
Abstract
Physiological responses to dehydration in amphibians are reasonably well documented, although little work has addressed this problem in hibernating animals. We investigated osmotic and metabolic responses to experimental manipulation of hydration state in the wood frog (Rana sylvatica), a terrestrial hibernator that encounters low environmental water potential during autumn and winter. In winter-conditioned frogs, plasma osmolality varied inversely with body water content (range 69–79%, fresh mass) primarily due to increases in sodium and chloride concentrations, as well as accumulation of glucose and urea. Decreased hydration was accompanied by a marked reduction in the resting rate of oxygen consumption, which was inversely correlated with plasma osmolality and urea concentration. In a separate experiment, resting rates of oxygen consumption in fully hydrated frogs receiving injections of saline or saline containing urea did not differ initially; however, upon dehydration, metabolic rates decreased sooner in the urea-loaded frogs than in control frogs. Our findings suggest an important role for urea, acting in concert with dehydration, in the metabolic regulation and energy conservation of hibernating R. sylvatica.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
One general characteristic of the Amphibia is a moist skin that provides little resistance to water flux. Yet, several species thrive in environments where desiccating conditions occur frequently and may persist for long periods. Decades of research on this apparent paradox has revealed a host of mechanisms for coping with water deficit among amphibians (Jørgensen 1997a). For example, water conservation is aided by synthesis and/or accumulation of organic and inorganic osmolytes that raise internal osmotic pressure, thereby reducing the transcutaneous gradient for water loss (Katz and Hoffman 1990; Shoemaker 1964). Increases in extracellular concentrations of the major plasma electrolytes sodium and chloride make up a significant portion of the osmotic increase (Shoemaker 1964). However, various organic solutes can accumulate in lieu of electrolytes in order to avert deleterious effects on macromolecules (Hochachka and Somero 2002). Among amphibians, the end-product of nitrogen metabolism, urea, is the predominant organic osmolyte accumulated during dehydration (Jørgensen 1997b), although some evidence suggests that glucose concentration may also increase (Churchill and Storey 1993; Edwards et al. 2004).
Several species of temperate anurans overwinter beneath a layer of forest detritus. They are susceptible to dehydration in this microenvironment because soil moisture reaches an annual minimum in autumn and freezing of the soil during winter further reduces its water potential (Spaans and Baker 1996). Myriad studies have addressed the responses to dehydration of active or estivating anurans, but those of hibernating frogs have largely been ignored. Such responses may be vastly different in overwintering frogs because various correlates of hibernation, including aphagia, hypothermia, altered hormonal state and patterns of gene expression, and decreased renal function, can strongly influence physiological state (Pinder et al. 1992).
Terrestrially-hibernating frogs apparently can become dehydrated and accumulate urea in late fall and winter. Body water contents of wood frogs (Rana sylvatica) kept in an outdoor enclosure were substantially lower and plasma urea concentrations ∼25-fold higher in autumn and winter than in spring, when rain and snow melt increased water availability (Costanzo and Lee 2005). In this same study, winter-conditioned frogs exposed to low water potential in the laboratory accumulated ∼90 mmol l−1 urea in plasma. Layne and Rice (2003) found >30 mmol l−1 urea in plasma of winter R. sylvatica, and the grey treefrog (Hyla versicolor) apparently can accumulate >50 mmol l−1 urea (J. R. Layne, unpublished data). However, it remains unclear how other organic osmolytes and ions respond during dehydration in terrestrially hibernating frogs. Costanzo and Lee (2005) reported that glucose increased during hibernation, but the portion of increase due to dehydration rather than freezing, which is a potent stimulator of glycogenolysis, was not determined.
It is likely that various osmolytes accumulated during water stress have ancillary physiological effects, some of which could be beneficial. In R. sylvatica, for example, glucose provides tissues with a ready energy source, and both glucose and urea are effective cryoprotective agents (Costanzo and Lee 2005). Preliminary findings indicate that elevated urea also depresses metabolism in certain tissues, potentially conserving limited energy reserves (Costanzo and Lee 2005). Although the rationale for metabolic regulation by urea has been proposed (Hand and Somero 1982; Yancey et al. 1982) definitive empirical evidence for such inhibition is lacking.
Our goal in this project was to examine physiological responses to dehydration of the terrestrial hibernator, R. sylvatica. We measured plasma concentrations of organic and inorganic osmolytes, and resting rates of oxygen consumption, in winter-conditioned frogs following experimental manipulation of body water content. To test the putative role of urea as a metabolic inhibitor, we compared rates of oxygen consumption in frogs administered physiological saline or saline containing urea. Finally, monitoring rates of oxygen consumption of these urea-loaded frogs during progressive dehydration allowed us to examine the metabolic effect of hyperuremia over a range of hydration states.
Materials and methods
Animals and acclimation
Male R. sylvatica were collected in southern Ohio at a breeding pond in February 2004, 2005, and 2006. Frogs were brought to Miami University, transferred to boxes containing damp moss, and held at 4°C. Some frogs were used in laboratory experiments within 6 weeks of capture. Others were transferred to a 48-m2 outdoor enclosure in a wooded area of the Miami University Ecological Research Center and used the following winter. We fed these frogs crickets thrice weekly and made water continually available. In late October, feeding was suspended and the frogs, on the verge of entering dormancy, were recaptured by hand, brought to the laboratory and held on damp moss in darkened boxes at 4°C for at least 2 weeks before being used in experiments. This husbandry protocol keeps frogs fully hydrated (body water content ∼78–80%, fresh mass; Costanzo et al. 1993; Layne and Lee 1987) with low concentrations of plasma urea (∼4 mmol l−1; Costanzo and Lee 2005) and presumably facilitates entry into dormancy.
