
Abstract
Information is needed on the ability of different crop management factors to maintain or increase soil C and N pools, especially in intensively tilled short crop rotations. Soil samples from field experiments in Maine were used to assess the effect of cover crop, green manure (GM) crop, and intermittent or annual amendment on soil C and N pools. These field experiments, of 6–13 years’ duration, were all characterized by a 2-year rotation with either sweet corn ( Zea mays L.) or potato ( Solanum tuberosum L.), and primary tillage each year. Total, particulate organic matter (POM), and soil microbial biomass (SMB)-C and -N pools were assessed for each experiment. Total C and N stocks were not affected by red clover ( Trifolium pratense L.) cover crop or legume GM, but were increased by 25–53% via a single application of papermill sludge or an annual manure and/or compost amendment. With the exception of continuous potato production which dramatically reduced the SMB-C and SMB-N concentration, SMB-C and -N were minimally affected by changes in cropping sequence, but were quite sensitive to amendments, even those that were primarily C. POM-C and -N, associated with the coarse mineral fraction (53–2,000 µm), were more responsive to management factors compared to total C and N in soil. The change in soil C fractions was a linear function of increasing C supply, across all experiments and treatments. Within these intensively tilled, 2-year crop rotations, substantial C and N inputs from amendments are needed to significantly alter soil C and N pools, although cropping sequence changes can influence more labile pools responsible for nutrient cycling.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Soil C and N stocks, and the distribution of C and N into pools or fractions of varying degradability, are altered by annual tillage operations that characterize many crop production systems. These types of changes can be attributed to both the increased mineralization that occurs as the frequency and intensity of tillage increases, (Beare et al. 1994a, 1994b; Haynes and Tregurtha 1999; Hussain et al. 1999), and by lesser amounts of C and N incorporated into the soil as crop residue or applied amendments, including animal manures. Labile C and N associated with the coarse mineral fraction (i.e., particulate organic matter; POM) and soil microbial biomass (SMB)-C and -N are noticeably impacted, while gross measurements like total soil C and N concentration may respond more slowly (Franzluebbers and Arshad 1996; Franzluebbers et al. 1999b). These changes can be dramatic for production systems that are characterized by frequent aggressive tillage operations and short rotations with low-residue crops, both of which describe the potato production system in northeastern USA and eastern Canada (Carter and Sanderson 2001).
There are a range of options available to attempt to remedy these detrimental soil effects, even in situations where frequent annual tillage continues. Without altering the crop rotation sequence, residue can be increased by inter- or relay-cropping with annual cover crops. The impact of nitrogenous cover crop residues on POM-C or POM-N and on SMB are generally transient in nature, not leading to increased soil organic matter (SOM) (Hu et al. 1997; Jackson 2000; Mendes et al. 1999). Alternatively, crop selection may be based on increasing the amount of residue remaining after harvest, or the entire plant biomass may be incorporated (green manure; GM) instead of harvesting. These approaches have the potential to add modest amounts of C (0.25–2 Mg ha-1) and N (20–200 kg ha-1) (Griffin and Hesterman 1991; McVay et al. 1989), a large portion of which may be rapidly mineralized rather than being converted to more stable forms (Quemada and Cabrera 1995).
The other approach used to increase C and N is to apply amendments, in the form of animal manures, compost, or other industrial byproducts. Such amendments can supply significantly greater amounts of C to soil than changes in cropping sequence (Aoyama et al. 1999; Drinkwater et al. 1998), ranging up to 8–12 Mg C ha-1 even when applied on N-rate bases. Rapid increases in total soil C and N and more labile POM pools have been accomplished via the annual or intermittent application of such materials (Chatigny et al. 1999; Gallandt et al. 1998; Rochette et al. 2000).
Four medium-term (6–13 years’ duration) field research projects conducted in Maine which began in 1987 offer the opportunity to evaluate the impacts of crop rotation and amendment options on soil C and N pools. Three of these experiments focused on potato ( Solanum tuberosum L.) production in northern Maine and one evaluated common bean ( Phaeseolus vulgaris L.) and sweet corn ( Zea mays L.) production in central Maine. All experiments are located on sandy loam soils derived from glacial till and include primary and secondary tillage every year. Soil samples obtained 6–13 years after the initiation of each experiment were used to assess the relative impacts of legume cover crop and GM, and periodic and annual compost and/or manure amendment on selected soil C and N pools.
