
Abstract
Background
The specificity of cerebrospinal fluid (CSF) dynamics in the immature brain still remains unknown. In our data previously published, the transependymal intraparenchymal CSF pathway (the minor pathway) plays a significant role in various degrees in the alternative CSF passage. Now, there is a growing consensus in the age differences in the outcome of neuroendoscopic ventriculostomy in treatment of non-communicating types of hydrocephalus. The authors discuss the clinical significance of the specific CSF dynamics and propose the new aspect of classification of hydrocephalus with a theory in the development of CSF from the pathophysiological point of view.
Patients and methods
Between January 2001 and March 2004, 122 hydrocephalic children were registered at the Jikei University Hospital Women’s & Children’s Medical Center (JWCMC), Tokyo. Our retrospective study for the efficacy of neuroendoscopic ventriculostomy confirmed the significantly high failure rate of neuroendoscopic ventriculostomy in treating hydrocephalus in neonates and infants with non-communicating hydrocephalus as the initial impression. The prospective CSF dynamic studies using cine-mode MRI and CT ventriculo-cisternography were then routinely started. Altogether, 9 out of 29 (31%) endoscopically treated cases needed shunt placement 3–30 weeks (mean 7.9 weeks) after the endoscopic procedure(s) (publication in preparation). Five out of 11 (45%) neonates/infants under 3 months, 3/5 (60%) infants at 7–12 months, 10/10 (100%) toddlers at 1–4 years and 3/3 (100%) schoolchildren at 5–17 years were cured, as in the condition of “post-endoscopic ventriculostomy arrested hydrocephalus”. The pattern of ventriculo-cisternography in neonatal/infantile cases revealed intraparenchymal predominant pattern (minor pathway) of the CSF dynamics rather than passage in the major pathway.
Discussion
The various basic investigations in rodents, cats and monkeys have suggested that CSF is absorbed not via Pacchionian bodies as the last end of the major pathway, which do not exist in these animals and are recognized after infantile period in human, but through the choroids plexus and the periventricular fenestrated venous capillaries into the deep venous channel. The high incidence of “failure to arrest hydrocephalus” by neuroendoscopic ventriculostomy in fetal, neonatal and infantile periods was considered to depend on the specific CSF dynamics, in which the major CSF pathway has not developed and the minor pathway has a significant role.
Proposal of theory
We herein propose a new aspect of classification for hydrocephalus with special reference to the CSF circulation in the minor CSF pathway, i.e. “minor pathway hydrocephalus”, differentiating the conventional classification by Dandy (communicating and non-communicating) or Russell (non-obstructive and obstructive) as “major pathway hydrocephalus”. We also herein propose a hypothesis that the CSF dynamics develop in the theory of evolution from the immature brain, as in the animals with the minor CSF pathway predominance, towards matured adult human brain together with completion of the major CSF pathway: the “evolution theory in CSF dynamics”.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
For over a century, the cerebrospinal fluid (CSF) dynamics have been extensively investigated as a challenging subject, both in basic research and in clinical analyses on pathophysiology (Fig. 1). Since the CSF circulation was considered as the third circulation of the body, the CSF dynamics have been considered in the circulatory passage, [31] the major and minor pathways. Hydrocephalus is defined as “disturbed CSF dynamics” or “disorder of harmony in CSF dynamics” [18] and classified as “communicating vs non-communicating” by Dandy and Blackfan (1914) [6] and “non-obstructive vs obstructive” by Russell (1949) [26]. These classical or traditional concepts have been supported as the “standard” in considering the pathophysiology of hydrocephalus in the individual case. However, one of the most controvertible issues in these concepts is the fact that during the developmental process in the neonatal and infantile periods, the CSF dynamics are not identical to the completed pattern as in the adult. Especially, the embryogenesis or morphological/functional development of the CSF dynamics is quite obscure. Since the details of “normal” CSF dynamics in the developmental process are unclear, the treatment goal for hydrocephalus, as the “normalization of the disturbed CSF circulation”, remains absolutely irresolute as the future subject to be solved in fetal, neonatal and infantile hydrocephalus. The ongoing discussion, shunt vs neuroendoscopic (third) ventriculostomy as the surgical choice in treatment for hydrocephalus affecting the immature brain, has not come to a conclusion yet [14, 22]. Since the balance of the individual role and function of the major/minor CSF pathway is totally unknown in the developmental process of the immature brain, the goal of the treatment for the individual hydrocephalus case varies along with the age of the patient and the type of the disturbed CSF circulation.