All experiments were performed at 4°C. Frogs were prepared for use in the experiments by removing bladder fluid via insertion of a polished glass cannula into the cloaca. We then measured standard body mass to the nearest 0.1 mg on an electronic balance.
Effect of hydration state on plasma osmolytes and metabolism
Frogs collected from the outdoor enclosure in autumn and exposed to simulated winter conditions in the laboratory were transferred from their holding boxes to individual plastic containers fitted with perforated lids. Our aim in this experiment was to cause dormant frogs to dehydrate to various degrees by exposing them to different hydric conditions. Frogs destined to dehydrate substantially (n = 8) were placed on a substratum of dry paper towel, whereas other frogs (n = 11) were placed on a pad of variably wetted moss. The latter group included frogs provided with sufficient moisture that they could remain fully hydrated throughout the experiment. All frogs were held in darkness for 11–13 days, after which each was used in a respirometry trial.
Respirometry trials were run on five occasions during December, January and February. Each trial used 3–5 frogs, including at least one frog from a “dry” cup and one from a “moist” cup. Immediately after completion of the trial, each frog was removed from its respirometry chamber, double-pithed, and dissected. Blood was collected from the aortic arch into heparinized microhematocrit tubes and centrifuged at 4,000g for 5 min, and the resulting plasma was collected and stored at −80°C for future analysis of osmolytes. The carcass was weighed to the nearest 0.1 mg, thoroughly dried at 65°C in an oven, and weighed again. Body water content, determined from the change in mass during drying, was expressed as a percentage of the carcass fresh mass.
Effect of urea loading and dehydration on metabolism
In this experiment, we manipulated blood urea concentration in order to test for an inhibitory effect of urea on metabolism. Frogs, taken from their holding boxes within 6 weeks of capture at the breeding pond, were placed in perforated plastic cups on dry paper towel and held in darkness at 4°C for 2–3 days. After body mass was reduced by ∼0.5 g (presumably via evaporation of body water), each frog in a control group (n = 5) was given an injection of isotonic phosphate-buffered saline (PBS; in g l−1; 6.10 NaCl, 0.15 KCl, 0.88 Na2HPO4, 0.15 KH2PO4; 230 mosmol kg−1), whereas each frog in a treatment group (n = 5) received PBS containing 1.5 M urea. The injectant (volume, ∼3% of standard body mass) was delivered into the dorsal lymph pad using a 27.5-gauge needle. Next, we held each frog inside a metabolic chamber for 6 days before measuring its rate of oxygen consumption. We assumed this incubation period allowed ample time for manifestation of any inhibitory effect of urea on metabolism. After the respirometry trial, each frog received an additional injection of PBS with or without urea, as appropriate, and was returned to its metabolic chamber. Oxygen consumption rate was measured 3 days later. Mean rates of oxygen consumption were compared between urea-loaded and control groups at both sample times using separate Student’s t tests.
In order to test for an inhibitory effect of urea on metabolism in conjunction with dehydration, the experiment described above was extended by periodically measuring oxygen consumption rates whilst urea-loaded and control frogs were progressively dehydrated inside their metabolic chambers. This was accomplished by passing a slow stream of air (4°C, relative humidity ∼60%) through the metabolic chambers for 14 h day−1, every other day. Frogs remained undisturbed inside the darkened chambers throughout the dehydration regimen. We measured the rate of oxygen consumption of each frog on days 11, 15, and 23 after the start of dehydration; these sample times corresponded to 16, 20, and 28 days after frogs received the second set of injections. Mean rates of oxygen consumption were compared between and within control and urea-loaded groups over the course of the dehydration regimen using a two-factor repeated-measures analysis of variance (ANOVA), followed by least-squares means comparisons.
After the final respirometry measurement, each frog was removed from its metabolic chamber and placed on its back to determine whether or not it was capable of righting itself. It was then weighed to the nearest 0.1 mg, double-pithed and dissected, and blood plasma was collected and stored for later analysis. Carcass water content was determined gravimetrically as previously described. We estimated the body water content of each frog prior to entering the dehydration regimen by adding the mass of water remaining in the carcass at the end of the experiment to the arithmetic difference between the initial and final standard mass values and dividing that sum by the initial standard mass. In making this estimate, we assumed the observed decrease in body mass during the experiment was due solely to water loss. Mean values for plasma osmolality, concentrations of organic osmolytes, and body water content were compared between urea-loaded and control groups using Student’s t tests. Analyses involving percentage data were performed after values had been arcsine/square-root transformed.