Materials and methods
Field experiments
Four different research projects were utilized to obtain soil samples for this research, representing six distinct field experiments. Two projects (clover and potato GM) comprised separate experiments for each rotation entry point, initiating 2-year rotations in consecutive years. Each experiment is described briefly below.
The Clover experiment was initiated on adjacent field plots in 1993 and 1994 (i.e., two entry points) in Orono, Maine, on a Nicholville very fine sandy loam (coarse-silty, mixed, frigid, Aquic Haplorthods). The main experimental factor in this experiment was a small grain-test crop rotation, with and without a red clover ( Trifolium pratense L.) cover crop interseeded in the first year of the rotation. There was also a single application of compost to each plot containing red clover, immediately prior to the 1996 and 1997 test crop. This amendment contained 7.1 and 3.2 Mg C ha-1 in 1996 and 1997, respectively. The original rotation sequence (1993–1995) was wheat ( Triticum aestivum L.) with and without red clover, followed by dry bean as the test crop, and included additional treatment factors to evaluate weed interference and N nutrition. Subsequently, sweet corn was used as the test crop. These two phases are described by Liebman and Gallandt (2002) and Davis (1998). Tillage was performed in the spring, usually by rototilling, and wheat residue and clover biomass were sampled immediately before destruction in most years. All treatment factors except clover intercropping were discontinued after the 1997 and 1998 sweet corn crops for the two entry points. Ten field replicates of each treatment were sampled in May 2000, under the assumption that the long-term residual effects of weed inclusion on soil properties were minimal. Fifteen soil cores (2.5 cm diameter by 20 cm deep) were taken in each plot. These cores were mixed thoroughly and dried in a forced-air oven at 40°C for 3 days.
The most widely implemented potato rotation sequence in Maine is a 2-year, oat ( Avena sativa L.)- or barley ( Hordeum vulgare L.)-potato rotation, with primary tillage occurring in the fall and secondary tillage occurring in the spring, just prior to planting. The following three research efforts all contain this industry standard or status quo treatment, and offer comparisons of different practices to the status quo. All were conducted at the Maine Agricultural and Forest Experiment Station farm in Presque Isle, Maine, on Caribou gravelly loam soil (fine-loamy, isotic, frigid, Typic Haplorthods). The first experiment, called Potato GM, is a simple comparison between oat-potato and legume GM-potato rotations. This experiment, which contained additional treatments not evaluated here, was initiated in 1990 and 1991 (two entry points representing separate experiments, as with clover). The oat-potato rotation has been constant since initiation; the GM crop was crimson clover ( Trifolium incarnatum L.) through 1993 and a mixture of peas ( Pisum sativum L.), hairy vetch ( Vicia villosa Roth), and oat since 1994, as described by Plotkin (2000). Oat and potato were managed following common practices for the area, including fertilization, liming, and pest control, and the GM crops did not received any fertilizer or weed control. Soil samples used here were taken in May 2000, as described above.
The second potato rotation experiment, called Potato Mixed, contains two management alternatives, relative to the status quo oat-potato rotation treatment that has been in place since 1987. These include a continuous potato treatment (since 1995, previously in a 2-year rotation), and an oat-potato rotation which received a single amendment application. The amended treatment received primary papermill sludge (45 Mg ha-1, dry weight) in fall 1995, and cull potato compost (6.5 Mg ha-1, dry weight) in spring 1996. This amendment contained 14.0 Mg C ha-1 and 170 kg N ha-1. Soil samples were taken from all entry points in May 2000.
