
Abstract
The physiological signal for torpor initiation appears to be related to fuel availability. Studies on metabolic fuel inhibition in placental heterotherms show that glucose deprivation via the inhibitor 2-deoxy-D-glucose (2DG) initiates a torpor-like state, whereas fatty acid deprivation via mercaptoacetate (MA) does not. As previous studies using inhibitors were limited to quantifying body temperature in placentals, we investigated whether inhibition of glucose or fatty acids for cellular oxidation induces torpor in the marsupial hibernator Cercartetus nanus, and how the response of metabolic rate is related to body temperature. Glucoprivation initiated a torpor-like state in C. nanus, but animals had much higher minimum body temperatures and metabolic rates than those of torpid food-deprived animals and arousal rates were slower. Moreover, 2DG-treated animals were thermoregulating at ambient temperatures of 20 and 12 °C, whereas food-deprived torpid animals were thermo-conforming. We suggest that glucoprivation reduces the hypothalamic body temperature set point, but only by about 8 °C rather than the approximately 28 °C during natural torpor. Reduced fatty acid availability via MA also induced a torpor-like state in some C. nanus, with physiological variables that did not differ from those of torpid food-deprived animals. We conclude that reduced glucose availability forms only part of the physiological trigger for torpor initiation in C. nanus. Reduced fatty acid availability, unlike for placental heterotherms, may be an important cue for torpor initiation in C. nanus, perhaps because marsupials lack functional brown adipose tissue.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Torpor is used by many heterothermic mammals to reduce energy expenditure during periods of limited food availability and/or cold stress (Boyer and Barnes 1999). During torpor, body temperature (T b), metabolic rate (MR) as well as other physiological functions are reduced, thereby lowering energy requirements. During entry into torpor, the set point for T b (T set) which is regulated in the hypothalamus, is lowered (Heller et al. 1977) resulting in a substantial reduction of MR which, in turn causes a fall in T b. Therefore over a wide range of T b, torpid animals are thermo-conforming thus no energy is required for thermoregulation and T b approaches ambient temperature (T a). The fall of T b, and in some species, metabolic inhibition, lowers MR well below basal MR (BMR; Geiser 1988). However, heterothermic endotherms are capable of thermoregulating T b above a species-specific T set during torpor and increasing T b via endogenous heat production at the end of a torpor bout.
While patterns of torpor have been described for many species, the physiological signals that trigger torpor onset and the role of different metabolic fuels during torpor are poorly understood. Many species enter torpor when food availability is limited; torpor is generally more pronounced in depth and duration when animals are food deprived (Tucker 1962; Geiser and Baudinette 1985; Geiser 1988; Geiser and Broome 1991; Song et al. 1998). This suggests torpor initiation is related to metabolic fuel availability.
Observations of the respiratory quotient (RQ) for a number of species suggest that during torpor, lipids are the predominant metabolic fuel (Snapp and Heller 1981; Willis 1982; Nestler 1990; Heldmaier et al. 1999). This is also true for arousal in some species (Nizielski et al. 1989; Heldmaier et al. 1999). Glucose is mainly metabolised during normothermia and, in some species, arousal (Snapp and Heller 1981; Nestler 1990). Evidence for a reduced use of glucose during torpor is supported by the decrease of plasma glucose concentrations prior to and during torpor in golden-mantled ground squirrels, Spermophilus lateralis, dormice, Eliomys quercinus, deer mice, Peromyscus maniculatus, and Siberian hamsters, Phodopus sungorus (Twente and Twente 1967; Atgie et al. 1990; Nestler 1991; Dark et al.1999; Heldmaier et al. 1999).
The role of different fuels has also been investigated by manipulating fuel availability using metabolic inhibitors (Dark et al. 1994, 1996; Dark and Miller 1998). 2-Deoxy-D-Glucose (2DG) is a non-metabolisable glucose analogue, which disrupts cellular glycolysis (Wick et al. 1957). Glucoprivation by 2DG has produced mixed results in placental heterotherms. 2DG induced a torpor-like state in Phodopus sungorus (Dark et al. 1994, 1996; Stamper et al. 1999). In contrast, Syrian hamsters (likely Mesocricetus auratus; Schneider et al. 1993) and Peromyscus maniculatus (Stamper and Dark 1997) did not substantially lower T b following 2DG-administration. S. lateralis also did not enter torpor after 2DG-treatment, but in naturally torpid individuals, 2DG induced arousal and slowed arousal rates (Dark and Miller 1998).
Whereas 2DG is effective in inducing a torpor-like state in Phodopus sungorus, the deprivation of fatty acids via the inhibitor mercaptoacetate (MA) did not reduce T b (Dark et al. 1994, 1996; Stamper et al. 1999). Similar observations were made after MA administration for S. lateralis (Dark and Miller 1998), and Peromyscus maniculatus (Stamper and Dark 1997). Moreover, fatty acid deprivation via MA initiated arousal in naturally torpid S. lateralis (Dark and Ruby 1993; Dark and Miller 1998). To our knowledge the only induction of a torpor-like state using a fatty acid inhibitor occurred in Syrian hamsters (likely M. auratus), but in this case methyl palmoxirate instead of MA was used (Schneider et al. 1993).
