
Abstract
The Australian desert ant Melophorus bagoti often follows stereotypical routes through a cluttered landscape containing both distant panoramic views and obstacles (plants) to navigate around. We created an artificial obstacle course for the ants between a feeder and their nest. Landmarks comprised natural objects in the landscape such as logs, branches, and tussocks. Many ants travelled stereotypical routes home through the obstacle course in training, threading repeatedly the same gaps in the landmarks. Manipulations altering the relations between the landmarks and the surrounding panorama, however, affected the routes in two major ways. Both interchanging the positions of landmarks (transpositions) and displacing the entire landmark set along with the starting position of the ants (translations) (1) reduced the stereotypicality of the route, and (2) increased turns and meanders during travel. The ants might have used the entire panorama in view-based travel, or the distal panorama might prime the identification and use of landmarks en route. Despite the large data set, both options (not mutually exclusive) remain viable.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Many animals use terrestrial objects, landmarks, for navigation (Shettleworth 2010). Landmarks may be encoded and represented in a map-like fashion, although this idea of ‘cognitive mapping’ is fraught with controversy in both vertebrate (Benhamou 1996; Bennett 1996) and invertebrate animals (Gould 1986; Wehner and Menzel 1990; Dyer 1991; Menzel et al. 2005; review, Shettleworth 2010). Just as often, landmarks are used to chart routes, which are typically stereotypical paths through a landscape dotted with objects. Route-following in vertebrate animals has been little studied (but see Calhoun 1963), but in insect navigation, the topic has received much attention (Rosengren 1971; Collett et al. 1992, 1993; Zhang et al. 1996; Kohler and Wehner 2005). We here begin to examine the mechanisms of route-following in a species known for the behaviour, the Australian red honey ant Melophorus bagoti.
M. bagoti lives in a wide range over semi-arid Central Australia, where the habitat is typically filled with plants, from trees and bushes to grass tussocks (Muser et al. 2005), where mated queens attempt to dig nests in the ground to start colonies (Schultheiss et al. 2010). The ant is the most thermophilic on the continent (Christian and Morton 1992), and forages in the heat of the day in the few hot months of summer. The heat of the ground surface makes it too volatile for chemical trails, so that the ants forage individually, relying on both path integration (Wehner et al. 2006; Narendra 2007; Narendra et al. 2007) and, more commonly, landmarks, through which they run stereotypical routes (Kohler and Wehner 2005; review, Cheng et al. 2009).
While M. bagoti’s stereotypical routes through natural terrain have been plotted (Kohler and Wehner 2005), many details of its route-following behaviour have yet to be characterised. We investigated the interaction between the use of individual landmarks along a route and the broad context of the panoramic view around the ant as she travels. The ants had to travel an artificial obstacle course between a feeder and their nest. Using natural materials found in the landscape to create landmarks along the route, objects such as logs, tussocks, and branches, we manipulated these movable but stationary landmarks on tests. After sufficient training, we effected various transformations, including the removal of the landmarks, transpositions of landmarks, in which two or more landmarks switched positions, and displacements, in which the whole array of landmarks was translated and/or rotated.
Based on much past research on the importance of contextual and panoramic cues, we expected that a mismatch between the usual route landmarks and the panoramic context would affect the route-following behaviour: the more the mismatch, the less the usual route would be followed. Panoramic cues might prime the retrieval of memories of landmarks and how to negotiate them (Collett et al. 2003), or they might also be used directly for guidance (Graham and Cheng 2009a). We also expected that a mismatch between the positions of the route objects and the panoramic cues would affect path characteristics, such as how much the ants meandered during travel, and whether and where the ants spent the most time searching, as they often did.
Materials and methods
Animals
Desert ants Melophorus bagoti, the red honey ant, from two nests were tested in situ near their nest from December 2008 through February 2009. Experimentation was carried out in the mornings until ~12:00 and afternoons from 14:30 onwards. The midday period was not conducive to experimentation as the ants often spend large amounts of time taking refuge on a plant (Christian and Morton 1992, personal observation).