The final potato experiment, designated Eco, is the potato ecosystem experiment. This large, interdisciplinary experiment was initiated in 1990. It compares two soil management systems (unamended or amended with compost and manure) and three pest management systems (conventional, reduced pesticide, and biological) in factorial combination, and in the context of 2-year rotations. The treatment structure is described in detail by Alford et al. (1996) and Gallandt et al. (1998). The amended plots received an annual application of potato waste compost (10 Mg ha-1, dry weight) and beef manure (22 Mg ha-1, dry weight), containing on average 8.1 Mg C ha-1 and 278 kg organic N ha-1. Soil samples were collected in May 1999. Although all rotation entry points occur each year, samples were collected only from plots that contained potato in 1998, and are thus an evaluation of the cumulative effect of four rotation cycles with contrasting amendment treatments over an 8-year period.
To investigate the impact of C supply on total, POM-, and SMB-C pools, we quantified the difference in C input between the status quo rotation and the alternative treatment(s) in each experiment. We used published and unpublished data sources, as indicated for each experiment above. This approach does not require a quantitative, long-term mass balance for C, for which there are insufficient data. Rather, only estimates of crop residue differences (e.g., oat straw versus GM) and amendment nutrient concentration and application rate were required. These data are mostly available, as these treatments were the primary foci of the experiments discussed here. For Clover and Potato GM, there were sufficient data to estimate C input for each of the two experiments. To standardize these inputs, we calculated an “annualized” difference in C input, equal to the estimated total difference in C input, divided by the number of years since the implementation of that treatment. Similarly, we calculated the change in each soil C pool due to the change in management (again, compared to the status quo rotation).
Evaluation of soil C and N pools
Soil (0.5–2 kg) from each plot was sieved (2 mm) either field moist or after air drying with forced air at 40°C. Samples were then archived in cardboard boxes until analyses were initiated. Samples were thoroughly mixed prior to obtaining subsamples for the determinations described below. A subsample (1–2 g) was pulverized in a Wig-L-Bug amalgamator/mixer (Cresent, Elgin, Ill.) as a preparatory step to total C and N determination. Total C and N were determined by dry combustion, using a Carlo Erba NC2500 (ThermoQuest Italia, Rodano, Italy) elemental analyzer.
POM-C and -N fractionation were determined following the methods described by Cambardella and Elliott (1992), with some minor modifications. A 5.00-g dry soil sample was dispersed in 15 ml of 0.5% hexametaphosphate solution, for 16 h on a reciprocal shaker. The soil slurry was poured first through a 500-µm screen and then through a 53-µm screen. This allowed for separation into coarse and fine sand fractions, respectively. The materials retained on the sieves was rinsed thoroughly with deionized water, and transferred quantitatively to a pre-weighed aluminum weighing crucibles. Samples were dried at 50°C overnight and weighed. Each fraction was then pulverized. Rather than determining the POM-C and POM-N concentrations by difference between whole soil and the silt plus clay fraction (<53 µm), as described by Cambardella and Elliott (1992), C and N for each fraction were determined directly by dry combustion of the residue. The concentration of C and N in the residue and as a proportion of total C or N were estimated.
SMB was estimated following the microwave irradiation procedure of Islam and Weil (1999) . Duplicate samples of 20 g dry soil from each plot were placed in small glass beakers, re-wetted to 60% water filled pore space (WFPS), and pre-incubated for 10 days. After the preincubation, one sample was then allowed to continue incubating for 10 additional days. The other sample (irradiated) was first subsampled (2.5 g, which was extracted in 2 M KCl) to determine the initial inorganic N concentration ( N i) for SMB-N. The sample was then wetted to 80% (WFPS) and irradiated in a microwave oven, receiving a total of 800 J g-1 dry soil in two 400-J bursts. The sample was stirred and allowed to cool after each irradiation. The irradiated soil was then inoculated with 1.0 g untreated soil, packed to original density, and wetted to 65% WFPS. Both the control and irradiated samples were enclosed in 2-l canning jars with 5.0 ml of 1 M NaOH (as the CO2 trap), and incubated for an additional 10 days. After this incubation period, the soil was dried at 65°C, and N i was determined as above for both control and irradiated soils. The CO2 trap was terminated with 1.5 M BaCl2, and titrated with standardized 1 M HCl.