All previous studies investigating the effects of metabolic fuel deprivation on thermoregulation were restricted to T b measurements without quantifying MR. As the reduction of T b induced by glucose deprivation may not represent torpor, but rather some other hypothermic state (Dark et al.1999), interrelations between T b and MR should provide a more direct understanding of the physiological state induced by metabolic fuel deprivation. Moreover, all studies regarding metabolic fuel deprivation to date have been conducted on placental mammals, which possess functional brown adipose tissue (BAT) (Nicol et al. 1997), an important thermogenic tissue not found in marsupials. Thus the two mammalian suborders may differ in fuel use.
Therefore our purpose was to evaluate the effect of metabolic fuel deprivation in a marsupial hibernator, the eastern pygmy-possum (Cercartetus nanus; body mass 15–38 g) a nocturnal, arboreal marsupial. This species is nectarivorous/insectivorous (Hume 1999), fattens extensively in captivity (40–90 g; Bartholomew and Hudson 1962) and can hibernate for months without food at low T a at any time of the year (Geiser 1993; Song et al. 1997). Although bouts of torpor in C. nanus can last for up to 1 month, this species also displays short bouts of torpor at high T a and at the beginning of a hibernation period. However, even during short bouts of torpor, MR is reduced to levels similar to those during hibernation (Geiser 1993; Song et al. 1997). We determined whether the glucose inhibitor, 2DG and the fatty acid inhibitor, MA affect T b and MR during thermoregulation in C. nanus. We used these data to examine whether physiological variables induced by fuel deprivation differ from placental mammals, and differ from those during torpor induced by food deprivation in C. nanus. These results may allow us to shed new light as to the potential mechanism underlying the hypothermic response to metabolic fuel deprivation.
Materials and methods
Animals
The adult C. nanus used in this study were originally collected from nest boxes on the Northern Tablelands, NSW (30°22′S, 152°45′E), but had been maintained at the University of New England for between one and several years and were accustomed to handling and respirometry measurements. Animals were housed in groups (3–4) in large aviaries (1.5×0.8×1.8 m). Each aviary contained wooden nest boxes with shredded paper and branches for climbing (details in Westman and Geiser 2003).
The aviaries were enclosed inside a room with large open windows, therefore animals were exposed to the natural photoperiod of Armidale, NSW. The T a during the experimental period (autumn and winter) was in the range of 5–25 °C, buffered relative to outside temperature, similar to a forest (Körtner et al. 2001). Animals were fed daily from a variety of foods including high protein iron-enriched baby cereal (Heinz) mixed with honey and water, (supplemented once per week with a multi-vitamin and mineral supplement), fresh apple, honey water, mealworms, and oats. Sunflower seeds, walnuts, and almonds were provided occasionally. Water was available ad libitum.
Body temperature
We measured T b using small (1.5 g) temperature-sensitive transmitters (Minimitter Model X-M, ±0.1 °C). Transmitter pulse rate was calibrated to the nearest 0.1 °C against a precision mercury thermometer in a water bath between 2 °C and 40 °C. Transmitters were implanted intraperitoneally under oxygen/Forthane anaesthesia, after which animals were allowed to recover for at least 14 days. The transmitter signal was received by ferrite rod antennae and multiplexed to an AM receiver, and the pulse rate interval was used to calculate T b.
Metabolic rate
MR was measured as the rate of oxygen consumption. Animals were placed into 0.75-l respirometry chambers in a temperature-controlled cabinet (±0.5 °C). Three animal channels and one reference channel (outside air) were scanned with solenoid valves. Each channel was read for 3 min in sequence, thereby giving one reading per animal every 12 min. The flow-rate of 400 ml/min was controlled with rotameters and measured with a mass flowmeter (FMA-5606, Omega, Stamford). Oxygen content of dry air leaving the animal chambers was measured with a single channel oxygen analyser (Ametek Applied Electrochemistry S-3A/1, Pittsburgh) fitted with a high-resolution output board (80335SE).
The T a inside the respirometers was measured to the nearest 0.1 °C with a digital thermocouple thermometer (DP116, Omega).
Analog outputs from flowmeter, O2 analyser, digital thermometer, and receiver were interfaced to a personal computer via a 14-bit A/D card. MR, T b and T a were measured simultaneously.
Animals were weighed before and after respirometry measurements and a linear decrease in body mass was assumed for calculation of mass-specific MR according to Eqn. 3a of Withers (1977).
From measurements of oxygen consumption, RMR was determined as the mean of the three (i.e. over 36 min) lowest consecutive values when animals were normothermic and at rest. Minimum MR was calculated as for RMR, but was taken when animals had a stable T b of less than 30 °C.
Metabolic inhibitors
The glucose oxidation inhibitor, 2DG (Sigma, Australia, 98% purity) was injected at three dose levels: 800, 1600 and 2400 mg/kg body mass. MA (Sigma, Australia, 99% purity) blocks fatty acid oxidation by interfering with mitochondrial membrane transport of fatty acids (Bauché et al. 1983). Doses of MA used were 600, 900 and 1,200 μmol/kg body mass. These dose levels were selected because they produced significant results in other similar-sized heterotherms (Dark et al. 1994, 1996; Stamper et al. 1999). The inhibitors were dissolved in saline to make an injectable volume of <0.2 ml per animal. Control animals were given an equivalent amount of saline. All injections were administered subcutaneously in the intrascapular area. Animals were injected only once per week.
