
Abstract
Chemosensory information is crucial for most insects to feed and reproduce. Olfactory signals are mainly used at a distance, whereas gustatory stimuli play an important role when insects directly contact chemical substrates. In noctuid moths, although the antennae are the main olfactory organ, they also bear taste sensilla. These taste sensilla detect sugars and hence are involved in appetitive learning but could also play an important role in food evaluation by detecting salts and bitter substances. To investigate this, we measured the responses of individual taste sensilla on the antennae of Spodoptera littoralis to sugars and salts using tip recordings. We also traced the projections of their neuronal axons into the brain. In each sensillum, we found one or two neurons responding to sugars: one NaCl-responsive and one water-sensitive neuron. Responses of these neurons were dose-dependent and similar across different locations on the antenna. Responses were dependent on the sex for sucrose and on both sex and location for glucose and fructose. We did not observe a spatial map for the projections from specific regions of the antennae to the deutocerebrum or the tritocerebrum/suboesophageal ganglion complex. In accordance with physiological recordings, back-fills from individual sensilla revealed up to four axons, in most cases targeting different projection zones.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The antennae of moths and other insects are multimodal sensory appendages bearing several types of cuticular sensilla innervated by sensory receptor neurons. The most prominent sensillum type on moth antennae contains receptor neurons detecting odorants involved in either intra-specific communication, in finding host plants or in searching food over a distance. In most insects, including moths, other sensillum types on the antennae house receptor neurons that detect gustatory and mechanosensory information as well as temperature and humidity (Altner et al. 1977). Moths, like many other insects, move their antennae actively to detect position, shape, mechanical texture and chemical identity of surrounding objects during various behaviours such as walking/searching for food sources, escaping from enemies, mate recognition and other intra-specific communication (Nishino et al. 2005). Taste and smell are essential for detecting food, mates and noxious stimuli in the environment. Whereas olfaction allows insects to discriminate between a large number of different odours in many different combinations, taste is more elementary allowing only the identification of a few categories of tastants (Masek and Scott 2010).
Gustatory receptor neurons are housed in taste sensilla and their dendrites are exposed to the environment through a single opening at the hair tip. Each insect taste sensillum usually contains four contact chemosensory neurons and one mechanosensory neuron (Singh 1997). Taste sensilla can be broadly distributed on the body surface of insects (mouthparts, tarsi, antennae, wings and ovipositor), where they are involved in eliciting various behaviours (Dethier 1976; Chapman 1982; Stocker 1994; Dahanukar et al. 2005). Taste neurons have historically been categorized as a function of the major stimulus category to which they respond. Gustatory neurons from the tarsi, the antennae and the mouthparts respond to sugars, and their activation elicits feeding responses in many insects such as proboscis extension reflexes (Menzel and Müller 1996; Fan et al. 1997), whereas activation of the taste neurons on the ovipositor of females may generate or inhibit egg-laying behaviour (Renwick 1989; Calas et al. 2007). On the other hand, the detection of aversive/repellent compounds, such as salts or secondary plant compounds (i.e. steroids, alkaloids), is important for the inhibition of feeding behaviour (Schoonhoven and van Loon 2002; Meunier et al. 2003b). There are indications in both vertebrates and invertebrates that primary taste modalities might be represented in separate zones within the central nervous system (Newland 1999; Wang et al. 2004; Marella et al. 2006; Accolla et al. 2007), but this zonation in spatial patterning is not as distinct as that observed in olfaction.
All antennal sensilla send their axons to the brain via the antennal nerve. The deutocerebrum receives the majority of the antennal sensory inputs (Rospars 1988; Homberg et al. 1989). There are two main parts of the deutocerebrum: the antennal lobe (AL) and ventral and adjacent to it, the antennal mechanosensory and motor centre (AMMC) (Homberg et al. 1989). The AL receives all olfactory information (Hansson and Anton 2000) and odours are represented as a functional map within the distinct compartments, the glomeruli of the antennal lobe (Rospars 1983; Hildebrand and Shepherd 1997; Vosshall et al. 2000; Galizia and Menzel 2001; Heisenberg 2003).
The AMMC neuropil does not appear to be organized into glomeruli or other distinct subdivisions and its boundaries with the surrounding areas cannot easily be defined (Kloppenburg et al. 1997). This region receives projections of the antennal mechanosensory axons in locusts, cockroaches, honeybees and moths (Homberg et al. 1989; Kloppenburg 1995) but also axons from contact chemosensory sensilla (Nishino et al. 2005; Jørgensen et al. 2006). Some antennal afferents pass the AMMC region with or without giving rise to arborisations within it and project further into the tritocerebrum/suboesophageal ganglion (SOG) region (Nishino et al. 2005; Jørgensen et al. 2006), and in some insects further into the thoracic ganglia (Kent and Hildebrand 1987; Barrozo et al. 2009). In addition to receiving afferents from the antennae, the SOG receives inputs from the gustatory receptor neurons of the mouthparts (Edgecomb and Murdock 1992; Mitchell et al. 1999; Kvello et al. 2006).
However, little is known about the functional mapping of chemosensory afferents in general, and about the pattern and distribution of projections of taste neurons from the antennae in the central nervous system. The projection areas described so far for presumptive gustatory afferents from the antennae are the AMMC and the tritocerebrum/SOG (Nishino et al. 2005; Jørgensen et al. 2006, 2007; Haupt 2007). There is some evidence in moths for a central organization according to the location of the taste sensilla on different head appendages: taste neurons from the proboscis in Heliothis virescens show a parallel finger-like projection pattern into the SOG antero-medially to the antennal gustatory neurons (Jørgensen et al. 2006). Last, in flies, the SOG also receives projections from thoracic afferents (Edgecomb and Murdock 1992) indicating that in addition to being involved in mapping taste information from antennal and mouth appendages, the SOG plays a role in central integration of gustatory information from different parts of the body. However, it is not known if sensilla on different parts of the antenna have different functional characteristics and thus might project differentially within the central nervous system.
The present paper describes responses of gustatory receptor neurons housed in contact chemosensory sensilla (sensilla chaetica) on different parts of the antennae of Spodoptera littoralis. First, their responses to sugars and salts are investigated to know whether antennal taste sensilla are homogeneous with respect to the stimuli tested. Second, the projections of these gustatory receptor neurons into the central nervous system are described, to identify possible differences with respect to antennal location (somatotopic map) or to modality (chemotopic map).
