-oxygenase-independent pathways. Regul Pept 134:1–8
Ostlund RE Jr, Yang JW, Klein S, Gingerich R (1996) Relation between plasma leptin concentration and body fat, gender, diet, age, and metabolic covariates. J Clin Endocrinol Metab 81:3909–3913
Zimmet P, Hodge A, Nicolson M, Staten M, de Courten M, Moore J, Morawiecki A, Lubina J, Collier G, Alberti G et al (1996) Serum leptin concentration, obesity, and insulin resistance in Western Samoans: cross sectional study. BMJ 313:965–969
Datta SR, Dudek H, Tao X, Masters S, Fu H, Gotoh Y, Greenberg ME (1997) Akt phosphorylation of BAD couples survival signals to the cell-intrinsic death machinery. Cell 91:231–241
Skov S, Klausen P, Claesson MH (1997) Ligation of major histocompatibility complex (MHC) class I molecules on human T cells induces cell death through PI-3 kinase-induced c-Jun NH2-terminal kinase activity: a novel apoptotic pathway distinct from Fas-induced apoptosis. J Cell Biol 139:1523–1531
Bjorbaek C, Uotani S, da Silva B, Flier JS (1997) Divergent signaling capacities of the long and short isoforms of the leptin receptor. J Biol Chem 272:32686–32695
Arico S, Pattingre S, Bauvy C, Gane P, Barbat A, Codogno P, Ogier-Denis E (2002) Celecoxib induces apoptosis by inhibiting 3-phosphoinositide-dependent protein kinase-1 activity in the human colon cancer HT-29 cell line. J Biol Chem 277:27613–27621
Kumar NV, Bernstein LR (2001) Ten ERK-related proteins in three distinct classes associate with AP-1 proteins and/or AP-1 DNA. J Biol Chem 276:32362–32372
Zhong SP, Ma WY, Quealy JA, Zhang Y, Dong Z (2001) Organ-specific distribution of AP-1 in AP-1 luciferase transgenic mice during the maturation process. Am J Physiol Regul Integr Comp Physiol 280:R376–R381
Shaulian E, Karin M (2001) AP-1 in cell proliferation and survival. Oncogene 20:2390–2400
Ashida R, Tominaga K, Sasaki E, Watanabe T, Fujiwara Y, Oshitani N, Higuchi K, Mitsuyama S, Iwao H, Arakawa T (2005) AP-1 and colorectal cancer. Inflammopharmacology 13:113–125
Rivat C, Rodrigues S, Bruyneel E, Pietu G, Robert A, Redeuilh G, Bracke M, Gespach C, Attoub S (2005) Implication of STAT3 signaling in human colonic cancer cells during intestinal trefoil factor 3 (TFF3)—and vascular endothelial growth factor-mediated cellular invasion and tumor growth. Cancer Res 65:195–202
Bromberg J (2002) Stat proteins and oncogenesis. J Clin Invest 109:1139–1142
Rouet-Benzineb P, Aparicio T, Guilmeau S, Pouzet C, Descatoire V, Buyse M, Bado A (2004) Leptin counteracts sodium butyrate_induced apoptosis in human colon cancer HT-29 cells via NF-kappaB signaling. J Biol Chem 279:16495–16502
Acknowledgements
This work was funded by the Norfolk and Norwich University Hospital Bicentenary Trust in the form of a research studentship to Dr. Ogunwobi. Further financial support was provided by The Royal Society, The Peel Medical Research Trust, The Mason Medical Research Foundation, The Institute of Biomedical Science, the Norfolk and Norwich University Hospital Inflammatory Bowel Disease Research Fund and National Health Service (NHS) Research and Development. Part of this work was presented in abstract form to the British Society of Gastroenterology, March 2005.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Ogunwobi, O.O., Beales, I.L.P. The anti-apoptotic and growth stimulatory actions of leptin in human colon cancer cells involves activation of JNK mitogen activated protein kinase, JAK2 and PI3 kinase/Akt. Int J Colorectal Dis 22, 401–409 (2007). https://doi.org/10.1007/s00384-006-0181-y
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00384-006-0181-y