Field site
The experiments took place on the grounds of the CSIRO Centre for Arid Zone Research, ~10 km south of Alice Springs, in a setting filled with trees (Acacia and Hakea), bushes, grass tussocks (mostly the invasive buffel grass), and buildings. Views associated with the two nests can be seen in Fig. 1.

Experimental settings and setups. a, b Photos of experimental settings and artificial landmarks used in Experiment 1 (a) and Experiment 2 (b), looking from the feeder towards the nest. c The transposition effected in Experiment 1, with the grey line showing an example of a training run. The same transposition was effected for all ants. d An example of a transposition used in Experiment 2, and a training run. The stars indicate nest location in c and d
Experimental setup
Two experiments with different setups were conducted on two different nests. Each nest was provided with free access to a feeder (a square plastic tub, 20 cm × 20 cm × 15 cm deep, sunk into the ground) 10 m North (Experiment 1) or West (Experiment 2) from their nest. During training, small pieces of cookies were scattered in the feeder for the ants to pick up, and sticks placed in the feeder allowed the ants to climb out, the walls of the plastic tub being extremely difficult for ants to climb. Between the nest and the feeder, we set up obstacles using natural materials, grass tussocks, leaves, branches, logs, and rocks, for the ants to navigate through. A grid of 1-m square units consisting of string wound around tent pegs stuck into the ground provided reference for recording an ant’s trajectory on gridded paper. The strings were off the ground and did not interfere with ant movements. Such a grid was also set up for tests on a distant test field.
Experiment 1 was set up with an ‘obstacle course’ of six landmarks placed directly on the ground (Fig. 1a). The landmark nearest the feeder was 2.5 m away, with each successive landmark 1 m farther; the farthest landmark thus lay 7.5 m from the feeder. Throughout training, the landmarks remained stationary in the same configuration. Experiment 2 was set up with four rows of landmarks between feeder and nest, the nearest row to the feeder 3.5 m away and the farthest row 7.5 m away (Fig. 1b). The rows were evenly spaced, with three objects in row 1, two in row 2, three in row 3, and two in row 4. For easy cue manipulations such as displacements, we placed all the objects in white half cylinders (80 cm long, 20 cm wide) sunk into the ground. Between the cylinders lay a gap of 40 cm. Throughout training, these landmarks also remained stationary in the same configuration. At the start of the journey home, we created a short stretch of alley to start the ants off in the correct general direction and prevent them from scattering in random directions. The alley was a gap 120 cm long and 40 cm wide with 10-cm-high white plastic walls on the sides formed with segments of ‘channels’. It was aligned in the feeder-nest direction except for one test in which the orientation of the landmark array was rotated (Distal Rotated, described below). On that test, the starting alley was also rotated to face the landmarks. The ants were forced to start their homeward journeys down this alley during training and on tests.
Procedure
On her first arrival at the feeder, each ant was painted for individual identification. Painted ants were free to travel back and forth between feeder and nest. After at least one full day of training, a painted ant might have her training return journey recorded and then tested. On these occasions, the ant was allowed to travel home with a piece of cookie (as during training). We recorded only the runs of ants that carried a piece of cookie the entire journey. Her path was recorded on a gridded piece of paper. We trapped the ant and picked her up when she reached the vicinity of the nest and began to make searching loops. Such an ant has run off her homebound vector based on path integration, so that the vector to run is now the zero vector. She is called a zero-vector ant. Zero-vector ants are used to test how ants use landmarks for navigation because they cannot derive a direction of travel based on path integration, and this because the zero vector does not specify a direction. We then brought the zero-vector ant back to the rim of the feeder and placed her on the ground to run home again. Her path was recorded again, and this time she was allowed to enter her nest. When the ant returned again to the feeder, she was tested again under zero-vector conditions. Typically, this was a matter of minutes, but the timing varied as we had typically other trained ants to test as well. If the three runs, one full-vector and two zero-vector runs, showed the same pattern in negotiating the obstacles, the ant would then be tested in a manipulated condition the next time she appeared at the feeder. The same pattern meant that an ant went between the obstacles in the same manner; technically, she crossed the same line segments connecting the landmarks in the same order. All tests were on zero-vector ants, each preceded by a normal full-vector homebound training run, which was recorded. The ant was captured in the vicinity of the nest, and then allowed to run home again under zero-vector conditions, this time with the manipulations effected on the landmarks. During manipulated tests, the sticks in the feeder were removed to prevent any ants in the feeder from exiting, this to prevent ants from being trained with a different setup.