Historical trend of research on CSF physiology and treatment for hydrocephalus in the twentieth century
It has been pointed out that the success rate in treatment for hydrocephalus in the neonatal and early infantile period by the neuroendoscopic ventriculostomy is significantly low, i.e. down to 35% [14]. There has been neither definitive explanation nor evidence for the pathophysiology of the failure of ventriculostomy in treating hydrocephalus involving the immature brain. On the basis of previously published clinical data and review of the experimental studies on the CSF dynamics in animals, the authors herein propose a new aspect in the classification of hydrocephalus with a hypothesis, the “evolution theory in CSF dynamics”, in the development of CSF circulation during the fetal, neonatal and early infantile period.
Traditional concept of CSF dynamics
The CSF dynamics are considered in two main mechanical passages: major and minor pathways. The major pathway starts from the bilateral lateral ventricles with choroid plexus as the significant CSF production source merging with CSF produced in the third and fourth ventricles, and CSF passes outside the ventricular system into the cisterns or subarachnoid space. The major absorption site is the arachnoid granulation (Pacchionian body) or villi into the sinus, mainly superior sagittal sinus (Weed 1914) [30]. The minor pathway includes the routes via the ventricular ependyma, interstitial and perivascular space, and perineural lymphatic channel. The absorption sites are located in various pia-arachnoid capillaries/leptomeninges and even in choroid plexus (Table 1). With the bidirectional volume movement of CSF in the major pathway, the CSF dynamics created the bulk flow (Dandy and Blackfan 1914) [6]. The rate of CSF production is approximately 500 ml over 24 h in humans, and because the CSF major pathway is some 130–140 ml, there may be physiologic turnover of the CSF three or four times every day. It is well known that an appreciable volume of CSF comes from sources other than choroid plexus in animals [3, 27, 28]. Communication between CSF in the major pathway and interstitial fluid (ISF) in the minor pathway plays an important role in maintaining homeostasis of the brain, keeping the microchemical environment in the physiological balance. The total amount of the CSF production when ISF is included is 330 ml/day [27].
Classical classification of hydrocephalus
Based on this traditional concept of CSF dynamics, the hydrocephalus has been defined as a state of “disturbed CSF circulation” [18] and classified classically into two types, communicating and non-communicating [5]. In the definition of Dandy’s communicating/non-communicating hydrocephalus, the communication of the CSF pathway is between the lateral ventricle and the lumbar subarachnoid space (confirmed by injection of dye into the lateral ventricle and detection by lumbar puncture) (Fig. 2). The terminology with obstructive/non-obstructive hydrocephalus [26], however, is not identical. The obstruction in the definition is at any region in the major CSF pathway including the ventricular system and entire cistern/subarachnoid space, so that the cause or condition for non-obstructive hydrocephalus is limited to either CSF overproduction by choroid plexus papilloma or CSF malabsorption due to sinus thrombosis (Fig. 3). Although there are numerous parameters for classification of hydrocephalus and the full range of classified subtypes of hydrocephalus is uncountable [17], the minor CSF pathway has not been considered as the direct cause for or any subtype of the classification of hydrocephalus.

Classical classification of hydrocephalus based on the “disturbed CSF dynamics” in the twentieth century #1

Classical classification of hydrocephalus based on the “disturbed CSF dynamics” in the twentieth century #2
Various forms of hydrocephalus and its specific CSF dynamics
Considering the most indicated treatment modality for and estimated prognosis of hydrocephalus, the classification of hydrocephalus becomes an important issue. The pathophysiological condition of disturbed CSF dynamics with a variety of underlying diseases may include numerous categories, parameters and criteria for classification [17]. The most crucial criterion of indication for neuroendoscopic ventriculostomy is the specific pattern of the disturbed CSF dynamics causing hydrocephalus in individual cases [19, 22]. Therefore, taking the specificity as the parameter of the classification, “communicating vs non-communicating” by Dandy [5] may still be the most proper among the presently available classifications. Since the basic concept in this classification is in the site of occlusion in the major CF pathway, especially the outlet of the fourth ventricle or intraventricular passage, the interact draining route after the individual procedure of ventriculostomy shall be the key to normalizing the disturbed CSF dynamics. The classification of “non-obstructive vs obstructive” by Russell [26] shall not be suitable in this sense. Actually, obstructive hydrocephalus can occur not only by occlusion within the ventricular system, but also in any location where the CSF flow in the subarachnoid space is blocked before being absorbed via the arachnoid granulation/villi (Pacchionian body).
These considerations are based on the concept of classifying the type of hydrocephalus by taking the disturbed CSF dynamics only in the major CSF pathway. The authors herein define the concept of these classifications as “major pathway hydrocephalus”.
Ontogenesis of CSF dynamics
Along with the neuronal maturation process in the central nervous system (CNS) (Fig. 4), the pathway of CSF circulation starts to develop, first forming the choroid plexus in the primitive lumen as the ventricle (Fig. 5). The first histological evidence of the choroidal differentiation is described as 41–44 embryonic days in the fourth ventricle, 44 days in the lateral and 57 days in the third ventricle. Although it was the findings for the pig embryo, Weed [31] defined that the roof of the fourth ventricle (area membrane inferiors; AMI) opens up at 8 weeks of gestational age immediately after the choroidal differentiation in the fourth ventricle is observed [12]. Osaka et al. [23] reported the formation of the subarachnoid space in as early as the fourth embryonic week in humans, which did not continue to the fourth ventricular opening. They also suggested that the beginning of the CSF formation is not likely a main force that expands and creates the subarachnoid space in the concept of the major CSF pathway in adults. According to Lemire et al. (1975) [15], the meninx primitiva begins to cogitate at 8 weeks, and the arachnoid is first indicated at 12 weeks and begins to separate from the dura mater at 20 weeks of gestational age (Fig. 6).