Procedures
Adult male and female C. nanus (mean body mass 36.7 ± 5.4 g) were administered each dose of 2DG and saline in weekly intervals in a lattice square design. Injections were administered between 0800 hours and 0900 hours, during their rest phase when natural torpor is typically expressed (Song et al. 1997). After injection, animals were placed into respirometry chambers for approximately 8 h or longer if they had not aroused from torpor by this time; O2 consumption, T b and T a were measured simultaneously. Each treatment was conducted at both T a 20 °C (n=8) and T a 12 °C (n=6). The same procedure was followed for MA. Each MA treatment was also conducted at both T a 20 °C (n=6) and T a 12 °C (n=7). To compare torpor induced by the inhibitors and natural torpor induced by food deprivation (FD), animals were placed into respirometry chambers in the late afternoon (1600 hours) for approximately 22–24 h. C. nanus usually enters torpor in the second part of the night or early hours of the morning, after a nocturnal period of activity. Food and water were not available in the respirometry chambers.
Torpor
Animals were considered to be torpid when T b was below 30 °C and MR was less than 75% of the RMR for normothermic animals at the same T a. Minimum T b was calculated as the mean of the lowest consecutive T b readings over at least 24 min. Fastest entry and arousal rates were measured as degrees Celsius per minute and were calculated as the most rapid change in T b over a 12-min interval.
Statistics
All data are presented as mean ±1 SD for 'n', the number of individuals. Frequency of animals with T b<30 °C was analysed using χ2 analysis. Normothermic variables were compared by Oneway Repeated Measures ANOVA (ORM ANOVA), followed by Tukey's pairwise comparisons (P<0.05). Incidence of animals with T b<30 °C varied with treatment and so physiological variables of animals with T b<30 °C were compared using General Linear Models (GLM), followed by Tukey's pairwise comparisons (P<0.05). Linear regressions were fitted using the method of least squares.
Results
Glucose oxidation inhibitor, 2DG
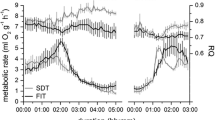
Temporal patterns of MR and T b in C. nanus differed among treatments, but were similar at both T as (representative examples at T a 20 °C in Fig. 1). FD C. nanus entered torpor during the scotophase, characterised by a decrease in MR and T b (Fig. 1a). Torpor was maintained for several hours with T b close to T a. Arousal occurred in the afternoon, indicated by a rapid increase in MR with an overshoot above resting MR, which rapidly increased T b. Both variables then returned to normothermic resting levels. 2DG significantly altered temporal patterns of MR and T b (Fig. 1b); MR and T b were reduced immediately after injection. However, MR was maintained at higher levels than those for FD animals and consequently T b was much higher than T a. Duration of the 2DG-induced hypothermic state was generally shorter than for FD torpor. During arousal from the 2DG-induced state, MR increased gradually and excluded a significant MR overshoot; this resulted in a slow rewarming process (Fig. 1b). Once T b and MR had returned to normothermic resting levels they did not differ from the FD or saline-treated animals (T a 20 °C: MR, P=0.54 T b, P=0.16; T a 12 °C: MR, P=0.96 T b, P=0.09; ORM ANOVA). Saline treatment did not reduce T b or MR at either T a (Fig. 1c). After injecting saline, animals expressed normothermic resting levels of MR and T b for the entire measurement period. Body mass of animals did not differ among treatments (T a 20 °C: P=0.08; T a 12 °C: P=0.95; ORM ANOVA).

Examples of temporal fluctuations of body temperature (T b ; open circles) and metabolic rate (MR; closed circles) for individual Cercartetus nanus after food deprivation (FD) (a), 2-deoxy-D-glucose (2DG) 2,400 mg/kg (b), saline treatment at T a 20 °C (c). The dark bars represent the scotophase and the asterisks represent time of injection
Administration of 2DG reduced T b at both T a 20 °C and T a 12 °C. The proportion of animals with a T b<30 °C was significantly affected by treatment (T a 20 °C: χ 2=21.33, df=4, P=0.0001; T a 12 °C: χ 2=26.61, df=4, P=0.0001; not shown). At 1,600 mg/kg 2DG, 100% of animals, and at 2,400 mg/kg 2DG >80% of animals reduced T b<30 °C at both T as. At 800 mg/kg 2DG, only 38% of animals reduced T b<30 °C at T a 20 °C and none at T a 12 °C. Saline did not decrease T b<30 °C in any animal. In contrast, during FD, 75% of animals entered torpor at T a 20 °C and 100% entered torpor at T a 12 °C.
The minimum T b of C. nanus with T b<30 °C was affected by treatment (Fig. 2a). FD C. nanus had significantly lower minimum T bs than animals from all 2DG treatments at both T a 20 °C (P=0.0001; GLM) and T a 12 °C (P=0.0001; GLM). T b in FD individuals was on average 6 °C lower at T a 20 °C and 14 °C lower at T a 12 °C than for 2DG treatment. Interestingly, for 2DG-treated individuals with T b<30 °C, the minimum T b did not differ among 2DG doses (P>0.05; Tukey), and at all 2DG doses the minimum T b was similar between T as (P=0.63; ANOVA) with a mean T b of 26.1±2.6 °C.