Materials and methods
Insects
Spodoptera littoralis moths were reared on a semi-artificial medium in our laboratory (Poitout and Bues 1974). Larvae were reared individually from the third to last instar. Male and female pupae were kept separately in groups of 40–60 at 20–24 °C, 55–75 % relative humidity, 16:8-h light:dark cycle. Male and female adult moths were kept separately in plastic containers and provided with a 10 % sucrose solution to feed ad libitum.
Scanning electron microscopy
The antennae were cut and air dried in a dessicator. The dried samples were mounted horizontally with double-sided tape with a conductive glue on a standard aluminium stub. Samples were coated successively with carbon and gold/palladium before examination in a scanning electron microscope (JEOL JSM 840).
Chemicals
Sucrose, glucose, fructose, potassium chloride, sodium chloride and choline chloride were purchased from Sigma-Aldrich (France). Compounds were diluted in ultra pure water and 1 mM KCl was added in the sugar solutions to ensure the conductivity of the stimulating solution. All solutions were kept at 4 °C and used at concentrations ranging from 0.1 to 100 mM.
Electrophysiology
Insect preparation and recording
Moths were constrained in a styrofoam block, with the head protruding over a flat surface. Each antenna was tightened to the support with tungsten hooks, wax and sticky tape. The insect was grounded using electrocardiogram gel deposited on the antenna (Redux® Gel, Parker Laboratories Inc., Fairfield, USA), which served as a bridge to a silver wire connected to the ground. Recordings presented here were all done from lateral sensilla. To stimulate a sensillum, the tip of the hair was covered during 2 s with an electrode containing the stimulus and in addition 1 mM KCl as an electrolyte for sugar solutions (Hodgson et al. 1955). Electrodes were made from borosilicate glass capillary tubes (GC-100 T, Clark, USA) pulled in two steps so that the tip had a diameter of about 40–60 μm (PC-10 vertical puller, Narishige, Japan). This electrode was directly fitted on a silver wire connected to a probe (TasteProbe DT-02, Syntech, Germany) mounted on a micromanipulator, and advanced under visual control (MZ12, Leica, France). The taste probe includes a contact detector that triggers offset compensation (Marion-Poll and Van der Pers 1996). Electric signals were further amplified and filtered (CyberAmp 320, Axon Instrument, USA; gain 1,000; 8th order Bessel pass-band filter: 1–2,800 Hz). Contacting a taste hair with the stimulus electrode triggered data acquisition and storage on a disc, under the control of a custom software, dbWave (Marion-Poll 1996).
Stimulation and data analysis
We tested three to five homologous sensilla per preparation. As control, we used 1 mM KCl, present in all experiments as an electrolyte in the recording pipette, because it is the least stimulating salt. Each sensillum was stimulated with the substance under test, presented in an increasing order of concentrations (0.1, 1, 10 and 100 mM). We waited at least 1 min between consecutive stimulations of the same hair. Sensilla were sampled along the antennal flagellum at the base (segments 1–10), in the middle (segments 20–40) and at the tip (the 10 terminal segments: segments 56–66). For this study, we recorded responses from 70 males and 110 females.
For each recording, spikes were detected and analyzed using dbWave. We evaluated the responses by counting spikes during the first second of recording. Whenever possible, we sorted the spikes according to their amplitude and shape, as well as by taking into account spike superposition (Meunier et al. 2003a). Spike counts had a Poisson distribution and so were analyzed using generalized linear models (Poisson regression) using the statistical software SAS (Cary, NC, USA).
Cross-adaptation test
In order to verify if the same taste neuron responds both to sugars and salts we performed cross-adaptation tests as follows. The response to a first stimulus (100 mM NaCl) was evaluated over a group of lateral sensilla. Five minute later, one of these sensilla was capped with 100 mM sucrose for 5 min. Immediately after adaptation to sucrose, we recorded the response of this sensillum to sucrose (100 mM) and immediately after switching pipettes to the initial stimulus (100 mM NaCl).
Staining of gustatory receptor neurons
Insect preparation
The insects were placed in a plastic tube and the head was immobilized with wax. The antennae were left free for mass staining and attached to a wax platform with tungsten hooks for single sensillum staining.
Staining
Mass staining was performed on the flagellum of S. littoralis. The flagellum was cut distally, medially or proximally and the cut end was covered with a glass capillary filled with a 1 % Neurobiotin (SP-1120, Vector Laboratories, Inc., Burlingame, USA) solution in ultrapure water.
Staining of single sensilla was performed by cutting one sensillum chaeticum at about half its length with sharpened forceps in a pool made of Vaseline and filled with distilled water. After 6 min the water was removed and replaced by a 1 % solution of Neurobiotin. The preparations were placed in Petri dishes moistened with a wet piece of tissue and kept for 7–10 days in the refrigerator at 4 °C to allow diffusion of the dye. The brains were dissected and fixed in 4 % paraformaldehyde (over night at 4 °C) and rinsed in phosphate buffer (Millonig’s) containing 0.25 % Triton X. The brains were then dehydrated and rehydrated to make membranes more permeable. Subsequently, they were rinsed in buffer and incubated over night at 4 °C in Millonig’s buffer containing 0.25 % TritonX, 1 % bovine serum albumin and Oregon green–avidin conjugate (Oregon Green®, Invitrogen™, France). Finally, the preparations were rinsed in phosphate buffer and transferred in Vectashield medium (Vectashield® Mounting Medium, Vector Laboratories, ABCYS, France).
Confocal laser scanning microscopy
The projections of stained receptor neurons were examined with a confocal laser scanning microscope equipped with an Argon laser (Leica SP2 AOBS, Leica Microsystems Heidelberg, Germany) using a 10× dry objective. The brains were scanned in frontal plane with a 1.5-μm step size and 1,024 × 1,024 resolution for single staining and 3-μm step size and 512 × 512 resolution for mass staining.
Reconstructions
The optical sections were used for the creation of three-dimensional reconstructions using a custom-made program in Matlab® (Louise Couton, Kiên Kiêu, Jean-Pierre Rospars). Some neurons were manually reconstructed from confocal stacks to visualize details of arborisation patterns. Maximum projections of optical sections were obtained through stacks transferred to ImageJ software (NIH, USA).