Neuronal maturation stages in human and rat. With permission by Oi et al. [20]

A rat fetus model of congenital hydrocephalus inducted by amino-nicotinamide injection on day 13 gestational age. Note the development of the developing choroid plexus and subarachnoid space normal control (right upper figures) and massive dilatation of the ventricle/central canal and subarachnoid space in the hydrocephalus model. With permission by Oi et al. [20]

CSF dynamics maturation (CSFDM) stages I–V in human
Regarding the CSF absorption sites, it is well known that there is no arachnoid granulation (Pacchionian body) in mouse, rat, rabbit and cat, even in their adulthood [11]. In large mammals and humans, the arachnoid granulation (Pacchionian body) appears in postnatal life or just before birth microscopically as villi [9, 11] and begins to function as the CSF reabsorption route in later age [29] (Fig. 7). Since the arachnoid villi are microscopic structures whereas granulations are gross prominent anatomical finding, the development of arachnoid granulations (Pacchionian body) has been used as a criterion of age in the radiological identification with a finding of parasagittal depressions of the calvarium [1]. The radiological evidence of the presence of arachnoid granulation is usually obtained first at 7 years and progressively develops up to 20 years of age [10]. The CSF reabsorption function by the arachnoid graduation may be obtained in late infantile period [29]. Without the expected functional development of the arachnoid graduation, the CSF dynamics may be maintained at the minor pathway with the drainage route via perineural space to the lymphatic system [4, 7, 8, 16, 25] (Fig. 8), via transependymal–interstitial to the perivascular/subpial space both in the brain and spinal cord [2, 12, 13], and via epithelium of the choroid plexus to the fenestrated capillaries and finally to the galenic venous system [11]. This “minor CSF pathway” is the main route for the CSF dynamics both in rodents or small mammals [2, 7, 16, 25] (Fig. 7) and developing immature brain in humans [24, 29] (Fig. 9).

Ontogenesis of arachnoid villi/granulation (Pacchionian body)

CSF dynamics in cat. The water-soluble contrast injected in the ventricle in an adult cat. Courtesy of Wolf Luedemann [16]

Minor CSF pathway in immature brain
Clinical significance and proposal of a theory
Between January 2001 and March 2004, 122 hydrocephalic children were registered to the Jikei University Hospital Women’s & Children’s Medical Center (JWCMC), Tokyo. Our retrospective study for the efficacy of neuroendoscopic ventriculostomy confirmed the significantly high failure rate of neuroendoscopic ventriculostomy in treating hydrocephalus in neonates and infants with non-communicating hydrocephalus hydrocephalus as the initial impression. The prospective CSF dynamic studies using cine-mode MRI and CT ventriculo-cisternography were then routinely started. Altogether, 9 out of 29 (31%) endoscopically treated cases needed shunt placement 3–30 weeks (mean 7.9 weeks) after the endoscopic procedure(s) (publication in preparation). Five out of 11 (45%) neonates/infants under 3 months, 3/5 (60%) infants at 7–12 months, 10/10 (100%) toddlers at 1–4 years and 3/3 (100%) schoolchildren at 5–17 years were cured, as in the condition of “post-endoscopic ventriculostomy arrested hydrocephalus”. The pattern of ventriculo-cisternography in neonatal/infantile cases revealed intraparenchymal predominant pattern (minor pathway) of the CSF dynamics rather than passage in the major pathway.
Since the CSF dynamics during the fetal and neonatal/early infantile periods are mainly maintained in the minor CSF pathway with premature development of superior sagittal sinus (Fig. 10), hydrocephalus occurring during these periods shall be defined as disturbed CSF circulation in the minor CSF pathway (minor CSF pathway hydrocephalus). The causative underlying conditions may include various pathologies, as in the perspective classification of congenital hydrocephalus (PCCH; Oi et al. 1998 [21]), i.e. primary dysgenetic or secondary such as intraventricular hemorrhage (IVH) in the fetal brain. Hydrocephalus occurring in PCCH stage I (before 22 weeks of gestational age), stage II (22–32 weeks), stage III (33 weeks; full term) or stage IV (neonatal period) is most likely defined as minor CSF pathway hydrocephalus (Fig. 11). The pathophysiology with such specific CSF dynamics of minor CSF pathway hydrocephalus is most likely closely related to the cause of failure of neuroendoscopic ventriculostomy along with the maturation in the development process of the “major CSF pathway” during the infancy after neonatal period.