SMB-C was calculated without subtracting the control soil, as in Franzluebbers et al. (1999a) and Voroney and Paul (1984):
where K c equals 0.45 (Jenkinson and Powlson 1976). SMB-N was calculated following Voroney and Paul (1984):
where C flush and N flush are the C and N evolved during the 10-day incubation of the irradiated soil.
In some cases, large differences in SMB-C existed between treatments, so we also calculated specific respiratory activity (SRA; mg CO2 evolved mg-1 SMB-C day-1), to account for the difference in pool size, as
Statistics were applied to each experiment individually, with Clover and Potato GM both comprising two separate, adjacent experiments. Where significant treatment effects were identified in an experiment at a probability level of 5% with ANOVA, a least significant difference was calculated to separate treatment means.
Results and discussion
Total C and N in soil
The inclusion of a legume cover crop (Clover) or legume GM (Potato GM) did not significantly alter the total C or N concentration in the soil (Table 1). This was not surprising as the total stocks of C and N in the soil are of sufficient size that the differences in C or N input, compared to the status quo cropping system, are small. For example, assuming a 0.2-m plow-depth and a soil bulk density of 1.2 Mg m-3, a soil C concentration of 20.0 g kg-1 soil is equivalent to a C pool of 48,000 kg C ha-1, with an associated total N pool of approximately 4,300 kg ha-1. To alter this pool by 10% would require stabilized C and N pools to increase by 4,800 and 430 kg ha-1, respectively. For comparative purposes, using data from Davis (1998) and Liebman and Gallandt (2002), the difference in C input for the two treatments in clover is approximately 900 kg ha-1 year-1, from the incorporation of the red clover cover crop. This points out the difficulty in altering total C and N stocks in soil with relatively minor changes in the cropping pattern, especially when using crops with easily degradable residues (such as legumes), where the mineralization of plant-derived C and N may be as high as 80%. Previous reports from long-term experiments are mixed. For example, Collins et al. (1992) compared continuous wheat and wheat-pea rotations, in place for 25 years, and found that total soil C and N were not affected, presumably because the residue inputs were similar for both systems. In potato systems, Angers et al. (1999) found that after 9 years, soil C and N were similar in barley-potato and ryegrass ( Lolium perenne L.)-potato rotations.
Several changes in the total C and N concentration are apparent in the Potato Mixed experiment. Focusing first on results from potato entry points (i.e., soil samples obtained after the potato crop), the total C and N concentration was the same after 5 years in continuous potato as in the oat-potato systems (Table 1). Although there are well-documented benefits associated with the production of potato in rotation, due primarily to the reduced incidence and/or severity of disease, there are no notable differences in the ability of short rotations to increase soil C and N over this short a period. The amended treatment in Potato Mixed, however, increased the soil C concentration by about 25%, compared to the monoculture and the 2-year rotation. Chantigny et al. (1999) showed a similar multi-year effect, with increased soil C resulting from a single application of de-inking paper sludge at rates of 50 and 100 Mg ha-1 (dry weight; C concentration of 387 g kg-1 dry matter). Gagnon et al. (2001) also applied pulp fiber residue (at 45 and 90 Mg ha-1) and monitored soil C for 30 months after application. They found that soil C was increased by 8–10 g C kg-1 soil by these applications, which contained three- to fivefold more C than was applied in our research.
The Potato Mixed experiment is the only one where the effect of the previous crop on soil parameters can be evaluated directly, by comparing entry points of the oat-potato and amended oat-potato systems. There was a significant effect due to the previous crop in this experiment. The oat-potato rotation had 3.4 g more C kg-1 soil and 0.26 g more N kg-1 soil after the oat phase than after the potato phase, when no amendment was applied. Moore et al. (2000) also found that the total C and N concentration was affected by the previous crop in long-term experiments, but primarily in longer (4 year) rotations that contained at least 2 consecutive years of perennial forage crops, similar to the results of Bremer et al. (1994). In the amended system, there were no differences due to the previous crop. Overall, the impact of a single amendment applied 5 years previously is still apparent even with these relatively insensitive indicators.