Respirometry
Resting rate of oxygen consumption was measured using closed-system respirometry. Each frog was habituated at 4°C in darkness for 24–30 h inside an acrylic metabolic chamber (Qubit Systems no. G115) with valved incurrent and excurrent ports. During this time, the port valves were kept open and the partial pressure of oxygen inside the chambers approximated the atmospheric level (∼21%). Rate of oxygen consumption was calculated from an overnight (1800–0900 hours) trial following the methods of Vleck (1987). Briefly, each chamber was ventilated with room air and an initial gas sample was drawn into a 60-ml Hamilton syringe fitted with a Teflon stopcock. The incurrent and excurrent valves were closed in close sequence in order to equilibrate chamber and ambient pressures. Duration of the trial was timed from that point. We did not monitor activities of the frogs during the trials, but assumed them to be quiescent due to the low temperature, season of testing, and lack of light, noise, and other stimuli. The final gas sample was taken the following morning.
Each gas sample was thermally equilibrated with room air and analyzed for partial pressure of oxygen using a single-cell Ametek S-3A O2 analyzer. Rate of oxygen consumption was calculated for each frog from the difference in readings from the two gas samples using equations from Vleck (1987). Values were expressed as microliters of O2 consumed per gram dry mass of frog per hour at STDP.
Plasma osmolyte analysis
Frozen samples of blood plasma were thawed on ice and assayed for osmolality and select organic and inorganic osmolytes. Osmolality was measured in 10-μl plasma samples using a Wescor vapor-pressure osmometer (model 5500; Logan, UT, USA) calibrated with NaCl standards. We used colorimetric assays to measure concentrations of the organic osmolytes urea (urease, no. B7551–120, Pointe Scientific, Canton, MI, USA), glucose (glucose oxidase, no. 510; Sigma, St. Louis, MO, USA), and lactate (lactate dehydrogenase, no. 735, Sigma). Samples of plasma from frogs in the first experiment were also analyzed using ion-selective electrodes for sodium (MI-420 Microelectrodes, Inc., Bedford, NH, USA) and chloride (ISM-146, Lazar Research Laboratories, Inc., Los Angeles, CA, USA) and a micro-reference electrode (MI-409, Microelectrodes, Inc.). Serial dilutions of 0.1 M NaCl standards were prepared in sodium- or chloride-free PBS and used to generate standard curves for these electrodes. An assayed standard derived from human serum (Ser-T-Fy, no. G427, Stanbio Laboratory, Boerne, TX, USA) was then used for point calibration of each electrode and the standard curves were adjusted as necessary.
Statistical analysis
Statistical analyses were performed using SAS 9.1 and StatView statistical programs and significance was accepted at P ≤ 0.050. Mean values are reported ±SEM.
Results
Effect of hydration state on plasma osmolytes and metabolism
All animals used in this experiment appeared in good health and were capable of coordinated movement at the conclusion of the trial. Changes in standard mass of individual frogs during the trial varied from +0.51 g to −4.51 g and presumably reflected only changes in hydration state. Body water content of these frogs ranged 69.0–79.1%, fresh mass. We regarded those frogs having a body water content near 79% fresh mass to be fully hydrated (Costanzo et al. 1993; Layne and Lee 1987) and representative of control animals.
Plasma osmolality ranged from 224–446 mosmol kg−1 (n = 19) and was inversely correlated (R 2 = 0.727; P < 0.001) with body water content (Fig. 1a). Plasma concentrations of urea (range 2.4–49.9 mmol l−1; n = 19) and glucose (range 0.8–16.7 mmol l−1; n = 19) were also inversely correlated (urea: R 2 = 0.693; P < 0.001, glucose: R 2 = 0.654; P < 0.001) with hydration level. However, lactate showed substantially less variation (range 3.3–10.1 mmol l−1; n = 19) and no association (R 2 = 0.010; P = 0.683) with body water content (Fig. 1b). We found considerable variation in plasma concentrations of both sodium (range 102–196 mEq l−1; n = 16) and chloride (range 64–102 mEq l−1; n = 16). Of the two ions, sodium had the greater variability among individuals with similar body water contents (Fig. 1c). Nevertheless, strong inverse correlations with body water content were found for sodium (R 2 = 0.382; P = 0.011) and chloride (R 2 = 0.836; P < 0.001).

Experimental manipulation of hydration in winter-conditioned R. sylvatica and its effect on a plasma osmolality; b plasma concentrations of principal organic osmolytes; and c plasma concentrations of major inorganic ions
Based on the linear regression equation, the difference in plasma osmolality between a frog with a body water content value of 69% and one with a value of 79% was estimated to be 154 mosmol kg−1 (Table 1). Corresponding differences in measured plasma osmolytes were 34 mmol l−1 for urea, 12 mmol l−1 for glucose, 69 mEq l−1 for sodium, and 35 mEq l−1 for chloride. Summing these values gave a total of 150 mosmol kg−1, which closely matched the osmotic differential. We calculated that the loss of water (∼4.8 g in the average-size frog) associated with a change in body water content from 79 to 69% would cause the concentration of extant solutes to increase 1.69-fold (Shoemaker 1964). Whereas corresponding differences in sodium and chloride concentrations were on that order (∼1.5-fold), differences in urea and glucose concentrations were substantially greater (∼5-fold and ∼13-fold, respectively), indicating that additional solute had been added to the blood as the frogs dehydrated.