A 7-month-old boy with Apert syndrome. The intraoperative observation confirming the immature superior sagittal sinus (SSS)

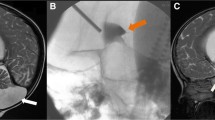
A 4-day-old girl with fetal IVH and hydrocephalus. The primary impression for occlusive site was the outlets of the IV ventricle. Neuroendoscopic procedure confirmed aqueductal occlusion, which was treated by aqueductal plasty and third-ventriculostomy (ETV). The postoperative ventriculo-cisternostomy of the CSF dynamics (4 days after injection of water-soluble contrast) demonstrated prolonged contrast stasis in all communicating ventricles and subarachnoid space. The contrast is more densely staying in the brain parenchyma through the ventricular wall (ependyma). This condition can be defined as “post-ETV communicating hydrocephalus” in the classical classification (major CSF pathway hydrocephalus) with aqueductal occlusion, which was treated by aqueductal plasty and ETV, but more than that, it suggests minor CSF pathway hydrocephalus
The indication for third ventriculostomy may gradually develop during infancy before 1 year of age to non-communicating type of hydrocephalus as “major CSF pathway hydrocephalus”, i.e. PCCH Stage V (infantile period up to 1 year of age after neonatal period).
This clinical evidence may be based upon the specific developmental process of CSF dynamics in human, which mimics the individual matured stage in the various species of animals.
We herein propose this clinically extremely significant aspect, namely, the “evolution theory in CSF dynamics”.
References
Basmajian JV (1952) The depressions for the arachnoid granulations as a criterion of age. Anat Rec 112(4):843–846
Becker DP, Wilson JA, Watson GW (1972) The spinal cord central canal: response to experimental hydrocephalus and canal occlusion. J Neurosurg 36(4):416–424
Bering EA Jr, Sato O (1963) Hydrocephalus: changes in formation and absorption of cerebrospinal fluid within the cerebral ventricles. J Neurosurg 20:1050–1063
Boulton M, Flessner M, Armstrong D, Hay J, Johnston M (1998) Determination of volumetric cerebrospinal fluid absorption into extracranial lymphatics in sheep. Am J Physiol 274(1 Pt 2):R88–R96
Dandy WE (1919) Experimental hydrocephalus. Ann Surg 70:129–142
Dandy WE, Blackfan KD (1914) Internal hydrocephalus. An experimental, clinical and pathological study. Am J Dis Child 8:406–482
Dohrmann GJ (1972) Cervical spinal cord in experimental hydrocephalus. J Neurosurg 37(5):538–542
Field EJ, Brierley JB (1948) The lymphatic connections of the subarachnoid space. Br Med J 1:1167–1171
Gomez DG, DiBenedetto AT, Pavese AM, Firpo A, Hershan DB, Potts DG (1982) Development of arachnoid villi and granulations in man. Acta Anat (Basel) 111(3):247–258
Grossman CB, Potts DG (1974) Arachnoid granulations: radiology and anatomy. Radiology 113(1):95–100
Hashimoto PH (2004) The cerebrospinal fluid as a tissue fluid of the nervous system? The route of CSF circulation and its clinical significance. Nervous System in Children (Shoni no Noshinkei) 29:217–223 (JPN)
Hochwald GM, Boal RD, Marlin AE, Kumar AJ (1975) Changes in regional blood-flow and water content of brain and spinal cord in acute and chronic experimental hydrocephalus. Dev Med Child Neurol Suppl 35:42–50
Katzman R, Schimmel H, Wilson CE (1968) Diffusion of inulin as a measure of extracellular fluid space in brain. Proc Rudolf Virchow Med Soc City N Y 26(Suppl):254–280
Koch D, Wagner W (2004) Endoscopic third ventriculostomy in infants of less than 1 year of age: which factors influence the outcome? Childs Nerv Syst 20(6):405–411
Lemire JR, Loeser JD, Leach RW, Alvord EC Jr (1975) Normal and abnormal development of the human nervous system. Parper & Row, Hagerstown, pp 283