An annual application of manure and compost, as done in the Eco experiment, increased the total C and N concentration by up to 53% and 43%, respectively, compared to the conventionally managed, 2-year barley-potato rotation that characterizes potato production in this area (Table 1). The trajectory of the increase in soil C through the first 6 years of this project is shown in Gallandt et al. (1998), with a significant increase in soil C (or SOM) beginning in the second year. In different experiments at the same location, Porter et al. (1999) showed that the application of potato waste compost plus manure increased the soil C concentration by 22% with a single application and 32% (approximately 6 g C kg-1 soil) with three successive annual applications.
POM-C and -N
POM represents between 18% and 40% of total C in these soils, depending on the experiment and treatment (Table 2). Between 60% and 80% of the POM-C is associated with fine sand particles (53–500 µm), and the remainder is associated with larger sand grains (500–2,000 µm).
Including legumes as a cover crop or GM crop (Clover and Potato GM, respectively) did not significantly affect the C concentration in the POM residue or POM-C as a proportion of total C in the soil (Table 2). In the Potato Mixed experiment, each POM-C fraction was enriched by the single C-rich amendment, compared to either the potato monoculture or the oat-potato rotation, which were similar. In some cases (e.g., coarse sand POM), this enrichment was >100%. These data demonstrate that it is the increased C supply that affects C fractions, and the comparison of the amended and unamended oat-potato rotations supports this interpretation. It is unlikely that differences arose from reduced C mineralization, as the frequency, intensity and timing of tillage were identical for all treatments. The amended oat-potato rotation also has a greater proportion of total C residing in the POM-C fractions, indicating that the residual C from prior amendment is not yet as humified as resident soil C. These are similar to findings of Gagnon et al. (2001), who showed that the application of pulp fiber residue increased macro-organic matter C (equivalent to POM-C described here) from 30% to 45–54% of total C, and increased POM-N from 10% to 15–17% of total N.
As expected, the annual application of manure and compost in Eco approximately doubles the C concentration in fine, coarse, and total POM-C fractions. Because fresh C is applied annually, the POM-C pool is differentially enriched. For example, POM-C makes up about 25% of total C in the unamended barley-potato rotation under conventional pest management; with annual amendment, this pool makes up nearly 40% of total soil C. Willson et al. (2001) found that POM-C increased by up to 50–55% in the second year of compost application, depending on the time of sampling. An evaluation of fallow frequency, rotation length, N fertilizer, and manure application in wheat rotations, conducted by Bremer et al (1994) also found that light fraction (LF) organic matter was a more sensitive indicator than total soil C, and was affected by all the management options examined. The use of manure increased total soil C from 57.8 to 62.7 Mg ha-1 (increase of 8.5%), while LF-C increased from 12.2% to 14.6% of total C (an increase of 19.7%).
Across these experiments, the parameters most sensitive to changes in management are POM-N concentration and content. Whereas the legume cover crop did not influence either gross pool size (of C or N) or POM-C, it often increased the proportion of total N residing in POM-N fractions (Table 3). Coarse fraction and total POM-N pools were generally enriched by 13–20%, compared to rotations that did not contain legume cover crops. The legume GM treatment (in Potato GM) did not have this effect. The difference between these two experiments, aside from the large number of field replicates and associated statistical power in Clover, may be due to the timing of primary tillage and soil sampling. Clover was tilled in the spring, with samples obtained immediately prior to tillage; Potato GM was tilled in the fall, with samples obtained the following spring.
Both intermittent and annual amendment increased POM-N pools dramatically (Table 3). Even though the amendment in potato mixed was primarily a C source, the N concentration in fine, coarse, and total POM was increased by 15–30% by this treatment, relative to the unamended rotation or 5 years of continuous potato. The amendments used in Eco contained substantial amounts of both C and organic N, and the changes in POM-N pools are of greater magnitude than in Potato Mixed; the concentration of N in POM was approximately doubled by amendment. Likewise, the proportion of soil N residing in POM was increased by up to 40% through annual amendment.