Results
Distribution of contact chemoreceptive sensilla on the antennae
The antennal flagellum of S. littoralis is composed of about 70 flagellomeres with similar topographic organization. Each flagellomere carries scales and a few sensilla on the dorsal side and a large number of sensilla trichodea and other sensillum types on the ventral side. Six sensilla chaetica with gustatory function are regularly disposed around the flagellomere, approximately in the middle of its length; two of these sensilla are found on the ventral side (Fig. 1a, b), two are on the dorsal side (Fig. 1b) and one sensillum is found on each side, at the border between the ventral and the dorsal fields (Fig. 1a, b). The terminal flagellomere bears an additional crown of 6–8 sensilla chaetica at the very tip (Fig. 1c) and only two taste sensilla are found on the flagellomeres at the base of the flagellum. In total, there are thus ~400 sensilla chaetica distributed more or less evenly along the antennal flagellum of S. littoralis. All these sensilla looked similar with SEM observations and occasionally a terminal pore could be observed at high magnification. No sexual dimorphism was detected in the number and distribution of sensilla chaetica.

Different views of the flagellum of the female moth S. littoralis. a Ventral view showing four sensilla chaetica: two ventral and one lateral on each side. b Lateral view showing lateral, dorsal and ventral taste sensilla. c Tip of the antenna showing a crown of six sensilla chaetica. Scale bars 100 μm
Response characteristics of gustatory receptor neurons on the antenna
Upon contact with a stimulation pipette, gustatory receptor neurons within sensilla chaetica emitted action potentials between 0.8 and 2.6 mV on a baseline noise of about 0.05 mV peak-to-peak. In most sensilla, the responses to sucrose, NaCl, and choline chloride were strongly phasic, with an initial burst of spikes of 100–200 ms followed by a gradually declining activity (Figs. 2, 3, 4, 5, 6). This initial burst was different from the activation of the mechanoreceptor neurons, which only fired smaller amplitude action potentials when strong lateral movements were imposed to the recording electrode and were not elicited by touching the tip of a sensillum with the recording electrode. We can thus exclude that a mechanosensitive neuron was interfering with the responses of chemosensitive neurons. Glucose elicited responses in fewer sensilla (data not shown) and with lower spike rates than responses to sucrose and fructose (see Fig. 7).

Typical electrophysiological recordings of gustatory receptor neurons (GRNs) from a lateral sensillum in response to increasing concentrations of sucrose. Two GRNs are firing as illustrated by the different sizes of the spikes (see enlarged lower trace and super-imposed spike shapes to the left of the recording traces). Note the highly phasic responses. Vertical scale bar 2 mV, horizontal scale bar 200 ms

Typical electrophysiological recordings of a gustatory receptor neuron (GRN) from a lateral sensillum in response to increasing concentrations of NaCl. A single GRN could be identified, responding in a phasic–tonic manner. Superimposed spikes detected in the recording are shown to the left of the recording traces (spike duration 6 ms). Vertical scale bar 2 mV, horizontal scale bar 200 ms

Cross-adaptation experiment between sucrose and NaCl on a lateral taste sensillum. After adaptation to a long stimulation with sucrose a response to NaCl is still evident, indicating that sucrose and NaCl are detected by different gustatory receptor neurons within the sensillum. Superimposed spike shapes detected in the recording are shown to the left of the recording traces. Vertical scale bar 2 mV, horizontal scale bar 200 ms

Responses of lateral taste sensilla from the antennae to choline chloride [known not to be perceived by gustatory receptor neurons (GRNs)]. a Recordings of a GRN in response to increasing concentrations of choline chloride. Superimposed spike shapes are shown to the left of the recording traces. Vertical scale bar 2 mV, horizontal scale bar 200 ms. b Dose–response curve to choline chloride (N = 25) with decreasing responses to increasing concentrations, indicating that the neuron is sensitive to osmotic pressure (water cell). Error bars indicate the standard error

Electrophysiological recordings of a gustatory receptor neuron in a lateral sensillum stimulated with increasing concentrations of KCl. Superimposed spike shapes are shown to the left of the recording traces and indicate shorter spike duration with increasing KCl concentration. The number of action potentials however does not change when the concentration of the tested solution is increased, suggesting that none of the neurons present in the sensillum responds to KCl. Vertical scale bar 2 mV, horizontal scale bar 200 ms

Dose–response curves from lateral sensilla in response to sucrose, fructose, glucose, KCl and NaCl as a function of the sensilla position on the antenna. a Females (N sucrose = 28–71, N fructose = 23–43, N glucose = 22–71, N KCl = 22–52, N NaCl = 28–88, N control = 41–107). b Males [N sucrose = 19–29, N fructose = 6–38, N glucose = 8–28, N KCl = 7–35, N NaCl = 11–56, N control (1 mM KCl) = 20–67]. The range of N values is given for each data point and stimulus. Error bars indicate the standard error. There are no significant differences in GRN sensitivity between different parts of the antennae when comparing dose–response curves (see Table 1)
It was usually difficult to determine how many cells were active in a given recording because the amplitudes and shapes of the spikes were very similar. However, we could monitor spike superposition (indicating that spikes originated from two neurons) and in some sensilla, the spike amplitudes were different so that we could discriminate them without ambiguity. Using these criteria, two distinct neurons, rather than one responding neuron, were found to respond to the sugars sucrose (Fig. 2), fructose and glucose in about 30 % of the recordings. For NaCl, we observed only one responding neuron (Fig. 3). Cross-adaptation experiments showed that the neurons responding to sucrose are different from the neurons responding to NaCl (Fig. 4).
Another neuron type present in sensilla chaetica was revealed by stimulation with choline chloride, which is known not to be perceived by gustatory receptor neurons in insects (Rees 1970; Merivee et al. 2004). We assume that increasing osmotic pressure by increasing the concentration of choline chloride led to decreasing numbers of action potentials in the recorded neuron (Fig. 5), corresponding to the responses of a “water cell”, as described, e.g. in flies (Fujishiro et al. 1984; Meunier et al. 2003b; Cameron et al. 2010). KCl did often not elicit dose-dependent responses in the neurons of the recorded sensilla, but we cannot exclude that antagonistic effects of salt- and water-sensitive neurons are responsible for this effect, because we could not differentiate between individual neurons (Figs. 6, 7; Table 1).