Luedemann W, von Rautenfeld DB, Samii M, Brinker T (2005) Ultrastructure of the cerebrospinal fluid outflow along the optic nerve into the lymphatic system. Childs Nerv Syst 21:96–103
Oi S (2004) Classification and definition of hydrocephalus—origin, controversy and assignment of the terminology. In: Cinalli G, Sainte-Rose C, Maixner W (eds) Pediatric hydrocephalus book, chapter 6. Springer, Berlin Heidelberg New York, pp 95–112
Oi S (2004) Development in harmony. Childs Nerv Syst 20:693–701
Oi S, Kudo H, Yamada H, Kim S, Hamano S, Urui S, Matsumoto S (1991) Hydromyelic hydrocephalus. Correlation of hydromyelia with various stages of hydrocephalus in postshunt isolated compartments. J Neurosurg 74(3):371–379
Oi S, Yamada H, Sato O, Matsumoto S (1996) Experimental models of congenital hydrocephalus and comparable clinical problems in the fetal and neonatal periods. Childs Nerv Syst 12(6):292–302
Oi S, Honda Y, Hidaka M, Sato O, Matsumoto S (1998) Intrauterine high-resolution magnetic resonance imaging in fetal hydrocephalus and prenatal estimation of postnatal outcomes with “perspective classification”. J Neurosurg 88:685–694
Oi S, Hidaka M, Honda Y, Togo K, Shinoda M, Shimoda M, Tsugane R, Sato O (1999) Neuroendoscopic surgery for specific forms of hydrocephalus. Childs Nerv Syst 15(1):56–68
Osaka K, Handa H, Matsumoto S, Yasuda M (1980) Development of the cerebrospinal fluid pathway in the normal and abnormal human embryos. Childs Brain 6(1):26–38
Pappenheimer JR, Hesey SR, Jordan EF (1961) Active transport of Diodrast and phenolsulfonphthalein from cerebrospinal fluid to blood. Am J Physiol 200:1–10
Potts DG, Deonarine V, Welton W (1972) Perfusion studies of the cerebrospinal fluid absorptive pathways in the dog. Radiology 104(2):321–325
Russell DS (1949) Observation on the pathology of hydrocephalus. Medical research council. Special report series No. 265. His Majesty’s Stationery Office, London, pp 112–113
Sato O, Asai T, Amano Y, Hara M, Tsugane R, Yagi M (1972) Extraventricular origin of the cerebrospinal fluid: formation rate quantitatively measured in the spinal subarachnoid space of dogs. J Neurosurg 36(3):276–282
Sato O, Bering EA Jr, Yagi M, Tsugane R, Hara M, Amano Y, Asai T (1975) Bulk flow in the cerebrospinal fluid system of the dog. Acta Neurol Scand 51(1):1–11
Turner L (1961) The structure of arachnoid granulations with observations on their physiological and pathological significance. Ann R Coll Surg Engl 29:237–264
Weed LH (1914) Studies on cerebrospinal fluid. III. The pathways of escape from the subarachnoid spaces with particular reference to the arachnoid villi. J Med Res 31:51–91
Weed LH (1916) The establishment of the circulation of cerebro-spinal fluid. Anat Rec 10:256–258
Weed LH (1916) The formation of the cranial subarachnoid spaces. Anat Rec 10:475–481
Acknowledgements
The authors are grateful to Ms. Noriko Ogawa for her excellent type/technical works. This study was supported by the Kawano Masanori Memorial Foundation for Promotion of Pediatrics and Grant-in-Aid for Scientific Research (B)15659344, The Ministry of Education, Science, Supports and Culture. C.D.R. had been invited as a visiting professor to the Division of Neurosurgery, The Jikei University School of Medicine, Tokyo.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Oi, S., Di Rocco, C. Proposal of “evolution theory in cerebrospinal fluid dynamics” and minor pathway hydrocephalus in developing immature brain. Childs Nerv Syst 22, 662–669 (2006). https://doi.org/10.1007/s00381-005-0020-4
Received:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00381-005-0020-4