a Mean minimum T b and b mean minimum MR of C. nanus with T b<30 °C after three doses of 2DG and FD treatments at ambient temperature (T a) 20 °C (dark bars) and T a 12 °C (light bars). Error bars show SD. Numbers in parentheses show number of animals. Animals treated with saline did not reduce T b<30 °C
Minimum mass-specific MR of C. nanus with T b<30 °C was also affected by treatment (Fig. 2b). At both T as, 2DG-treated animals had significantly higher minimum MR (six times higher at T a 20 °C and nine times higher at T a 12 °C) than FD animals, reflecting a smaller T b−T a differential in the latter (P=0.0001; GLM; both T as). Similar to minimum T b, there was no significant difference in minimum MR among 2DG doses at either T a. However, minimum MR for 2DG-treated animals was about double at T a 12 °C compared to T a 20 °C.
The duration of time with T b<30 °C was significantly affected by treatment at both T a 20 °C and 12 °C (P<0.005; GLM; Fig. 3). FD animals maintained T b<30 °C longer than any of the 2DG-treated animals (T a 20 °C, about 3–7 times and at T a 12 °C, about 3–4 times longer). However, duration for 2DG-treated animals with T b<30 °C, was similar at all doses (P>0.05; Tukey) and between T a (P=0.57; ANOVA).

Duration of time C. nanus had T b<30 °C after three doses 2DG and FD treatments at T a 20 °C (dark bars) and T a 12 °C (light bars). Error bars show SD. Numbers in parentheses show number of animals. Animals treated with saline did not reduce T b<30 °C
The fastest rate of T b decrease was significantly affected by treatment at T a 20 °C (P=0.001; GLM), but not at T a 12 °C (P=0.559; GLM). At T a 20 °C, the decline in T b was faster in animals treated with the two highest 2DG doses than for FD animals or those treated with the lowest 2DG dose (P<0.05; Tukey).
The fastest rate of T b increase was affected by treatment at both T as (P=0.0001; GLM both T as; Fig. 4). At T a 20 °C, arousal rate of FD animals was about 3–4 times faster than for all 2DG treatments and at T a 12 °C arousal rate of FD animals was 5–7 times faster. There was no significant effect of dose among 2DG treatments (P>0.05; Tukey), and arousal rate of all 2DG-treated animals was also not significantly different between T a (P=0.49; ANOVA).

Mean maximum rate of T b increase in C. nanus with a T b<30 °C after three doses 2DG, and FD treatments at T a 20 °C (dark bars) and T a 12 °C (light bars). Error bars show SD. Numbers in parentheses show number of animals. Animals treated with saline did not reduce T b<30 °C
Fatty acid oxidation inhibitor, MA
Temporal patterns of MR and T b were affected by the inhibition of fatty acid oxidation in one of two ways (examples at T a 12 °C in Fig. 5). Immediately after injection either: (i) MR and T b were decreased for several hours, followed by a rapid arousal which was comparable to FD animals (Fig. 5b), or (ii) animals remained normothermic with MR and T b at resting values throughout the measurement period, which did not differ significantly from those of saline treated animals (T a 20 °C: MR P=0.82; T b P=0.21; T a 12 °C: MR P=0.48, T b P=0.24; GLM; Fig. 5c). Saline treatment also did not reduce T b or MR in any animal. Body mass was also not significantly different among treatments (T a 20 °C: P=0.45, T a 12 °C: P=0.30, ORM ANOVA).

Examples of temporal fluctuations of T b (open circles) and MR (closed circles) for individual C. nanus at T a 12 °C after food deprivation (a), and mercaptoacetate (MA) 1,200 μmol/kg (b, c). The dark bars represent the scotophase and the asterisks represent time of injection
Administration of MA reduced T b in some C. nanus at both T a 20 °C and T a 12 °C. The proportion of animals with a T b<30 °C varied significantly with treatment (T a 20 °C: χ 2=14.66, df=4, P=0.005; T a 12 °C: χ 2=21.05, df=4, P=0.0001). At T a 20 °C, 17% of animals reduced T b<30 °C at 600 μmol/kg MA, and 33% at 900 μmol/kg MA, but none at the highest MA dose. At T a 12 °C, 29% of animals reduced T b<30 °C at 600 μmol/kg MA dose, and 43% at 1,200 μmol/kg MA, but none at the 900 μmol/kg dose. The highest frequency of T b<30 °C occurred for FD animals, with 83% reducing T b<30 °C at T a 20 °C and 100% at T a 12 °C. Again no animals reduced T b when given saline alone. Animals which responded to MA-treatment by reducing T b<30 °C comprised only two individuals out of six at T a 20 °C, and three individuals of seven at T a 12 °C.
Of those C. nanus that reduced T b<30 °C, the minimum T b reached was not affected by treatment at either T a (T a 20 °C: P=0.90; T a 12 °C: P=0.45; GLM; Fig. 6a). Therefore there was no significant difference in minimum T b among MA doses or between MA-treated and FD animals.