Sensitivity of taste sensilla across the antenna
The sensilla in males and females responded in a similar way to most of the substances we tested (Poisson regression: 3-way interaction, sex × substance × concentration: χ 22 = 5.72, P = 0.126, see also Table 1). To test potential differences in responses of gustatory receptor neurons as a function of the position of the recorded sensillum along the antenna, we compared the responses of lateral sensilla chaetica sampled at the base, at the middle and at the tip of the antenna for both sexes to all concentrations of each substance. At each location (tip, middle, base), the responses of the sensilla were dose-dependent for sucrose, fructose and NaCl, but not for glucose or KCl (Table 1). Males and females exhibited some variation in the intensity of the responses (Fig. 7).
To test whether there was an influence of sex on the intensity of the response at each location, we performed separate analyses for each substance (Table 2). The sensilla of females responded with a higher average rate of spiking than males towards all the sugars (sucrose, fructose and glucose, see also Fig. 7). For sucrose, the rate of spiking was on average greater for females but did not depend on the location of the sensilla on the antennae. For fructose and glucose, however, the rate of spiking depended on both sex and the location of the sensillum (see also Fig. 7). For NaCl, the rate of spiking in the sensilla of females was on average slightly greater but it did not depend on location. KCl did not have a strong effect on the rate of spiking in either males or females and responses were not dependent on location (Table 2; Fig. 7).
Central projections of non-olfactory antennal afferents
By applying neurobiotin on 73 cut antennae, 39 successfully stained preparations were obtained in which axons reaching the brain through the antennal nerve were visible. Of these 39 successfully stained preparations, 14 out of 25 attempts were from the base of the antenna, 19 out of 24 attempts from the middle of the antenna and 6 out of 24 attempts from the tip (Table 3). Some axons, presumably from olfactory neurons, projected into the antennal lobes, the others bypassed the AL postero-laterally and projected ipsilaterally in four areas.
The first area (A1) is situated in the deutocerebrum posterior to the antennal lobes and close to the oesophagus (Fig. 8a, b, d). These axons leave the axon bundle coming from the antennal nerve before it reaches the AMMC region. In most of the preparations, we observed a fork-like shape of the axon branches in area A1 (Fig. 8a). The second projection area (A2) is the AMMC, which is known to be the primary centre for the processing of mechanosensory information from the antennae (Fig. 8a, b, d). Some axons arborised within the AMMC and then projected postero-medially into the SOG as a finger-like projection with very few branches, targeting a third projection area (A3) (Fig. 8b, c, d). The fourth area (A4) concerned the axons leaving the AMMC with very few or no arborisations and projecting postero-medially into the SOG/tritocebrum region. These axons give rise to dense arborisations dorso-laterally to the endings of the “finger-like” projections in area 4 (Fig. 8c, d). In one preparation this type of neuron was found to send one branch to the deutocerebrum in the area situated postero-medially to the AL (data not shown). In two preparations where the proximal part of the connectives to the thoracic ganglia was present, we could observe one axon going further from the SOG to the thoracic ganglia (data not shown).

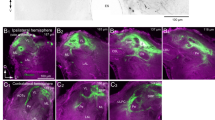
Optical sections and schematic representation of the S. littoralis brain (parafrontal view) with mass stained afferences from the antennal nerve. a Receptor neuron projections in the posterior part of the deutocerebrum, underneath the antennal lobe (AL) (area A1) and in the antennal motor and mechanosensory centre (AMMC, area A2). b Massively stained receptor neurons in the glomeruli of the antennal lobe, the AMMC, and within the medial suboesophageal ganglion (SOG, area A3). c Massively stained receptor neurons, showing projections to the medial SOG and the posterior SOG/tritocerebrum (area A4). d Schematic representation of all projection areas in a frontal view. d dorsal, l lateral. Scale bars100 μm. Z indicates depth of optical sections. Note that images originate from different preparations with slightly different orientations
We compared the mass stainings obtained by cutting the antennae at different levels. No obvious correlation of the projection patterns according to the location of the stained sensilla on the flagellum could be found (Table 3).
Neuron projections originating from individual taste sensilla
Twenty-eight successful preparations were obtained from 77 attempts to stain individual sensilla. Twelve of the successful preparations contained more than five stained neurons suggesting that more than one sensillum was stained, probably due to damages that occurred during the manipulations. One to four neurons could be identified in 16 preparations in which single sensilla were stained. The axons of these neurons ran tightly together when leaving the antennal nerve, bypassing the AL. In eight preparations (out of 16), one axon left the others before entering the AMMC and projected into the postero-medial part of the deutocerebrum, posterior to the AL (area A1 defined in the previous section) (Figs. 9a, 10).

Manual reconstructions of the central projections of individual axons from antennal sensillum chaeticum of S. littoralis in a frontal view. Insets show partial projections of optical sections from the area indicated in each square. a Axon projecting to the deutocerebrum in an area located posterior to the antennal lobe and close to the oesophagus (A1). b Axon projecting to the SOG region (A3) after giving rise to arborisations in the antennal mechanosensory and motor centre area (A2). c Axon projecting into the SOG/tritocerebrum area (A4). Note varicosities on all axonal branches. Oe oesophagus, SOG suboesophageal ganglion. Scale bars 75 μm

Confocal micrographs of axon projections of individual gustatory receptor neurons in S. littoralis and 3D reconstructions of the target areas. a Section of a brain showing projections in the AMMC, medial SOG and in the SOG/tritocerebrum (maximum projection of the sections of the posterior 150 μm of the brain). Scale bar 75 μm. b Three-dimensional reconstruction of the preparation in a, frontal view. c Projection of a 85-μm stack of the anterior part of a different brain, showing receptor neuron axons bypassing the antennal lobe (arrows). d Three-dimensional reconstruction of the preparation in c, frontal view. A1–A4 projection zones of antennal axons, d dorsal, l lateral, SOG suboesophageal ganglion. Scale bars 200 μm
Three other types of axons could be identified. A first type projected only into the AMMC (area A2) The second type showed massive arborisations within the AMMC and projected further postero-medially into the SOG (area A3) (Figs. 9b, 10). Its finger-like process gives rise to very few or no arborisations (Figs. 9b, 10). The third type of axons passed through the AMMC with very few or no arborisations inside and projected with widespread arborisations into the SOG/tritocerebrum covering a triangular-shaped area (A4) dorso-laterally to the finger-like projection area (Figs. 9c, 10). In individual preparations, never more than one axon terminated in areas A1, A3 and A4, whereas two axons terminated in about half of the preparations in area A2 and in three preparations even three axons were found to end in zone A2.