Manipulated tests
In Experiment 1 (Fig. 1a), the only manipulation consisted of transposing the landmarks while keeping one landmark in each of the landmark positions used in training (Fig. 1c). In Experiment 2, we effected the transposition on all rows, within each row. For rows 2 and 4, with only 2 landmarks, only one transposition is possible. Rows 1 and 3 have 3 landmarks each, and two transpositions are possible in which the positions of all three landmarks are switched. We picked the transposition for each ant that preserved the pair of landmarks between which the ant usually travelled. For example, suppose that the landmarks are aligned A, B, C, from left to right, and the ant habitually travelled between B and C (Fig. 1d). The transposition would change the arrangement to B, C, A, allowing the continued possibility of travelling between B and C. This same transposition was effected if the ant habitually travelled to the right of C.
A second set of transformations consisting of landmark translations was conducted in Experiment 2 only. For these transformations, the entire array of landmarks was rigidly translated to the right, looking from the feeder to the nest. The tested ant’s starting point was likewise translated. Geographically, it was only possible to translate to the right, where open space was found. In a translation, the direction of the array was preserved; geometrically, no rotation was effected. We effected translations in increments measured in landmark units, with a 1-unit move placing one landmark to its neighbour’s position 1.2 m to the right. The four translations effected were: Translation 1 (1.2 m to the right), Translation 3 (3.6 m to the right), Translation 8 (9.6 m to the right), and Translation Distal (to a distant, completely unfamiliar test field ~100 m away).
We tested ants in a number of other conditions in Experiment 2, some to gather data on other manipulations, others as controls. In the Panorama test, ants were allowed to return home from the feeder on the training field, but all the experimental landmarks were removed, leaving solely the distant panoramic cues for orientation. The Distal Rotated test probed the significance of the compass direction of the landmarks on a distal test field. Thus, the test took place on the distal test field as in the Translation Distal test, but the landmark array was rotated by 80° clockwise, as was the starting alley. On the Distal No Landmark test, the ants were again tested on the distal unfamiliar test field, but without any of the experimental landmarks present. Finally, a control group was tested in the Translation Distal condition (called Distal Control). The Distal Control group was trained to home from the same feeder without any landmarks. They were tested on the distal test field with the landmarks in place that their experimental counterparts had been trained with. This condition tested whether the ants had any untrained tendency to approach the landmarks as beacons, as ants sometimes do (Graham et al. 2003).
Data analysis
We present the gist of data analysis here, leaving details for the “Results” section to minimise repetitiveness. Paths were digitised using GraphClick™ software into coordinates with (0, 0) being the start of the journey, the x-axis representing left–right travel (negative to the left), and the y-axis representing homeward travel in the positive direction. For Experiment 1, we noted whether the sides chosen corresponded with the same side (left or right) of a landmark or with the Earth-based position (to the same side irrespective of which landmark was at the position); depending on the ant’s habitual path, these predictions sometimes coincided. For Experiment 2, we also noted whether an ant on a transposition test followed the landmarks (going through the gap defined by the same pair of landmarks irrespective of their position) or the Earth-based position (going through the same Earth-based gap irrespective of the landmarks on either side), or travelled another way (all others). On other tests in Experiment 2, we also classified travel through each row exhaustively, as described in “Results”.
Indirectness and curvature of paths
When the visual cues were transformed, ants often hesitated, turned left and right as they travelled, and sometimes appeared to scan the visual surround (a behaviour that we are analysing in another work). We calculated a measure of such ‘wiggling’, called Meander, and compared it across all test conditions, including training runs.