a Mean minimum T b and b mean minimum MR of C. nanus with T b<30 °C after three doses MA and FD treatments at T a 20 °C (dark bars) and T a 12 °C (light bars). Error bars show SD. Numbers in parentheses show number of animals. Scale as for Fig. 2 for better comparison. Animals treated with saline did not reduce T b<30 °C
Minimum mass-specific MR of C. nanus with T b<30 °C was also not affected by MA treatment (T a 20 °C: P=0.19; T a 12 °C: P=0.13; GLM; Fig. 6b). Minimum MR of MA-treated animals were similar among MA treatment and between MA-treated and FD animals at both T as (P>0.05; Tukey).
Duration of time with T b<30 °C was significantly affected by treatment at T a 12 °C (P=0.01; GLM), but not at T a 20 °C (P=0.65; GLM). At T a 12 °C, duration of T b<30 °C in MA-treated animals was about half that of FD animals, but this reduced duration reflects that FD animals entered torpor during the scotophase whereas MA treatment did not occur until the morning.
Maximum rates of T b decrease and increase did not differ between treatments at either T a. Therefore maximum entry and arousal rates of FD animals and MA-treated animals were not different.
Discussion
Our study shows that metabolic inhibition of glucose and fatty acid oxidation does affect thermoregulation in the heterothermic marsupial C. nanus. Similar to the placental heterotherm Phodopus sungorus (Dark et al. 1994; Stamper et al. 1999), 2DG caused a significant reduction of T b when administered during the rest phase, suggesting that glucose may be an important energy substrate for normothermic thermoregulation. However, unlike for most placental heterotherms (Dark et al. 1994; Stamper and Dark 1997; Dark and Miller 1998), inhibition of fatty acid oxidation with MA also resulted in a reduction of T b in some C. nanus.
Glucose oxidation inhibitor
Reduced glucose availability for cellular oxidation by the inhibitor 2DG had several effects on thermoregulation in C. nanus. The most notable of these was a substantial decrease of MR and T b immediately after treatment. These reductions were clearly a response to glucoprivation as animals treated with saline under the same conditions did not reduce MR or T b. Interestingly, all 2DG treatments resulted in a reduction of T b to a mean minimum of about 26 °C regardless of dose or T a. Although more animals decreased T b after treatment with the highest 2DG doses, the minimum T b of individuals with T b<30 °C was not dose dependent. Therefore the minimum T b does not appear to depend on how much glucose was deprived but merely that it was deprived. Nevertheless, minimum T bs of 2DG-treated animals were maintained well above the T as used in this study. This was also reported for male hamsters (Phodopus sungorus) treated with 2DG (Dark et al. 1996). As for minimum T b, there was no dose effect on minimum MR, so all 2DG doses resulted in similar mean minimum MR at the same T a. However, minimum MR of 2DG-treated animals was affected by T a, with MR increasing with decreasing T a.
Decreases of MR and T b caused by 2DG treatment may indicate that torpor is initiated in response to glucoprivation. However comparisons of physiological variables for the 2DG-induced state and torpid FD animals reveal considerable differences. In contrast to 2DG treatment, the minimum T b of torpid FD animals approached T a as has previously been shown for this species (Geiser 1993; Song et al. 1997). Minimum MR of FD animals was much lower than that of 2DG-treated animals. Importantly, minimum MR of 2DG-treated animals increased significantly with decreasing T a, whereas FD animals did not (Fig. 7). Therefore, torpid FD animals conserved considerably more energy than 2DG-treated animals. The posture of 2DG-treated and FD animals with T b<30 °C also differed. FD animals curled up in a spherical shape with their heads and tails tucked under their bodies, a posture typical of hibernating mammals. In contrast, 2DG-treated animals did not adopt this posture; they remained outstretched with heads raised and often looked around. If both treatments (glucoprivation and FD) result in torpor, why do the physiological responses differ?