Discussion
Distribution of antennal taste sensilla
The number and distribution of sensilla chaetica in S. littoralis (about 400 sensilla) on the antennal flagellum are similar to what have been found in other Lepidoptera with filiform antennae like O. nubilalis (Cornford et al. 1973) and H. virescens (Jørgensen et al. 2006). In this work, we show that the sensilla chaetica in S. littoralis are contact receptors, responding to sugars and salts. We concentrated our electrophysiological recordings and backfills on lateral sensilla, which were the most accessible with the used techniques.
Functional segregation of receptor neurons is correlated with segregated central axon projections
Sensilla chaetica of insects typically house four taste neurons and one mechanoreceptor as shown by ultrastructural observations in other moths like Ephestia kuehniella (Anderson and Hallberg 1990; Chaika and Sinitsina 1997) or in Yponomeuta sp. (Cuperus 1985). The mechanoreceptive function is consistent with our SEM observations that sensilla chaetica have a basal articulated socket. It is also consistent with our electrophysiological recordings, which showed action potentials of an additional neuron when the hairs were moved laterally by the recording electrode (data not shown).
Although it was usually not possible to resolve individual spike types from each neuron in our recordings, co-occurrence of spikes from different neurons induced visible superposition of spikes. This criterion and cross-adaptation experiments lead us to propose that sugars and NaCl elicit action potentials in different neurons. In addition, our data suggest the existence of a water cell, responding to osmotic pressure changes, when stimulated with different doses of choline chloride and in some cases with KCl. A separation of the spikes of individual receptor neurons within a sensillum was unfortunately not possible and therefore interpretations have to be considered with caution. Cross-adaptation experiments using KCl and choline chloride would be necessary to solve this question. Recordings from sensilla chaetica in the noctuid moth H. virescens have shown responses to more than four different compounds in individual sensilla, but the spectrum of tested stimuli was wider than in our study and related compounds might be detected by the same neuron (Jørgensen et al. 2007). Our data correspond roughly to findings in Drosophila melanogaster, where each gustatory neuron encodes different categories of tastants and are therefore called sugar cell, water cell, and salt cell according to their best stimulus (Singh 1997). A water cell was initially described in Diptera (Evans and Mellon 1962; Wieczorek and Köppl 1978; Meunier et al. 2000, 2003b; Inoshita and Tanimura 2006) and was found in several orders of insects, including in the larvae (Schoonhoven and van Loon 2002) and adults (Chapman 2003) of phytophagous Lepidoptera.
Mass-fills from the antennae of S. littoralis allowed us to describe four distinct projection areas within the central nervous system (denoted here A1 to A4) in addition to the antennal lobe, without a possibility to distinguish between mechanosensory and gustatory fibres. In many other insects, axons from the mechanosensory neurons have a larger diameter than the fibres from chemosensory neurons. In gustatory sensilla on the legs of the locust and the fly, mechanosensory fibres have a larger diameter than chemosensory fibres and project in a somatotopic manner into the thoracic ganglia (Murphey et al. 1989; Newland et al. 2000). We suppose that mechanosensory fibres in S. littoralis project mainly to the AMMC/dorsal lobe area, as found in other Lepidoptera and in the cockroach (Camazine and Hildebrand 1979; Nishino et al. 2005). Mass stainings of the antennae in H. virescens have revealed two main projection areas for mechanosensory and chemosensory neurons: a fan-shaped region within the AMMC, corresponding to area A2 in S. littoralis and a finger-like projection within the dorsal SOG, corresponding to area A4 in our study. In some cases, however, a third projection within the ventral SOG, corresponding to area A3 in our study, has also been observed in H. virescens (Jørgensen et al. 2006). The mechanosensory neuron from individual sensilla in H. virescens could be identified because of a larger axon diameter than the gustatory fibres, and it often, but not exclusively, terminated within the AMMC (Jørgensen et al. 2006).
All projections resulting from antennal staining were restricted to the ipsilateral side of the brain, as shown in H. virescens (Jørgensen et al. 2006) and Periplaneta americana (Nishino et al. 2005). This pattern seems specific to antennal sensilla since projections from taste sensilla on other head appendages can also be contralateral, as shown for proboscis sensilla in H. virescens (Kvello et al. 2006) and labellar hairs in Phormia regina (Edgecomb and Murdock 1992).
Stainings from individual antennal sensilla revealed that gustatory neurons within the same sensillum in most cases project to separate areas and only one area (A2) received frequently the endings of more than one receptor neuron axon. This might indicate that zone A2 might receive not only mechanosensory fibres but eventually also axons from sugar-responding neurons, whereas neurons with other response profiles might project to the remaining three zones. This result is consistent with the hypothesis that each of the four areas gathers neurons with similar response characteristics, representing a form of chemotopic map. Gustatory neurons seem to target both a deutocerebral region and two areas within the tritocerebrum/SOG, indicating that antennal gustatory information is transmitted in parallel to the brain segment corresponding to the antennae, the deutocerebrum, and to the tritocerebrum/SOG, which receives also direct gustatory input from the mouthparts and from neurons passing through and branching in the thoracic ganglia (Jørgensen et al. 2006). In flies, one or more axons originating from taste sensilla on the legs ascend from the thoracic ganglia towards the brain and arborise in the same region of the SOG as neurons from labellar hairs (Murphey et al. 1989; Edgecomb and Murdock 1992). The tritocerebrum/SOG seems thus to serve not only as primary, but also as secondary integration centre for gustatory information from different parts of the body, as shown by intracellular recording and staining of central gustatory neurons within the SOG in H. virescens (Kvello et al. 2010).