Effect of nest entrance relocation
A natural, unplanned event allowed us to analyse the effect of nest location on training paths. During the course of experimentation, the nest entrance relocated from a point slightly to the right of the y-axis to a location slightly to the left. We compared the training runs of ants trained before and after this transition (details in “Results”).
Unless otherwise stated, results of statistical tests were considered significant at alpha = 0.05.
Results
Our observations showed that the ants readily solved the obstacle course between the feeder and the nest during training. Many ants established stereotypical routes through the same gaps spontaneously. Experiment 2 had larger obstacles, making a larger deviation for ants to go through a non-habitual gap. A higher proportion of ants made three consecutive runs through the same gaps in Experiment 2 (192 of 210 or 91.4%) than in Experiment 1 (66 of 112 or 58.9%; p < 0.001, Fisher’s exact test).
Transposition tests
Transposing landmarks clearly affected the stereotypical routes that the ants took to head home, in both Experiment 1 and Experiment 2 (Fig. 2). At the time of testing, all ants (100%) had traversed the same pattern of gaps back to their nest for three consecutive homing journeys. On the transposition test, only 56 of 72 choices (77.8%) in Experiment 1 and 33 of 72 choices (45.8%) in Experiment 2 followed the habitual gap in Earth-based coordinates (Earth-based or Both in Fig. 2c, d), a higher proportion in Experiment 1 (p < 0.001, Fisher’s exact test).

Routes in the transposition test. a, b Paths from training runs (top) and from transposition tests (bottom) in Experiment 1 (a, n = 12) and Experiment 2 (b, n = 18). The training runs are from the runs preceding the transposition test. The rectangles in b demarcate the starting alley. c Choice of side at each row (with a single landmark) on transposition tests in Experiment 1. An Earth-based choice means choosing the usual Earth-based side at that row, irrespective of what landmark was at that row. A landmark choice means choosing the side of the displaced landmark that an ant habitually took in training. Sometimes both these criteria were satisfied. d Choice of gaps at each of the 4 rows (see panel b) on transposition tests in Experiment 2. An Earth-based choice means choosing the usual Earth-based gap irrespective of what landmarks make up the gap. A landmark choice means choosing the gap through the displaced landmarks
In Experiment 1, the ants were more likely to follow the Earth-based route, striking the habitual route through what were now ‘wrong’ landmarks, than they were to steer around the side of a transposed landmark that they habitually headed for in training (Fig. 2a, c). In fact, the ratio was 3:1. Ignoring choices categorised as Both or Other, 10 ants had at least 1 Earth-based or Landmark choice. Averaging across individual ants, the mean proportion of Earth-based choices [Earth-based/(Earth-based + Landmark)] was 0.75. The 95% confidence interval exceeded 0.5. In Experiment 2, ants followed the Earth-based routes (flanked by ‘wrong’ landmarks) more in rows 1, 2, and 4, and the landmarks more in row 3 (Fig. 2d). Considering only Earth-based and Landmark choices (and ignoring 9 Other choices), the average proportion of Earth-based choices across individual ants was 0.57, not significantly different from 0.5.
Panorama test
On the Panorama test, the ants travelled home successfully from the feeder on the training field with the training landmarks removed (Fig. 3). Only one of 18 ants failed the test, not reaching rows 3 and 4 (Never Reached choices). Even without the habitual landmark obstacles, the ants mostly went through the habitual landmarkless ‘gap’ at row 1 (Earth-based choices, Fig. 3a). Thereafter, this tendency diminished somewhat, with more ants travelling past the row somewhere else (Else choices).