Mass-specific MR of C. nanus as a function of T a using different thermal strategies. Normothermic resting animals (closed circles; taken from Song et al. 1997), minimum MR of 2DG-treated animals with T b<30 °C (open circles), non-regulating torpid FD animals (closed squares) and non-regulating torpid MA treated animals (open squares). Regression equations were: (1) MR=3.689−0.106T a, r 2=55.7%, P=0.0001 (closed circles), (2) MR=2.822−0.099T a, r 2=67.3%, P=0.0001 (open circles), (3) MR=0.235−0.005T a, r 2=8.7%, P=0.352 (closed squares), and (4) MR=0.395−0.009T a, r 2=13%, P=0.380 (open squares). The dashed line demonstrates the response of MR to decreasing T a of thermoconforming C. nanus over a wide T a range and in contrast the solid line shows the proportional increase of MR with decreasing T a below T set in thermoregulating C. nanus (from Song et al. 1997). Therefore 2DG-treated animals show the same response of proportional thermoregulation with decreasing T a as normothermic animals and torpid animals below T set. MA-treated animals show the same response of thermoconforming as FD animals
The increase in minimum MR with decreasing T a demonstrates that in the measured T a range, 2DG-treated animals were thermoregulating in order to maintain T b, although at a reduced level (Fig. 7). This suggests that glucose deprivation by 2DG reduces the hypothalamic T set as occurs in natural torpor (Heller et al. 1977). However T set is reduced by only approximately 8 °C. In contrast, during natural torpor in C. nanus, T set is lowered by approximately 28 °C (Song et al.1997) and only when T a falls below about 6 °C, proportional thermoregulation is initiated to maintain T b during steady state torpor (Fig. 7). Both T a 20 °C and 12 °C fall below the T set of 2DG-treated animals and thus proportional thermoregulation is initiated to maintain T b at approximately 26 °C. This response is similar to normothermic C. nanus at T as below the thermal neutral zone, where MR increases with decreasing T a to maintain a high stable T b of about 34 °C (Fig. 7). Due to the relatively high T set of 2DG-treated animals, MR was only about 20% lower than that of normothermic thermoregulating animals at the same T a. Conversely, T set of torpid FD C. nanus was below the T as of 20 °C and 12 °C. These individuals were thermoconforming, T b approximated T a, and MRs were 95% lower than normothermic animals at the same T a. Based on these results, it is plausible to expect that 2DG-treated C. nanus at T as above 26 °C would also be thermoconforming and would exhibit a relatively greater reduction of MR, but the high T a and consequently high T b would prevent the reduction of MR to the values of FD animals at a much lower T b (Song et al. 1997; present study).
The lack of response to an increased dose of 2DG on MR and T b suggests that 2DG does not affect these variables via metabolic fuel inhibition, but rather via an influence on the hypothalamic T set. Injection of 2DG into specific sites in both the anterior and posterior hypothalamus of rats (Shiraishi and Mager 1980a) resulted in a reduction of T b, suggesting that the hypothalamus is sensitive to glucose levels. When 2DG was administered peripherally to rats heat production fell and T b was reduced by up to 3 °C (Shiraishi and Mager 1980b), returning to normal levels after several hours.
Our results suggest that C. nanus responds to 2DG by decreasing T b via a shift in T set, but that the shift is more pronounced than in homeothermic rats. As T set was not lowered to that usually seen in torpid C. nanus, reduced glucose availability likely plays only a part in torpor initiation. While we cannot preclude that higher 2DG doses than measured here may further decrease T set, the lack of response to a 3-fold increase in dose rate makes this scenario unlikely.
The duration of torpor for 2DG-treated animals was much shorter than that of torpid FD animals. This is because FD animals entered torpor during the scotophase whereas 2DG treatment did not begin until the morning, yet it is interesting that 2DG-treated animals did not maintain torpor longer by arousing later in the day. All 2DG-treated animals aroused at similar times in the afternoon. As there was no apparent effect of dose or T a on torpor duration in 2DG-treated animals this may reflect the length of activity of 2DG. The T b of 2DG-treated animals increased at a very slow rate with no overshoot of MR as is observed in FD animals. Dark and Miller (1998) also observed slow rewarming in torpid S. lateralis treated with 2DG and suggested that glucose must be required during arousal, a conclusion supported by RQ measurements (Willis 1982; Snapp and Heller 1981; Nestler 1990). Once 2DG is removed from the body, T set may increase slowly back to a normothermic level and heat production fuelled by available glucose increases slowly to increase T b. Slower arousal rates with 2DG treatment has been suggested to be due to the suppression of sympathetic nerve activity to interscapular BAT in rats (Egawa et al. 1989). However this interpretation is unlikely in marsupials because they do not possess functional BAT (Nicol et al. 1997).
Fatty acid oxidation inhibitor
Lipoprivation via MA also reduced MR and T b substantially in some C. nanus, but the incidence of T b reduction of MA-treated animals varied with dose and T a and only occurred in a few of the same individuals at all doses. The reason for this variability is not known as it was not related to gender, age, or BM, but may be due to individual variation in fatty acid levels and/or types prior to MA treatment. All animals received the same type of food the evening prior to treatment, however, as animals were maintained in small groups (3–4), it is likely that not all animals consumed the same amount of food, which may have influenced the levels of circulating metabolic fuels.
In contrast to 2DG, minimum T b and MR of MA-treated animals that did reduce T b<30 °C were similar to those of torpid FD animals (Fig. 7) and MA did not slow the rate of T b increase during arousal. Given that the physiological variables of MA-treated and torpid FD animals were similar, it appears that lipoprivation may play a role in torpor induction in C. nanus. It appears that T set is reduced to the same level observed during natural torpor or at least below T a 12 °C, as MA-treated animals were thermoconforming and had minimum MR similar to torpid FD animals (Fig. 7). The posture of MA-treated and FD animals with T b<30 °C was similar with all animals adopting the posture typical of hibernating mammals as described previously.