Segregation of the gustatory receptor neuron projections as a function of the quality of the taste stimulus has been shown in different insects. In Drosophila, two antagonistically responding taste neurons have been described in the same sensillum (Hiroi et al. 2004). One is encoding the presence of attractive stimuli like sugars and salts at low concentrations while the second one responds to aversive stimuli like bitter compounds and salts at high concentration. Neurons detecting deterrent substances, situated in sensilla on the labellum, seem to project bilaterally in the tritocerebrum/SOG and neurons responding to phagostimulants project ipsilaterally into distinct areas (Thorne et al. 2004; Wang et al. 2004). These observations support the working hypotheses proposed by Chapman (2003) that phagostimulatory and deterrent neurons are the basic labelled lines of the insect taste receptor system and that these lines are represented in different areas of the CNS. These findings are confirmed by studies on central neurons processing information from contact chemoreceptive sensilla. Recordings from SOG interneurons in the fly Sarcophaga bullata showed that interneurons responding to NaCl stimulation of the labellar lobes did not respond to sucrose stimulation and vice versa (Mitchell and Itagaki 1992). In Locusta migratoria, neurons from the SOG responded both to chemical and mechanical stimulations but the time course of the responses to host plant versus non-host plant stimuli was different (Rogers and Simpson 1999).
Functionally uniform sensilla along the antenna give rise to uniform central projection patterns of sensory neurons
One of our objectives was to test if sensilla from different flagellomeres could be considered as functionally identical or if sensilla located at the tip would have different sensitivities, under the rationale that terminal sensilla are more likely to be involved in sampling tastants than those at the base of the antenna. Our current data support the first hypothesis because we did not find any compound-specific sensilla at a specific location or a clear gradient of sensitivity along the antenna. Nor did we find strong evidence that the responses of sensilla exhibited location-specific dose–responses: only males exhibited location-specific responses to glucose and fructose, with higher sensitivity at the base of the antenna. In accordance with the physiologically similar responses of taste neurons along the antenna, we did not discover differences in central projections of taste neurons originating from sensilla at different positions on the antenna. A more detailed analysis of axon branches within each projection area would, however, be necessary to detect or exclude somatotopic patterns at a finer scale.
In summary, the sensilla chaetica present on the antenna of S. littoralis adults are remarkably homogenous in their morphology, distribution and sensitivity across the length of the antenna. The absence of compound-specific sensillum types is possibly related to the limited number of substances we have used in this study. In a recent study, Jørgensen et al. (2007) have found several sensillum types responding to antifeedant molecules in H. virescens. Pilot experiments performed on S. littoralis did not allow us to find similar responses (A. Popescu, unpublished observations). While these experiments focused on food-related molecules, one should also consider non-volatile lipophilic molecules which are important either to assess the quality of a host plant (Udayagiri and Mason 1997; Powell et al. 1999; Grant et al. 2000; Steinbauer et al. 2004; Müller and Riederer 2005), or which are important in the context of mating, like cuticular molecules which can be either inhibitory (Lacaille et al. 2007) or excitatory aphrodisiacs.
Although our study indicates that the responses of the taste sensilla do not change much along the antenna, in H. virescens, it was reported that the number of sensilla chaetica responding to sucrose increases significantly from the base to the tip of the flagellum (Jørgensen et al. 2007). Our conclusions are based on recordings made from the lateral sensilla and it is possible that variations occur within the population of ventral or dorsal sensilla not sampled here. The second difference lies in the sample size: our observations are based on sampling 900 hairs over 180 insects while H. virescens observations rely on 132 sensilla from 11 moths. Further studies are needed to resolve this discrepancy.
Conclusions
While olfactory sensilla outnumber taste sensilla by at least a factor of 100 in S. littoralis, the number of taste sensilla on the antenna is not negligible. It represents a total of about 400 sensilla chaetica, i.e. 1,600 taste neurons and 400 mechanoreceptor neurons. It remains now to be seen how insects really use these sensilla and if they provide them enough information to discriminate many chemicals. That the lateral taste sensilla from each flagellomere are functionally almost identical militates against the hypothesis that this organ is used in fine discrimination of tastants. Buccal appendages and even legs stand in sharp contrast with the antennal taste system because (1) the density of their taste sensilla is higher and because (2) more variation has been found in the responses of sensilla from the same sensillar field to sugars (Liscia et al. 1998; Hiroi et al. 2002), salts (Maes and Den Otter 1976) and antifeedant molecules (Meunier et al. 2003b). We hypothesize that the antennal taste system is relatively primitive and is used as a rather general detection system indicating food or unpleasant/non-palatable tastants, passing the relay to other taste organs when finer discrimination tasks are necessary.
Our study reveals the target areas of gustatory and their associated mechanosensory neurons originating from antennal taste sensilla. Together with the broad knowledge on processing of olfactory stimuli, these data will be important for future research on integration of different sensory modalities in contexts such as host plant evaluation and appetitive or aversive learning, where olfactory, gustatory and mechanosensory stimuli interact to elicit specific behaviours. Although the honeybee has been the main model for research on appetitive learning associating olfactory and gustatory stimuli for a long time (for review see Menzel and Muller 1996), both aversive and appetitive learning paradigms are now well established in moths (Fan et al. 1997; Hartlieb et al. 1999; Skiri et al. 2005; Jørgensen et al. 2007) allowing to approach peripheral and central processing of multimodal signals.