Because a high degree of multicollinearity in predictor variables rendered multiple regression an unsuitable tool, we used separate regression analyses to evaluate relationships between rate of oxygen consumption \( (\ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} ) \) and individual plasma osmolytes, plasma osmolality, and body water content. Generally, lower \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \) was found in frogs having lower body water contents (R 2 = 0.225; P = 0.040; n = 19; Fig. 2). \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \) ranged from 28.0–51.4 μl g−1 h−1 and the linear regression model suggested that frogs having the lowest body water content (69% of fresh mass) had \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \) values that were ∼20% below those of fully hydrated (79% of fresh mass) frogs.

Experimental manipulation of hydration in winter-conditioned R. sylvatica and its effect on rate of oxygen consumption
\( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \) was also inversely correlated with plasma osmolality, which showed a stronger relationship with \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \) than did body water content (Table 2). Regression analyses of individual plasma osmolytes and \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \) revealed a significant association with urea, but not with any other solute. Visual inspection of the binomial scatter-plots suggested that certain variables would better fit a logarithmic model and, indeed, higher coefficients of determination resulted after log-transforming the data for plasma osmolality (R2 = 0.419; P = 0.003) and urea (R2 = 0.428; P = 0.003). Moreover, these two regressions gave similar measures of fit, indicating that plasma urea concentration was as good a predictor of \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \) as total plasma osmolality (Fig. 3).

Variation in rate of oxygen consumption in winter-conditioned R. sylvatica as a function of plasma osmolality or plasma urea concentration (inset)
Effect of urea loading and dehydration on metabolism
Urea loading did not significantly (P = 0.930) alter \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \) relative to that of saline-injected control frogs. Mean values of \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \) were 36.9 ± 2.0 μl g−1 h−1 for urea-loaded frogs (n = 5) and 36.7 ± 0.5 μl g−1 h−1 for control frogs (n = 5). Augmenting blood urea level via the second injection had no effect on \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \). However, \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \) in urea-loaded and control frogs, ∼41 μl g−1 h−1, was slightly higher at this time than after the first set of injections.
We determined the metabolic effect of hyperuremia over a range of hydration states by monitoring \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \) in the urea-loaded and saline-injected control frogs during progressive dehydration. At the end of this trial, all frogs were capable of righting themselves when placed on their backs. Urea-loaded and control frogs lost 29.2 ± 1.2% (n = 5) and 24.6 ± 1.6% (n = 5) of their initial standard body mass, respectively, and mean body water content did not differ between the groups (Table 3). Expectedly, plasma osmolality and urea concentration were markedly higher in urea-loaded frogs, whereas plasma glucose and lactate levels in these frogs were similar to those of controls (Table 3). Unexpectedly, the differential in plasma osmolality between urea-loaded and control frogs, ∼225 mosmol kg−1, was substantially greater than can be explained by differences in measured urea concentrations (∼125 mmol l−1). Plasma glucose and lactate levels in both control and urea-loaded frogs, and osmolality and urea levels in control frogs, were comparable to those in frogs with similar body water contents from the initial experiment (Fig. 1).
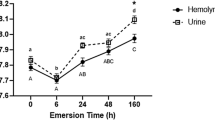
\( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \) in urea-loaded and control frogs fell as dehydration progressed, reaching a level ∼25% below initial values by the end of the trial (two-factor repeated-measures ANOVA, P < 0.001). The decrease in \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \) was first detected on day 11 in urea-loaded frogs, but not until day 23 in control frogs (Fig. 4). With the exception of the final measurement, urea-loaded frogs had nominally (but not significantly, P = 0.309) lower mean \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \) values than control frogs at each sampling. However, residual analysis from the ANOVA indicated that outlying \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \) values of two control frogs in the initial sample potentially confounded the result. Omitting those values from the analysis rendered the interaction between time and group significant (P = 0.042), indicating that urea-loading did, in fact, influence the response to dehydration. Accordingly, mean \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \) of urea-loaded and control frogs differed significantly on day 11 (Fig. 4). At the end of the trial, body water content of frogs in both groups was ∼70% fresh mass (Table 3). Mean \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \) was ∼30 μl g−1 h−1, comparable to that measured in similarly hydrated frogs in the first experiment.

Effect of urea-loading followed by progressive dehydration on rate of oxygen consumption in winter-conditioned R. sylvatica. Asterisks denote means (±SEM; n = 5) that differ significantly from the corresponding initial value. Dagger denotes that the mean for the urea-loaded group differed significantly from that of the control group at the same sample time. Initial measurements were made 2 days prior to beginning the dehydration regimen (day 0). Symbols at day -2 and day 23 are offset for clarity
Discussion
Behavioral and physiological responses of amphibians to a dehydrating environment are generally well understood for “active” animals. Various combinations of water-conserving behaviors, such as posturing, aggregation, and burrowing, are used in concert with a suite of physiological adaptations that minimize water loss to the environment. Increasing the osmolality of body fluids is a universal strategy in amphibian water conservation and is achieved by passively concentrating extant solutes via solvent loss as well as synthesizing new osmolytes. The classes of osmolytes accumulated, and the extent to which they are accrued, reflect the necessity for maintaining an intracellular milieu that is compatible with protein function (Hochachka and Somero 2002). Solutes that are benign to macromolecules are referred to as “compatible osmolytes” and are usually accumulated in favor of more perturbing osmolytes, such as inorganic ions (Yancey et al. 1982). Besides their colligative effects, many accumulated solutes have other physiological roles. For instance, glucose and urea are both important cryoprotectants in hibernating R. sylvatica (Costanzo and Lee 2005).