Previous studies involving reduced fatty acid availability due to MA did not observe torpor induction in placental heterotherms (Dark et al. 1994; Stamper and Dark 1997; Dark and Miller 1998; Stamper et al. 1999). However, MA-treatment reduced whole blood glucose concentrations in Phodopus sungorus (Dark et al. 1994) and Spermophilus lateralis (Dark and Miller 1998), suggesting increased metabolism of glucose in response to a reduction in fatty acid availability for oxidation. Why some C. nanus initiated torpor specifically in response to MA treatment while other species do not is unknown, but may reflect differences in fuel use. Previous studies of the effects of MA-treatment have been conducted on placental heterotherms which, unlike marsupials, posses functional BAT (Nicol et al. 1997) and it is possible our results reflect this difference. Torpor may be inhibited in placental heterotherms when fatty acid availability is low because non-shivering thermogenesis used during arousal relies on lipolysis from BAT (Nedergaard and Cannon 1984) whereas marsupials rely heavily on shivering (Dawson 1989), which requires glucose metabolism.
Torpor initiation due to decreased fatty acid availability in C. nanus differs from observations of naturally torpid golden-mantled ground squirrels (S. lateralis), which aroused in a dose-dependent manner when treated with MA (Dark and Miller 1998). At the highest MA dose 40% S. lateralis remained torpid despite the large reduction of available fatty acids for oxidation, perhaps because the dose rate used was limited to 1,200 μmol/kg. In C. nanus, individual variation in fatty acid or carbohydrate levels prior to MA treatment may reflect the different responses to MA treatment.
C. nanus may enter torpor when fatty acid availability is decreased to conserve remaining lipid supplies. Reduced food availability requires reliance on stored lipids and the reduction of lipid stores to a critical threshold level may signal torpor initiation. High lipid stores may actually inhibit torpor in some marsupials, since administration of leptin reduces torpor use in Sminthopsis macroura (Geiser et al. 1998), but this species enters daily torpor exclusively rather than hibernation.
Conclusions
Reduced availability of both glucose and fatty acids induced a torpor-like state in the marsupial C. nanus. Reduced glucose availability may form part of the trigger for torpor initiation in this species, as it is predominately nectarivorous and may be highly dependent on glucose metabolism. Therefore reduced glucose availability when food is restricted may be an initial trigger that acts on the hypothalamus at torpor onset. However, during entry into natural torpor, T b falls well below that caused by glucose deprivation and consequently other mechanisms must be involved.
Decreased fatty acid availability also induced a torpor-like state in some C. nanus, which was similar to FD torpor suggesting similar underlying metabolic signals. Access to lipid-rich food in the form of insects is likely to be low in winter when nectar availability can be temporarily high, and accumulation of lipid stores therefore requires synthesis from sugars. Thus, at times of low food availability, the decrease in lipid stores may act as a signal for entry into deep torpor.
Abbreviations
- BAT :
-
brown adipose tissue
- BMR :
-
basal metabolic rate
- 2DG :
-
2-deoxy-D-glucose
- FD :
-
food deprived
- GLM :
-
general linear models
- MA :
-
mercaptoacetate
- MR :
-
metabolic rate
- RQ :
-
respiratory quotient
- T a :
-
ambient temperature
- T b :
-
body temperature
- T set :
-
body temperature set point
References
Atgie C, Nibbelink M, Ambid L (1990) Sympathoadrenal activity and hypoglycemia in the hibernating garden dormouse. Physiol Behav 48:783–787
Bartholomew GA, Hudson JW (1962) Hibernation, estivation, temperature regulation, evaporative water loss, and heart rate of the pigmy possum, Cercaertus nanus. Physiol Zool 35:94–107
Bauché F, Saourault D, Giudicelli Y, Nordmann J, Nordmann R (1983) Inhibition in vitro of acyl-CoA dehydrogenases by 2-mercaptoacetate in rat liver mitochondria. Biochem J 215:457–464
Boyer BB, Barnes BM (1999) Molecular and metabolic aspects of mammalian hibernation. BioScience 49:713:124
Dark J, Ruby NF (1993) Metabolic fuel utilization during hibernation. In: Carey C, Florant GL, Wunder BA, Horwitz B (eds) Life in the cold: ecological, physiological, and molecular mechanisms. Westview, Boulder, pp 167–174
Dark J, Miller DR (1998) Metabolic fuel privation in hibernating and awake ground squirrels. Physiol Behav 63:59–65
Dark J, Miller DR, Zucker I (1994) Reduced glucose availability induces torpor in Siberian hamsters. Am J Physiol 267:R496–R501
Dark J, Miller DR, Licht P, Zucker I (1996) Glucoprivation counteracts effects of testosterone on daily torpor in Siberian hamsters. Am J Physiol 270:R398–R403
Dark J, Lewis DA, Zucker I (1999) Hypoglycemia and torpor in Siberian hamsters. Am J Physiol 276:R776–R781
Dawson TJ (1989) Responses to cold of monotremes and marsupials. In: Wang LCH (ed) Advances in comparative and environmental physiology 4. Springer, Berlin Heidelberg New York