References
Accolla R, Bathellier B, Petersen CCH, Carleton A (2007) Differential spatial representation of taste modalities in the rat gustatory cortex. J Neurosci 27:1396–1404
Altner H, Sass H, Altner I (1977) Relationship between structure and function of antennal chemoreceptive, hygroreceptive, and thermoreceptive sensilla in Periplaneta americana. Cell Tissue Res 176:389–405
Anderson P, Hallberg E (1990) Structure and distribution of tactile and bimodal taste tactile sensilla on the ovipositor, tarsi and antennae of the flour moth, Ephestia kuehniella (Zeller) (Lepidoptera, Pyralidae). Int J Insect Morphol Embryol 19:13–23
Barrozo RB, Couton L, Lazzari CR, Insausti TC, Minoli SA, Fresquet N, Rospars JP, Anton S (2009) Arthropod Struct Dev 38:101–110
Calas D, Berthier A, Marion-Poll F (2007) Do European corn borer females detect and avoid laying eggs in the presence of 20-Hydroxyecdysone? J Chem Ecol 33:1393–1404
Camazine SM, Hildebrand JG (1979) Central projections of antennal sensory neurons in mature and developing Manduca sexta. Soc Neurosci Abstr 5:155
Cameron P, Hiroi M, Ngai J, Scott K (2010) The molecular basis for water taste in Drosophila. Nature 465:91–95
Chaika SY, Sinitsina EE (1997) Antennal sensory organs of the flour moth Ephestia kuehniella Zell. (Lepidoptera: Pyralidae). Moscow Univ Biol Sci Bull 52:29–35
Chapman RF (1982) Chemoreception: the significance of receptor numbers. Adv Insect Physiol 16:247–333
Chapman RF (2003) Contact chemoreception in feeding by phytophagous insects. Annu Rev Entomol 48:455–484
Cornford ME, Rowley WA, Klun JA (1973) Scanning electron-microscopy of antennal sensilla of European corn borer, Ostrinia nubilalis Lepidoptera-Pyralidae. Ann Entomol Soc Am 66:1079–1088
Cuperus PL (1985) Ultrastructure of antennal sense organs of small ermine moths, Yponomeuta spp (Lepidoptera, Yponomeutidae). Int J Insect Morphol Embryol 14:179–191
Dahanukar A, Hallem EA, Carlson JR (2005) Insect chemoreception. Curr Opin Neurobiol 15:423–430
Dethier VG (1976) The hungry fly: a physiological study of the behavior associated with feeding. Harvard University Press, Cambridge
Edgecomb RS, Murdock LL (1992) Central projections of axons from taste hairs n the labellum and tarsi of the blowfly, Phormia regina Meigen. J Comp Neurol 315:431–444
Evans DR, Mellon D (1962) Electrophysiological studies of a water receptor associated with taste sensilla of the blowfly. J Gen Physiol 45:487–500
Fan RJ, Anderson P, Hansson BS (1997) Behavioural analysis of olfactory conditioning in the moth Spodoptera littoralis (Boisd.) (Lepidoptera: Noctuidae). J Exp Biol 200:2969–2976
Fujishiro N, Kijima H, Morita H (1984) Impulse frequency and action potential amplitude in the labellar chemosensory neurones of Drosophila melanogaster. J Insect Physiol 30:317–325
Galizia CG, Menzel R (2001) The role of glomeruli in the neural representation of odours: results from optical recording studies. J Insect Physiol 47:115–130
Grant GG, Zhao B, Langevin D (2000) Oviposition response of spruce budworm (Lepidoptera: Tortricidae) to aliphatic carboxylic acids. Environ Entomol 29:164–170
Hansson BS, Anton S (2000) Function and morphology of the antennal lobe: new developments. Annu Rev Entomol 45:203–231
Hartlieb E, Anderson P, Hansson BS (1999) Appetitive learning of odours with different behavioural meaning in moths. Physiol Behav 67:671–677
Haupt SS (2007) Central gustatory projections and side-specificity of operant antennal muscle conditioning in the honeybee. J Comp Physiol A 193:523–535
Heisenberg M (2003) Mushroom body memoir: from maps to models. Nature Rev Neurosci 4:266–275
Hildebrand JG, Shepherd GM (1997) Mechanisms of olfactory discrimination: converging evidence for common principles across phyla. Annu Rev Neurosci 20:595–631
Hiroi M, Marion-Poll F, Tanimura T (2002) Differentiated response to sugars among labellar chemosensilla in Drosophila. Zoolog Sci 19:1009–1018
Hiroi M, Meunier N, Marion-Poll F, Tanimura T (2004) Two antagonistic gustatory receptor neurons responding to sweet-salty and bitter taste in Drosophila. J Neurobiol 61:333–342
Hodgson ES, Lettvin JY, Roeder KD (1955) Physiology of a primary chemoreceptor unit. Science 122:417–418
Homberg U, Christensen TA, Hildebrand JG (1989) Structure and function of the deutocerebrum in insects. Annu Rev Entomol 34:477–501
Inoshita T, Tanimura T (2006) Cellular identification of water gustatory receptor neurons and their central projection pattern in Drosophila. Proc Natl Acad Sci USA 103:1094–1099
Jørgensen K, Kvello P, Almaas TJ, Mustaparta H (2006) Two closely located areas in the suboesophageal ganglion and the tritocerebrum receive projections of gustatory receptor neurons located on the antennae and the proboscis in the moth Heliothis virescens. J Comp Neurol 496:121–134
Jørgensen K, Almaas TJ, Marion-Poll F, Mustaparta H (2007) Electrophysiological characterization of responses from gustatory receptor neurons of sensilla chaetica in the moth Heliothis virescens. Chem Senses 32:863–879
Kent KS, Hildebrand JG (1987) Cephalic sensory pathways in the central nervous system of larval Manduca sexta (Lepidoptera: Sphingidae). Philos Trans R Soc London B Biol Sci 315:1–36
Kloppenburg P (1995) Anatomy of the antennal motoneurons in the brain of the honeybee (Apis mellifera). J Comp Neurol 363:333–343
Kloppenburg P, Camazine SM, Sun XJ, Randolph P, Hildebrand JG (1997) Organization of the antennal motor system in the sphinx moth Manduca sexta. Cell Tissue Res 287:425–433
Kvello P, Almaas TJ, Mustaparta H (2006) A confined taste area in a lepidopteran brain. Arthrop Struct Dev 35:35–45
Kvello P, Jørgensen K, Mustaparta H (2010) Central gustatory neurons integrate taste quality information from four appendages in the moth Heliothis virescens. J Neurophysiol 103:2965–2981
Lacaille F, Hiroi M, Twele R, Inoshita T, Umemoto D, Maniere G, Marion-Poll F, Ozaki M, Francke W, Cobb M et al (2007) An inhibitory sex pheromone tastes bitter for Drosophila males. PLoS One 2:e661