Rana sylvatica, one of the most dehydration-tolerant of all North American ranids, (Schmid 1965), probably remains in negative water balance during winter. It seems to prefer well-drained upland habitats for overwintering (Baldwin et al. 2006; Regosin et al. 2003) and any moisture in its hibernaculum becomes unavailable once frost permeates the duff and soil. Depending on prevailing weather conditions, R. sylvatica can remain partly dehydrated and hyperuremic throughout winter, until spring rains increase moisture availability (Costanzo and Lee 2005).
In the present study, the general osmotic response to dehydration in winter-conditioned R. sylvatica was similar to that previously shown for “active” or estivating anurans. Plasma osmolality increased via solute concentration by a net loss of body water, and also through accumulation of certain solutes. Levels of sodium and chloride ions increased sharply as body water was lost, but apparently the rise reflected only the concentrating effect of the reduced solvent volume, rather than an increase in number of ions (Shoemaker 1964). Although these increases were substantial, we doubt that our dehydrated frogs suffered any harm. Indeed, many amphibians tolerate an elevation in plasma sodium concentration by ∼100 mEq l−1 (Shoemaker et al. 1992) and can avoid osmotic damage at nonpermeant osmolyte concentrations below ∼440 mosmol l−1 (Hillman 1978a).
Accumulation of organic solutes is a common strategy used by organisms to increase internal osmotic pressure without risking ionic perturbation of macromolecules (Hochachka and Somero 2002). Mobilization of urea, and to a lesser extent, glucose, accounted for a substantial increase in plasma osmolality during dehydration of winter-conditioned R. sylvatica. Urea accumulation in these frogs was consistent with previous reports for non-hibernating amphibians (Balinsky 1981; Katz and Hoffman 1990) and also compared favorably with that of R. sylvatica maintained in an outdoor enclosure during autumn and winter (Costanzo and Lee 2005).
Glucose mobilization is not widely recognized as a part of the amphibian response to dehydration, yet, in the present study, plasma glucose levels increased markedly as frogs lost water to the environment. Earlier reports attest that dehydration leads to hyperglycemia in R. sylvatica (Churchill and Storey 1993) and the terrestrial hibernator, Pseudacris triseriata (Edwards et al. 2004), suggesting that glucose contributes to water conservation in some species. Nevertheless, in our dehydrated frogs glucose accounted for only ∼3% of the total osmotic pressure.
In our first experiment, the difference in plasma osmolality between fully-hydrated and dehydrated frogs, ∼150 mosmol kg−1, was mostly accounted for by changes in concentrations of the solutes we measured. Frogs may have accumulated other osmolytes, such as inorganic ions, amino acids, and other metabolites, but the combined osmotic contribution of these solutes must have been minimal.
Previous investigations of metabolic responses to dehydration in amphibians have documented decreases in measures of activity metabolism, such as aerobic scope, maximum metabolic rate, and locomotor endurance, but found minimal change in resting metabolic rate (Gatten 1987; Gil and Katz 1996; Hillman 1978a, 1987; Preest et al. 1992). In the present study, dehydration of winter-conditioned R. sylvatica resulted in a sharp decrease in resting metabolic rate (Fig. 2). Our finding that plasma lactate concentration was independent of hydration level implies that metabolism in dehydrated frogs remained primarily aerobic; thus, the observed fall in \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \) apparently reflected a reduction in total metabolic rate. Other workers have observed hypometabolism in estivating or diapausing ectotherms, but did not elucidate the relationship between metabolism and dehydration, which often initiates or accompanies these states (Flanigan et al., 1991; Kennett and Christian 1994; Secor 2005; Williams and Lee 2005). Apparently, our findings for R. sylvatica constitute the first evidence for a direct link between hydration state and resting metabolism in any dormant amphibian.
Our experimental results offer several clues about the mechanism underlying the observed hypometabolism in dehydrated frogs. First, the strong relationship between \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \) and plasma osmolality suggests that hyperosmotic activity of body fluids is involved. Reduced water concentration and associated solute crowding potentially imposes osmophobic and hydrophobic effects on macromolecules. Because both osmophobicity and hydrophobicity are important determinants of proper protein folding as well as construction of multi-protein complexes (Hochachka and Somero 2002), modification of protein folding or protein-protein interactions could lead to an altered metabolic state. On the other hand, we noted that plasma concentrations of sodium and chloride, which rose dramatically in dehydrating frogs, were not correlated with; \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2}; \) thus, it seems unlikely that the response could be attributable to purely osmotic effects.
Alternatively, hypometabolism in dehydrated R. sylvatica could result from an accumulation of a particular solute(s). We found that \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \) was strongly correlated with urea, but not with any of the other measured osmolytes, and that urea was as good a predictor of \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \) as plasma osmolality (Table 2). Possibly, then, urea played a direct role in depressing metabolism of these frogs. In principle, this effect could stem from its influence on the structure and function of key regulatory enzymes. When present in a specific ratio with urea, counteracting solutes (such as TMAO and other methylamines) mitigate the perturbing effects of urea, permitting enzymes to function properly in tissues that have constitutively high levels of urea (Yancey and Burg 1990; Yancey et al. 1982). However, in the absence of such compounds, urea could alter enzyme activity. Co-accumulation of counteracting solutes with urea occurs in elasmobranches and the mammalian kidney, but in the few species of amphibians that have been examined concentrations of these compounds are minimal (i.e., ≤10 mmol l−1; Withers and Guppy 1996; Wray and Wilkie 1995).