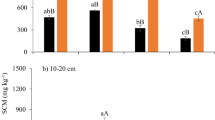
Soil microbial biomass
Legume inclusion in the cropping sequence influenced SMB in only one instance; SMB-C was increased by the red clover cover crop in Clover, from 258 to 296 mg C kg-1 soil (Table 4). Rather than being a long-term, cumulative effect of cover crop, this may reflect transient differences between treatments at the time of sampling. In mid-May, the wheat-sweet corn treatment would contain only decomposing lignified residue from the previous years’ wheat crop, while the wheat(clover)-sweet corn treatment would contain similar residue and actively growing red clover, with the clover providing fresh substrate via root exudates and root turnover. The lack of a significant effect on SMB parameters in samples taken after the sweet corn phase of the same rotation supports this interpretation.
Five years of continuous potato production severely reduced SMB-C and SMB-N, compared to all other treatments in Potato Mixed. The specific activity of the microbial community was also much higher in the continuous potato system, increasing by about 75% compared to all other treatments. The detrimental effect of potato is also shown after the potato phase in the unamended oat-potato rotation, where SMB-N was significantly lower that after the oat phase. This effect was ameliorated by the one-time application of the amendment, where the amended treatment had higher SMB-C and SMB-N, when compared to the non-amended potato phase of the rotation (but not after the non-amended oat phase).
The principle effect in Eco was, again, due to the application of annual amendments with no significant pest management effect. Both SMB-C and SMB-N were, in some cases, more than doubled by the amendment. Although Lalande et al. (1998) found only small, relatively transient changes in SMB-C associated with the application of compost, most others report substantial changes from this amendment. For example, Rochette and Gregorich (1998) found that the application of composted (“rotted”) manure more than doubled SMB. Willson et al. (2001) also found reported increases of >250%, but primarily early in the season, immediately after application.
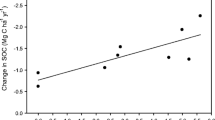
Impact of C input on soil C pools
The difference in C inputs in these experiments ranged from –900 kg ha-1 year-1 (continuous potato versus oat-potato rotation in Potato Mixed) to 8,100 kg ha-1 year-1 (for amended versus unamended plots in Eco). Figure 1 shows that changes in total, POM, and SMB-C pools are strongly influenced by the amount of C being supplied. These data also show that this relationship holds true despite obvious differences in the composition of the inputs across this set of experiments. For example, the C:N ratio ranged from <15 (for red clover biomass) to >100 (for primary papermill sludge).

Relationship between changes in C input and changes in total soil C ( top panel), soil particulate organic matter C ( POM-C; middle panel) and soil microbial biomass C ( SMB-C; bottom panel)
There have been a number of reports of the general relationship between C supply and soil C pools, obtained from medium- to long-term field experiments. For example, differences in straw production in wheat-fallow and wheat rotations was related to changes in soil C content (Campbell et al 1999), although the nature of the relationship was not specified. Similar results were reported by Fauci and Dick (1994) and Drinkwater et al (1998) for different rotations and amendments, and Angers (1992) for alfalfa versus corn monoculture, although these results are confounded to some degree by differences in rotation length and/or tillage frequency. Total and labile C pools in the soil reflect both the quantity and composition of inputs, and the mineralization of fresh and resident C as stimulated by tillage operations; clearly, the quantity of C plays a central role in the intensively tilled systems examined here.
In conclusion, a range of production strategies were evaluated to either minimize or reverse the rapid loss of C and N in intensively tilled short rotations. We found that the insertion of a cover crop or the substitution of GM for grain in 2-year rotations had a minimal effect on soil C pools and total N, but could increase the more labile POM-N fraction. Five years of continuous potato production severely reduced the SMB-C and N concentration, relative to a 2-year, oat-potato rotation system. The application of either a single, large amendment or the annual application of compost and manure led to large increases in total, particulate, and microbial biomass C and N. The quantity of C returned to the soil by these practices strongly controls changes in soil C pools.