a Gap choices on the Panorama test in Experiment 2, in which the usual landmarks along the route had been removed (n = 18). The now non-existent ‘gaps’ were defined by Earth-based positions (Earth-based choice). Crossing the row anywhere other than the Earth-based gap was called “Else”. b Runs on the Panorama test, with the dotted lines indicating the usual positions of the landmarks missing on the test. The star indicates the nest position, and the rectangles demarcate the starting alley
Displacement tests
On translation tests in Experiment 2, the entire array of landmarks was shifted to the right without rotation, with the ant’s starting point likewise shifted. For each row in each translation, we defined four categories of travel. The ant could travel through its habitual gap, taking the Usual Route. She could choose another gap (Wrong Gap). Or she could use the Panorama. This was defined as a combination of travelling through a gap to the left of the usual one, on the row in question and all subsequent rows, and eventually heading towards the nest. Determining whether an ant headed to the nest proved completely unambiguous. On exiting the landmark array, an ant either headed towards the nest and entered it or else she searched around in loops. Finally, she might never reach a row. The unsurprising strong influence of the usual panorama encountered on training runs is indicated by marked differences in the nature of routes as a function of the distance of translation (Fig. 4). The proportion of ants going through the habitual gap (Usual route choices) diminished with distance of translation. The Panorama category increased at distances of 3.6 m (Translation 3) and 9.6 m (Translation 8) translation, but at 9.6 m, the number of ants that never reached the last row (and never found the nest) increased. Of course, at the distal test field at which no familiar panoramic cues were found, the use of any familiar panoramic cues was impossible. A statistical comparison can be made by considering only row 1 choices, such that the data are independent, with one choice from each ant across all test conditions. Excluding the Never Reached category (just one ant in the Translation Distal test), the proportions of categories of choices differed across translation conditions (χ 2 (6) = 29.2, p < 0.001).

Gap choices and path trajectories on tests with landmark displacements in Experiment 2. The ants’ starting position was displaced along with the landmarks. a Row-by-row gap choices. The usual route indicates an ant going through the same gap defined by the displaced landmarks that she took in training. A Panorama choice was defined as a choice to the left of the usual gap, followed by the ant eventually heading home. b–e Paths on translation tests, from Translation 1 (b, n = 18), Translation 3 (c, n = 20), Translation 8 (d, n = 16), and Translation Distal (e, n = 11). Solid black lines indicate paths defined as following the panoramic cues. (Paths of the Distal Rotated test appear in Fig. 5.) The star indicates the nest position, and the rectangles demarcate the starting alley. f Summary of gap choices in different displacement tests averaging all rows
Navigational behaviour changed row by row as well (Fig. 4a). The choice of the usual route diminished row by row. Except for one tie in Translation Distal, the monotonically decreasing pattern held for all translation tests. The exact probability of a monotonically decreasing sequence of 4 is 1/(4!) = 0.042. The exact probability for 3 such sequences (in Translation 1, Translation 3, and Translation 8) is 0.0423 = 7.2−5.
We found the opposite trend with the Panorama choice: this increased row by row, monotonically with a tie in each of Translation 1, Translation 3, and Translation 8. (The Translation Distal and Distal Rotated tests produced no Panorama choices for obvious reasons.) We refrain from conducting inferential statistics here because the pattern is dictated by the definition of the Panorama choice. If the choice of one row is Panorama, the choice on all subsequent rows must be Panorama as well. More informative is the row at which the ant first chose the Panorama option. Combining only Translation 3 and Translation 8, in which most of the Panorama choices were made, Panorama was chosen first 10 times in row 1, 8 times in row 3, and once each in row 2 and row 4.
A higher proportion of ants never reached a row before turning around in search loops as translation distance increased, with the Never Reached category comprising a majority of rows 3 and 4 on the distal test field when the landmarks were not rotated (Translation Distal), and comprising 100% of the runs when the landmarks were rotated by 80° (Distal Rotated). In the Translation Distal test, the vast majority of ants reached the first row, but less than 20% of the ants chose the usual gap in row 1, and a majority of ants never reached row 3 (Fig. 4a). In the Distal Rotated test, only a minority reached the first row. Most ants searched in loops (Figs. 4a, 5b), as did ants in the Distal No Landmark test (Fig. 5c) and the Distal Control test (Fig. 5d). The landmarks on the Distal Control test, never encountered before by the ants, did not attract the ants to them.