The notion that, in the absence of counteracting solutes, elevated urea might contribute to metabolic regulation in dormancy by reversibly inhibiting key metabolic enzymes was proposed long ago (Hand and Somero 1982; Yancey et al. 1982) and remains prevalent in the contemporary literature (Arad 2001; Shpun et al. 1992; Somero 1986; Withers and Guppy 1996; Wright 1995). Nevertheless, this hypothesis garners empirical support only from a handful of in vitro studies. Urea in physiological concentrations, in some cases as low as 45 mmol l−1, inhibits activity of certain metabolic enzymes, such as phosphofructokinase (Cowan and Storey 2002; Hand and Somero 1982), lactate dehydrogenase (Fuery et al. 1997), and Na–K ATPase (Kaji et al. 1998). Urea also slows growth of cells in culture (Yancey and Burg 1990) and apparently depresses \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \) in certain tissues of winter R. sylvatica (Costanzo and Lee 2005). Our present findings are unique in making a strong direct link between elevated urea and hypometabolism at the organismal level.
Although the urea-hypometabolism hypothesis is an attractive explanation for our finding, results of our urea-loading experiment suggest complexity in the mechanism by which urea induces hypometabolism. For example, whereas augmenting blood urea levels in fully-hydrated frogs had no discernable effect on \( \ifmmode\expandafter\dot\else\expandafter\.\fi{V}{\text{O}}_{2} \), hyperuremia in combination with dehydration was a potent stimulus, as metabolic depression was sooner manifested in urea-loaded frogs than in control frogs (Fig. 4). Although we cannot explain this outcome at present, one possibility is that dehydration-induced hydrophobic and osmophobic effects on metabolic enzymes rendered them more susceptible to the perturbing effects of urea. Alternatively, perhaps dehydration triggered changes in gene transcription that resulted in expression of urea-sensitive isozymes. Notably, urea sensitivity of various metabolic enzymes is both species- and isoform-specific (Cowan and Storey 2002; Fuery et al. 1997; Grundy and Storey 1994) and estivation in urea-accumulating anurans, which is accompanied by profound hypometabolism, is associated with production of kinetically-distinct forms of certain enzymes (Fuery et al. 1997).
At the end of our dehydration trials, mean plasma urea concentration for urea-loaded frogs was 124 mmol l−1 higher than that for control frogs, whereas the corresponding differential for plasma osmolality was 226 mosmol kg−1 (Table 3). We do not know the reason for the lack of congruence between these values, but we can offer a possible explanation. First, we are confident in the osmolality data because the mean values for both groups were close to the respective values predicted from solvent volume reduction, osmolyte accumulation, and (in the urea-loaded group) urea augmentation. The disparity, therefore, apparently stems from an unexpectedly low uremia in the urea-loaded frogs. Our urea-loading protocol should have increased plasma urea by about 140 mmol l−1 (assuming uniform dispersion of the solute throughout all fluid compartments of the body and a body water content of 79% fresh mass) and, during dehydration, plasma urea would have further risen to 235–268 mmol l−1, the actual concentration depending on whether or not these frogs synthesized urea, as did control frogs (Table 3). However, we measured only 162 mmol l−1 in these frogs. What accounts for the disparity? These frogs did not urinate and urea efflux through amphibian skin is negligible (Garcia-Romeu et al. 1981). Catabolism is improbable because, to our knowledge, the ornithine–urea cycle is irreversible and frogs lack urease and other mechanisms for recycling urea. Possibly, the apparent shortfall reflects a disproportionate distribution of the urea to intracellular spaces. Recent findings indicate that some classes of urea transporters facilitate movement of urea against its concentration gradient leading to sequestration of this solute in certain tissues (McDonald et al. 2006). Whether or not these transporters generally occur in amphibian tissues remains to be determined; however, in frogs subjected to dehydration, water inside muscle cells is somehow defended at the expense of vascular water (Hillman 1978b). The potential role of urea sequestration in cell volume regulation in amphibians merits additional study.
Despite the large difference in plasma osmolality, urea-loaded and control frogs apparently lost similar amounts of water during experimental dehydration. This result seemingly contradicts the notion that urea accumulation is an effective water-conservation response. However, because these frogs were exposed to air, and because unsaturated air has a much more negative water potential than body fluids with solutes at almost any physiological concentration (Spotila et al. 1992), the very large vapor gradient driving water efflux would have minimized the osmotic advantage held by the urea-loaded group.
Rana sylvatica encountering low environmental water potential during autumn and winter tend to dehydrate and accumulate urea, which serves as a cryoprotectant (Costanzo and Lee 2005) and, as our present findings suggest, a metabolic inhibitor. The latter function would conserve the endogenous energy substrates needed for reproductive activities, which occur in late winter, before feeding resumes. The 20–25% reduction in metabolism in our R. sylvatica is substantial, though relatively modest compared to the ∼75% reduction exhibited by some anurans in estivation (Pinder et al. 1992). We suspect that a more substantial inhibition would be manifested in R. sylvatica at even slightly higher body temperatures. Metabolism in our winter-conditioned (4°C) frogs was already quite low and, perhaps, unamenable to much further reduction. Additionally, the stabilizing effect of cold on proteins could have partly countered the destabilizing effect of urea (Hochachka and Somero 2002). Although the thermal dynamics of the hypometabolic response remain to be established, given the high Q10 for metabolism at typical winter temperatures (Pinder et al. 1992), its importance in energy conservation would increase markedly with even an incremental rise in body temperature.