Liscia A, Majone R, Solari P, Barbarossa IT, Crnjar R (1998) Sugar response differences related to sensillum type and location on the labella of Protophormia terraenovae: a contribution to spatial representation of the stimulus. J Insect Physiol 44:471–481
Maes FW, Den Otter CJ (1976) Relationship between taste cell responses and arrangement of labellar taste setae in the blowfly Calliphora vicina. J Insect Physiol 22:377–384
Marella S, Fischler W, Kong P, Asgarian S, Reukhert E, Scott K (2006) Imaging taste responses in the fly brain reveals a functional map of taste category and behavior. Neuron 49:285–295
Marion-Poll F (1996) Display and analysis of electrophysiological data under MS-Windows. Entomol Exp Appl 80:116–119
Marion-Poll F, Van der Pers JNC (1996) Un-filtered recordings from insect taste sensilla. Entomol Exp Appl 80:113–115
Masek P, Scott K (2010) Limited taste discrimination in Drosophila. Proc Natl Acad Sci USA 107:14833–14838
Menzel R, Muller U (1996) Learning and memory in honeybees: from behavior to neural substrates. Annu Rev Neurosci 19:379–404
Merivee E, Renou M, Mänd M, Luik A, Heidemaa M, Ploomi A (2004) Electrophysiological responses to salts from antennal chaetoid taste sensilla of the ground beetle Pterostichus aethiops. J Insect Physiol 50:1001–1013
Meunier N, Ferveur JF, Marion-Poll F (2000) Sex-specific non-pheromonal taste receptors in Drosophila. Curr Biol 10:1583–1586
Meunier N, Marion-Poll F, Lansky P, Rospars JP (2003a) Estimation of the individual firing frequencies of two neurons recorded with a single electrode. Chem Senses 28:671–679
Meunier N, Marion-Poll F, Rospars JP, Tanimura T (2003b) Peripheral coding of bitter taste in Drosophila. J Neurobiol 56:139–152
Mitchell BK, Itagaki H (1992) Interneurons of the subesophageal ganglion of Sarcophaga bullata responding to gustatory and mechanosensory stimuli. J Comp Physiol A 171:213–230
Mitchell BK, Itagaki H, Rivet MP (1999) Peripheral and central structures involved in insect gustation. Microsc Res Tech 47:401–415
Müller C, Riederer M (2005) Plant surface properties in chemical ecology. J Chem Ecol 31:2621–2651
Murphey RK, Possidente D, Pollack G, Merritt DJ (1989) Modality-specific axonal projections in the CNS of the flies Phormia and Drosophila. J Comp Neurol 290:185–200
Newland PL (1999) Processing of gustatory information by spiking local interneurons in the locust. J Neurophysiol 82:3149–3159
Newland PL, Rogers SM, Gaaboub I, Matheson T (2000) Palrallel somatotopic maps of gustatory and mechanosensory neurons in the central nervous system of an insect. J Comp Neurol 425:82–96
Nishino H, Nishikawa M, Yokohari F, Mizunami M (2005) Dual, multilayered somatosensory maps formed by antennal tactile and contact chemosensory afferents in an insect brain. J Comp Neurol 493:291–308
Poitout S, Bues R (1974) Linoleic acid requirement of Lepidoptera reared in artificial medium, Noctuidae, Quadrifinae and Plusiinae: Chrysodeixis chalcites Esp., Autographa gamma L., Macdunnoughia confusa Stph., Trichoplusia ni Hbn. Ann Nutr Aliment 28:173–187
Powell G, Maniar SP, Pickett JA, Hardie J (1999) Aphid responses to non-host epicuticular lipids. Entomol Exp Appl 91:115–123
Rees CJC (1970) The primary process of reception in the type 3 (‘water’) receptor cell of the fly, Phormia terranovae. Proc R Soc Lond B 174:469–490
Renwick JA (1989) Chemical ecology of oviposition in phytophagous insects. Experientia 45:223–228
Rogers SM, Simpson SJ (1999) Chemo-discriminatory neurones in the sub-oesophageal ganglion of Locusta migratoria. Entomol Exp Appl 91:19–28
Rospars JP (1983) Invariance and sex-specific variations of the glomerular organization in the antennal lobes of a moth, Mamestra brassicae, and a butterfly, Pieris brassicae. J Comp Neurol 220:80–96
Rospars JP (1988) Structure and development of the insect antennodeutocerebral system. Int J Insect Morphol Embryol 17:243–294
Schoonhoven LM, van Loon JJA (2002) An inventory of taste in caterpillars: each species its own key. Acta Zool Acad Sci Hung 48:215–263
Singh RN (1997) Neurobiology of the gustatory systems of Drosophila and some terrestrial insects. Microsc Res Tech 39:547–563
Skiri HT, Ro H, Berg BG, Mustaparta H (2005) Consistent organization of glomeruli in the antennal lobes of related species of heliothine moths. J Comp Neurol 491:367–380
Steinbauer MJ, Schiestl FP, Davies NW (2004) Monoterpenes and epicuticular waxes help female autumn gum moth differentiate between waxy and glossy Eucalyptus and leaves of different ages. J Chem Ecol 30:1117–1142
Stocker RF (1994) The organization of the chemosensory system in Drosophila melanogaster: a review. Cell Tissue Res 275:3–26
Thorne N, Chromey C, Bray S, Amrein H (2004) Taste perception and coding in Drosophila. Curr Biol 14:1065–1079
Udayagiri S, Mason CE (1997) Epicuticular wax chemicals in Zea mays influence oviposition in Ostrinia nubilalis. J Chem Ecol 23:1675–1687
Vosshall LB, Wong AM, Axel R (2000) An olfactory sensory map in the fly brain. Cell 102:147–159
Wang Z, Singhvi A, Kong P, Scott K (2004) Taste representations in the Drosophila brain. Cell 117:981–991
Wieczorek H, Köppl R (1978) Effect of sugars on the labellar water receptor of the fly. J Comp Physiol A 126:131–136
Acknowledgments
This work was supported by a doctoral fellowship to AP from the doctoral school ABIES and the bilateral exchange program Aurora (EGIDE). Romina Barrozo and Sebastian Minoli are acknowledged for help with the figures and Dominique Van Oort for help with the scanning electron microscopy. We thank Delphine Calas-List for helpful comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
F. Marion-Poll and S. Anton share senior authorship.
Rights and permissions
About this article
Cite this article
Popescu, A., Couton, L., Almaas, TJ. et al. Function and central projections of gustatory receptor neurons on the antenna of the noctuid moth Spodoptera littoralis . J Comp Physiol A 199, 403–416 (2013). https://doi.org/10.1007/s00359-013-0803-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00359-013-0803-0