References
Alford AR, Drummond FA, Gallandt ER, Groden E, Lambert DA, Liebman M, Marra MC, McBurnie JC, Porter GA, Salas B (1996) The ecology, economics, and management of potato cropping systems: a report of the first four years of the Maine potato ecosystem project. Maine Agricultural and Forest Experiment Station bulletin 843. University of Maine, Orono, Me.
Angers DA (1992) Changes in soil aggregation and organic carbon under corn and alfalfa. Soil Sci Soc Am J 56:1244–1249
Angers DA, Edwards LM, Sanderson JB, Bissonnette N (1999) Soil organic matter quality and aggregate stability under eight potato cropping sequences in a fine sandy loam of Prince Edward Island. Can J Soil Sci 79:411–417
Aoyama M, Angers DA, N’Dayegamiye A, Bissonnette N (1999) Protected organic matter in water-stable aggregates as affected by mineral fertilizer and manure applications. Can J Soil Sci 79:419–425
Beare MH, Cabrera ML, Hendrix PF, Coleman DC (1994a) Aggregate-protected and unprotected organic matter pools in conventional- and no-tillage soils. Soil Sci Soc Am J 58:787–795
Beare MH, Hendrix PF, Coleman DC (1994b) Water-stable aggregates and organic matter fractions in conventional- and no-tillage soils. Soil Sci Soc Am J 58:777–786
Bremer E, Janzen HH, Johnston AM (1994) Sensitivity of total, light fraction and mineralizable organic matter to management practices in a Lethbridge soil. Can J Soil Sci 74:131–138
Cambardella CA, Elliott ET(1992) Particulate soil organic matter changes across a grassland cultivation sequence. Soil Sci Soc Am J 56:777–783
Campbell CA, Biederbeck VO, McConkey BG, Curtin D, Zentner RP (1999) Soil quality—effect of tillage and fallow frequency. Soil organic matter quality as influenced by tillage and fallow frequency in a silt loam in southwestern Saskatchewan. Soil Biol Biochem 31:1–7
Chantigny MH, Angers DA, Beauchamp CJ (1999) Aggregation and organic matter decomposition in soils amended with de-inking paper sludge. Soil Sci Soc Am J 63:1214–1221
Carter MR, Sanderson JB (2001) Influence of conservation tillage and rotation length on potato productivity, tuber disease and soil quality parameters on a fine sandy loam in eastern Canada. Soil Tillage Res 63:1–13
Collins HP, Rasmussen PE, Douglas CL Jr (1992) Crop rotation and residue management effects on soil carbon and microbial dynamics. Soil Sci Soc Am J 56:783–788
Davis AS (1998) Effects of organic nitrogen sources and timing of nitrogen fertilizer application on crop-weed interference. M.S. thesis. University of Maine, Orono, Me.
Drinkwater LE, Wagoner P, Sarrantonio M (1998) Legume-based cropping systems have reduced carbon and nitrogen losses. Nature 396:262–265
Fauci MF, Dick RP (1994) Soil microbial dynamics: short- and long-term effects of inorganic and organic nitrogen. Soil Sci Soc Am J 58:801–806
Franzluebbers AJ, Arshad MA (1996) Soil organic matter pools during early adoption of conservation tillage in northwestern Canada. Soil Sci Soc Am J 60:1422–1427
Franzluebbers AJ, Haney RL, Hons FM, Zuberer DA (1999a) Assessing biological soil quality with chloroform fumigation-incubation: why subtract a control. Can J Soil Sci 79:521–528
Franzluebbers AJ, Langdale GW, Schomberg HH (1999b) Soil carbon, nitrogen, and aggregation in response to type and frequency of tillage. Soil Sci Soc Am J 63:349–355
Gagnon B, Lalande R, Fahmy SH (2001) Organic matter and aggregation in a degraded potato soil as affected by raw and composted pulp residue. Biol Fertil Soils 34:441–447
Gallandt ER, Mallory EB, Alford AR, Drummond FA, Groden E, Liebman M, Marra MC, McBurnie JC, Porter GA (1998) Comparison of alternative pest and soil management strategies for Maine potato production systems. Am J Alternat Agric 13:146–161
Griffin TS, Hesterman OB (1991) Potato response to legume and fertilizer nitrogen sources. Agron J 83:1004–1012