In conclusion, winter-conditioned R. sylvatica respond to experimental dehydration by accumulating various inorganic and organic solutes, including urea, and by reducing metabolic rate. Our experimental results suggested that hypometabolism in these frogs is a response to elevated urea in combination with reduced hydration. Accordingly, we postulate that the pattern of dehydration and urea accumulation upon entry into dormancy, and rehydration and elimination of urea upon arousal (Costanzo and Lee 2005), may control metabolic depression and reactivation in R. sylvatica and other terrestrially hibernating frogs. Furthermore, given that urea accumulation and metabolic depression are coupled physiological responses in dormancy in numerous amphibians (Pinder et al. 1992), lungfishes (Chew et al. 2004; Loong et al. 2005), and molluscs (Arad 2001; Rees and Hand 1993), this system of metabolic regulation may be generally important in diverse taxa of ectothermic animals.
We thank P. Baker and M. Elnitsky for aid in the frog collections and R. Schaefer for assisting with the statistical analyses. Conducted in partial fulfillment of the requirements for the M.S. degree, Department of Zoology, Miami University, awarded to T. Muir. Experimental procedures were approved by the Institutional Animal Care and Use Committee of Miami University (protocol 629). Supported by the National Science Foundation Grant IOB 0416750 to J. Costanzo.
References
Arad Z (2001) Desiccation and rehydration in land snails—a test for distinct set points in Theba pisana. Isr J Zool 47:41–53
Baldwin RF, Calhoun AJK, DeMaynader PG (2006) Conservation planning for amphibian species with complex habitat requirements: a case study using movements and habitat selection of the wood frog Rana sylvatica. J Herpetol 40:442–453
Balinsky JB (1981) Adaptation of nitrogen metabolism to hypertonic environment in Amphibia. J Exp Zool 215:335–350
Chew SF, Chan NKY, Loong AM, Hiong KC, Tam WL, Ip YK (2004) Nitrogen metabolism in the African lungfish (Protopterus dolloi) aestivating in a mucus cocoon on land. J Exp Biol 207:777–786
Churchill TA, Storey KB (1993) Dehydration tolerance in wood frogs: a new perspective on development of amphibian freeze tolerance. Am J Physiol 265:R1324–R1332
Costanzo JP, Lee RE (2005) Cryoprotection by urea in a terrestrially hibernating frog. J Exp Biol 208:4079–4089
Costanzo JP, Lee RE, Lortz PH (1993) Physiological responses of freeze-tolerant and -intolerant frogs: clues to evolution of anuran freeze tolerance. Am J Physiol 265:R721–R725
Cowan KJ, Storey KB (2002) Urea and KCl have different effects on enzyme activities in liver and muscle of estivating versus nonestivating species. Biochem Cell Biol 80:745–755
Edwards JR, Jenkins JL, Swanson DL (2004) Seasonal effects of dehydration on glucose mobilization in freeze-tolerant chorus frogs (Pseudacris triseriata) and freeze-intolerant toads (Bufo woodhousii and B. cognatus). J Exp Zool 301A:521–531
Flanigan JE, Withers PC, Guppy M (1991) In vitro metabolic depression of tissues from the aestivating frog Neobatrachus pelobatoides. J Exp Biol 161:273–283
Fuery CJ, Attwood PV, Withers PC, Yancey PH, Baldwin J, Guppy M (1997) Effects of urea on M4-lactate dehydrogenase from elasmobranchs and urea-accumulating Australian desert frogs. Comp Biochem Physiol B 117:143–150
Garcia-Romeu F, Masoni A, Isaia J (1981) Active urea transport through isolated skins of frog and toad. Am J Physiol 241:114–123
Gatten REJ (1987) Activity metabolism of anuran amphibians: tolerance to dehydration. Physiol Zool 60:576–585
Gil N, Katz U (1996) Oxygen consumption, heart rate and respiratory movements are maintained almost unchanged in toads (Bufo viridis) on soil without access to free water. J Arid Environ 33:237–245
Grundy JE, Storey KB (1994) Urea and salt effects on enzymes from estivating and non-estivating amphibians. Mol Cell Biochem 131:9–17
Hand SC, Somero GN (1982) Urea and methylamine effects on rabbit muscle phosphofructokinase. J Biol Chem 257:734–741
Hillman SS (1978a) The roles of oxygen delivery and electrolyte levels in the dehydrational death of Xenopus laevis. J Comp Physiol 128:169–175
Hillman SS (1978b) Some effects of dehydration on internal distributions of water and solutes in Xenopus laevis. Comp Biochem Physiol 61:303–307
Hillman SS (1987) Dehydrational effects on cardiovascular and metabolic capacity in two amphibians. Physiol Zool 60:608–613
Hochachka PW, Somero GN (2002) Biochemical adaptation. Oxford University Press, New York
Jørgensen CB (1997a) 200 years of amphibian water economy: from Robert Townson to the present. Biol Rev 72:153–237