Egawa M, Yoshimatsu H, Bray GA (1989) Effects of 2-deoxy-D-glucose on sympathetic nerve activity to interscapular brown adipose tissue. Am J Physiol 257:R1377–R1385
Geiser F (1988) Reduction of metabolism during hibernation and daily torpor in mammals and birds: temperature effect of physiological inhibition? J Comp Physiol B 158:25–37
Geiser F (1993) Hibernation in the eastern pygmy possum, Cercartetus nanus (Marsupialia: Burramyidae). Aust J Zool 41:67–75
Geiser F, Baudinette RV (1985) The influence of temperature and photophase on daily torpor in Sminthopsis macroura (Dasyuridae: Marsupialia). J Comp Physiol B 156:129–134
Geiser F, Broome LS (1991) Hibernation in the mountain pygmy possum Burramys parvus (Marsupialia). J Zool (Lond) 223:593–602
Geiser F, Körtner G, Schmidt I (1998) Leptin increases energy expenditure of a marsupial by inhibition of daily torpor. Am J Physiol 275:R1627–R1632
Heldmaier G, Klingenspor M, Werneyer M, Lampi BJ, Brooks SP, Storey KB (1999) Metabolic adjustments during daily torpor in the Djungarian hamster. Am J Physiol 276:E896–E906
Heller HC, Colliver GW, Beard J (1977) Thermoregulation during entrance into hibernation. Pflügers Arch 369:55–59
Hume I (1999) Marsupial nutrition. Cambridge University Press, Cambridge
Körtner G, Brigham RM, Geiser F (2001) Torpor in free-ranging tawny frogmouths (Podargus strigoides) Physiol Biochem Zool 74:789–797
Nedergaard J, Cannon B (1984) Preferential utilization of brown adipose tissue lipids during arousal from hibernation in hamsters. Am J Physiol 247:R506–R512
Nestler JR (1990) Relationships between respiratory quotient and metabolic rate during entry to and arousal from daily torpor in deer mice (Peromyscus maniculatus). Physiol Zool 63:504–515
Nestler JR (1991) Metabolic substrate change during daily torpor in deer mice. Can J Zool 69:322–327
Nicol SC, Pavlides D, Andersen NA (1997) Nonshivering thermogenesis in marsupials: absence of thermogenic response to β3-adrenergic agonists. Comp Biochem Physiol A 117:399–405
Nizielski SE, Billington CJ, Levine AS (1989) Brown fat GDP binding and circulating metabolites during hibernation and arousal. Am J Physiol 257:R536–R541
Schneider JE, Friedenson DG, Hall AJ, Wade GN (1993) Glucoprivation induces anestrus and lipoprivation may induce hibernation in Syrian hamsters. Am J Physiol 264:R573–R577
Shiraishi T, Mager M (1980a) Hypothermia following injection of 2-deoxy-D-glucose into selected hypothalamic sites. Am J Physiol 239:R265–R269
Shiraishi T, Mager M (1980b) 2-deoxy-D-glucose-induced hypothermia: thermoregulatory pathways in rat. Am J Physiol 239:R270–R276
Snapp BD, Heller HC (1981) Suppression of metabolism during hibernation in ground squirrels (Citellus lateralis). Physiol Zool 54:297–307
Song X, Körtner G, Geiser F (1997) Thermal relations of metabolic rate reduction in a hibernating marsupial. Am J Physiol 273:R2097–R2104
Song X, Körtner G, Geiser F (1998) Temperature selection and use of torpor by the marsupial Sminthopsis macroura. Physiol Behav 64:675–682
Stamper JL, Dark J (1997) Metabolic fuel availability influences thermoregulation in deer mice (Peromyscus maniculatus). Physiol Behav 61:521–524
Stamper JL, Dark J, Zucker I (1999) Photoperiod modulates torpor and food intake in Siberian hamsters challenged with metabolic inhibitors. Physiol Behav 66:113–118
Tucker VA (1962) Diurnal torpidity in the Californian pocket mouse. Science 136:380–381
Twente JW, Twente JA (1967) Concentrations of D-glucose in the blood of Citellus lateralis after known intervals of hibernating periods. J Mamm 48:381–386
Westman W, Geiser F (2003) Captive breeding, growth, and development of thermoregulation in the eastern pygmy-possum (Cercartetus nanus) In: Goldingay RL, Jackson SM (eds) The biology of Australian possums and gliders. Southern Cross University Press, Lismore (In Press)
Wick AN, Drury DR, Nakada HI, Wolfe JB (1957) Localization of the primary metabolic block produced by 2-Deoxyglucose. J Biol Chem 224:963–969
Willis JS (1982) Intermediary metabolism in hibernation. In: Lyman CP, Willis JS, Malan A, Wang LCH (eds) Hibernation and torpor in mammals and birds. Academic Press, New York, pp 124–139
Withers PC (1977) Measurement of V̇O2, V̇CO2, and evaporative water loss with a flow-through mask. J Appl Physiol 42:120–123
Acknowledgements
We thank S. Cairns for advice on statistics and M. Brigham, N. Christian, R. Drury and C. Turbill for critically reading the manuscript. This study was financially supported by a UNE postgraduate research grant to W. Westman and a UNE research grant and ARC grant to F. Geiser. All experiments were carried out with approval from the University of New England Animal Ethics Committee and under licence from New South Wales National Parks and Wildlife Service.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by I.D. Hume
Rights and permissions
About this article
Cite this article
Westman, W., Geiser, F. The effect of metabolic fuel availability on thermoregulation and torpor in a marsupial hibernator. J Comp Physiol B 174, 49–57 (2004). https://doi.org/10.1007/s00360-003-0388-y
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00360-003-0388-y