Haynes RJ, Tregurtha R (1999) Effects of increasing periods under intensive arable vegetable production on biological, chemical, and physical indices of soil quality. Biol Fertil Soils 28:259–266
Hu S, Grunwald NJ, van Bruggen AHC, Gamble GR, Drinkwater LE, Shennan C, Demment MW (1997) Short-term effects of cover crop incorporation on soil carbon pools and nitrogen availability. Soil Sci Soc Am J 61:901–911
Hussain I, Olson KR, Ebelhar SA (1999) Long-term tillage effects on soil chemical properties and organic matter fractions. Soil Sci Soc Am J 63:1335–1341
Islam KR, Weil RR (1998) Microwave irradiation of soil for routine measurement of microbial biomass. Biol Fertil Soils 27:408–416
Jackson LE (2000) Fates and losses of nitrogen from a nitrogen-15-labeled legume cover crop in an intensively managed vegetable system. Soil Sci Soc Am J 64:1404–1412
Jenkinson DS, Powlson DS (1976) The effects of biocidal treatment on metabolism in soil. V. A method for measuring soil biomass. Soil Biol Biochem 8:209–213
Lalande R, Gagnon B, Simard RR (1998) Microbial biomass C and alkaline phosphatase activity in two compost amended soils. Can J Soil Sci 78:581–587
Liebman M, Gallandt E (2002) Differential responses to red clover ( Trifolium pratense) residue and ammonium nitrate fertilizer by common bean ( Phaeseolus vulgaris) and wild mustard ( Brassica kaber). Weed Sci 50:521–529
McVay KA, Radcliffe DE, Hargrove WL (1989) Winter legume effects on soil properties and nitrogen fertilizer recommendations. Soil Sci Soc Am J 53:1856–1862
Mendes IC, Bandick AK, Dick RP, Bottomley PJ (1999) Microbial biomass and activities in soil aggregates affected by winter cover crops. Soil Sci Soc Am J 63:873–881
Moore JM, Klose S, Tabatabai MA (2000) Soil microbial biomass carbon and nitrogen as affected by cropping system. Biol Fertil Soils 31:200–210
Plotkin JMB (2000) The effects of green manure rotation crops on soils and potato yield and quality. M.S. thesis. University of Maine, Orono, Me.
Porter GA, Opena GB, Bradbury WB, McBurnie JC, Sisson JA (1999) Soil management and supplemental irrigation effects on potato. I. Soil properties, tuber yield, and quality. Agron J 91:416–425
Quemada M, Cabrera ML (1995) Carbon and nitrogen mineralized from leaves and stems of four cover crops. Soil Sci Soc Am J 59:471–477
Rochette P, Gregorich EG (1998) Dynamics of soil microbial biomass C, soluble organic C and CO2 evolution after three years of manure application. Can J Soil Sci 78:283–290
Rochette P, Angers DA, Cote D (2000) Soil carbon and nitrogen dynamics following application of pig slurry of the 19th consecutive year: carbon dioxide fluxes and microbial biomass carbon. Soil Sci Soc Am J 64:1389–1395
Voroney RP, Paul EA (1984) Determination of k C and k N in situ for calibration of the chloroform fumigation-incubation method. Soil Biol Biochem 16:9–14
Willson TC, Paul EA, Harwood RR (2001) Biologically active soil organic matter fractions in sustainable cropping systems. Appl Soil Ecol 16:63–76
Acknowledgements
The assistance of E. Mallory in conducting laboratory assessments is gratefully acknowledged. The assistance of Matt Liebman and Adam Davis in providing information on the CLOVER experiment is also appreciated. Mention of trade names or commercial products in this article is solely for the purpose of providing specific information, and does not imply recommendation or endorsement by the U.S. Department of Agriculture.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Griffin, T.S., Porter, G.A. Altering soil carbon and nitrogen stocks in intensively tilled two-year rotations. Biol Fertil Soils 39, 366–374 (2004). https://doi.org/10.1007/s00374-004-0725-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00374-004-0725-7