Jørgensen CB (1997b) Urea and amphibian water economy. Comp Biochem Physiol A 117:161–170
Kaji DM, Lim J, Shilkoff W, Zaidi W (1998) Urea inhibits the Na–K pump in human erythrocytes. J Membr Biol 165:125–131
Katz U, Hoffman J (1990) Changing plasma osmolality—a strategy of adaptation in anuran amphibia to water scarcity under burrowing conditions. In: Hanke W (eds) Biology and physiology of amphibians. Gutav Fischer Verlag, New York, pp 350–356
Kennett R, Christian K (1994) Metabolic depression in estivating long-neck turtles (Chelodina rugosa). Physiol Zool 67:1087–1102
Layne JR Jr, Lee RE (1987) Freeze tolerance and the dynamics of ice formation in wood frogs (Rana sylvatica) from southern Ohio. Can J Zool 65:2062–2065
Layne JR, Rice ME (2003) Postfreeze locomotion performance in wood frogs (Rana sylvatica) and spring peepers (Pseudacris crucifer). Can J Zool 81:2061–2065
Loong AM, Hiong KC, Lee SML, Wong WP, Chew SF, Ip YK (2005) Ornithine-urea cycle and urea synthesis in African lungfishes, Protopterus aethiopicus and Protopterus annectens, exposed to terrestrial conditions for six days. J Exp Zool 303A:354–365
McDonald MD, Smith CP, Walsh PJ (2006) The physiology and evolution of urea transport in fishes. J Membr Biol 212:93–107
Pinder AW, Storey KB, Ultsch GR (1992) Estivation and hibernation. In: Feder ME, Burggren WW (eds) Environmental physiology of the amphibians. The University of Chicago Press, Chicago, pp 250–274
Preest MR, Brust DG, Wygoda ML (1992) Cutaneous water loss and the effects of temperature and hydration state on aerobic metabolism of canyon treefrogs, Hyla arenicolor. Herpetologica 48:210–219
Rees BB, Hand SC (1993) Biochemical correlates of estivation tolerance in the mountainsnail Oreohelix (Pulmonata: Oreohelicidae). Biol Bull 184:230–242
Regosin JV, Windmiller BS, Reed JM (2003) Terrestrial habitat use and winter densities of the wood frog (Rana sylvatica). J Herpetol 37:390–394
Schmid WD (1965) Some aspects of the water economy of nine species of amphibians. Ecology 46:261–269
Secor SM (2005) Physiological responses to feeding, fasting and estivation for anurans. J Exp Biol 208:2595–2608
Shoemaker VH (1964) The effects of dehydration on electrolyte concentrations in a toad, Bufo marinus. Comp Biochem Physiol 13:261–271
Shoemaker VH, Hillman SS, Hillyard SD, Jackson DC, McClanahan LL, Withers PC, Wygoda ML (1992) Exchange of water, ions, and respiratory gases in terrestrial amphibians. In: Feder ME, Burggren WW (eds) Environmental physiology of the amphibians. The University of Chicago Press, Chicago, pp 125–150
Shpun S, Hoffman J, Katz U (1992) Anuran Amphibia which are not acclimable to high salt, tolerate high plasma urea. Comp Biochem Physiol A 103:473–477
Somero GN (1986) Protons, osmolytes, and fitness of internal milieu for protein function. Am J Physiol 251:R197–R213
Spaans EJA, Baker JM (1996) The soil freezing characteristic: its measurement and similarity to the soil moisture characteristic. Soil Sci Soc Am J 60:13–19
Spotila JR, O’Connor MP, Bakken GS (1992) Biophysics of heat and mass transfer. In: Feder ME, Burggren WW (eds) Evironmental physiology of the amphibians. The University of Chicago Press, Chicago, pp 59–80
Vleck D (1987) Measurement of O2 consumption, CO2 production, and water vapor production in a closed system. J Appl Physiol 62:2103–2106
Williams JB, Lee RE (2005) Plant senescence cues entry into diapause in the gall fly Eurosta solidaginis: resulting metabolic depression is critical for water conservation. J Exp Biol 208:4437–4444
Withers PC, Guppy M (1996) Do Australian desert frogs co-accumulate counteracting solutes with urea during aestivation? J Exp Biol 199:1809–1816
Wray S, Wilkie DR (1995) The relationship between plasma urea levels and some muscle trimethylamine levels in Xenopus laevis: a 31P and 14N Nuclear magnetic resonance study. J Exp Biol 198:373–378
Wright PA (1995) Nitrogen excretion: three end products, many physiological roles. J Exp Biol 198:273–281
Yancey PH, Burg MB (1990) Counteracting effects of urea and betaine on colony-forming efficiency of mammalian cells in culture. Am J Physiol 258:R198–R204
Yancey PH, Clark ME, Hand SC, Bowlus RD, Somero GN (1982) Living with water stress: evolution of osmolyte systems. Science 217:1214–1222
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by H.V. Carey.
Rights and permissions
About this article
Cite this article
Muir, T.J., Costanzo, J.P. & Lee, R.E. Osmotic and metabolic responses to dehydration and urea-loading in a dormant, terrestrially hibernating frog. J Comp Physiol B 177, 917–926 (2007). https://doi.org/10.1007/s00360-007-0190-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00360-007-0190-3