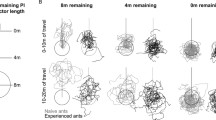
Meander in all tests in Experiment 2 and paths of the three remaining conditions. a Meander in all conditions, computed from the orientations of 30-cm line segments calculated from the digitised (x, y) coordinates of the path. Each successive segment was formed by drawing a circle of 30 cm radius around the end of the previous segment and noting where the path crossed this circle. The circle was placed at the starting point for segment 1. Meander was defined as the average absolute angular difference between successive segments in radians. Conditions sharing the same letter are not significantly different by Tukey’s post hoc tests. Individual points in the training condition cannot be readily identified because there were 499 data points. LM stands for landmark. b–d Paths in the Distal Rotated (b, n = 15), Distal No Landmark (c, n = 29) and Distal Control (d, n = 15) conditions. In c, the dotted lines represent the positions of the landmark obstacles encountered during training, now absent on the test. The star indicates the nest position, and the rectangles demarcate the starting alley
Meander of paths
On all the tests in Experiment 2, we devised a measure of Meander by dividing the path into 30-cm line segments. A circle of 30 cm radius was placed at the start of a route and a straight line segment drawn to where the route crossed this circle to deliver segment 1. The circle was then centred at the end of segment 1, and where the route crossed the circle delivered the end of segment 2, etc. The Meander index measures how much the path changes direction from segment to segment. The absolute angular deviation in radians from one segment to the next was averaged over all segments. Thus, 0 radians meant that the two segments were collinear, while π radians meant that the ant turned straight back.
Variances of Meander differed significantly across conditions by O’Brien’s test (O’Brien 1979) (Fig. 5a, p < 0.001). We then used Tukey’s post hoc test to compare all pairs of conditions (Fig. 5a). The training condition produced the least Meander. Even a translation of 1.2 m (Translation 1) produced more Meander than training runs. Transposition produced more Meander than Translation 1. The Panorama test, in which the landmarks on the training field were removed for the test, also produced more Meander than training runs. This shows that the removal of the training landmarks made the ants turn back and forth more. Nevertheless, the powerful role of the distant panorama is revealed by the fact that paths on this test had less Meander than tests on the distal test field (conditions 7–10 in Fig. 5a). Finally, rotation of the landmarks on the distal test field did not produce any noticeable effects on Meander (compare conditions 7 and 8).
Distribution in feeder-nest direction
Another informative path characteristic is how much of each path is distributed across the length of the journey. Because in some conditions, ants initiated searching loops from the start, we extended the range along the feeder-nest direction to 4 m behind the feeder (−4 to 10 m). The training condition, the Panorama test, and tests on the distal test field were analysed. For each ant, we calculated the median point, spatially, of the path, and the interquartile spread. Tukey’s post hoc comparisons were then used to compare the conditions pairwise (Table 1). Runs from training and Panorama tests (Figs. 2b, 3b, respectively) had medians near the halfway point (5 m) and wide spreads, reflecting straight runs. Groups with no relevant landmark information of any kind (Distal No Landmark, Fig. 5c; and Distal Control, Fig. 5d), either because no landmarks were provided (Distal No Landmark) or because they were not trained with landmarks (Distal Control), showed peaks near the starting point (0 m) with lower interquartile ranges than training runs, reflecting searching centred on the starting point. Thus, ants not trained with landmarks were not attracted to them. Ants that were trained with landmarks headed towards the training landmarks on the distal test to some extent (Translation Distal and Distal Rotated groups, Figs. 4e and 5b, respectively). On this measure, the directional orientation of the landmarks seemed to matter: when the landmarks were rotated by 80°, the manipulation induced the ants (in the Distal Rotated group) to exhibit a median at a shorter distance from the start.
Effect of nest location on paths
Our final analysis was based on a natural manipulation that the ants themselves effected. During the course of experimentation, the ants shifted their nest entrance, from very slightly to the right of our y-axis to slightly to the left of the y-axis. Considering ants with at least three consecutive training runs through the same gaps, the gap choices differed as a function of nest location (Fig. 6), biased more to the right when the nest was to the right. Considering each row separately, and only choices in which at least one group had ≥5 ants making that choice, χ 2 tests of independence showed that the distributions differed significantly between the two groups at each row (ps < 0.005). This pattern shows that the ants tended to weave an obstacle course around the vector direction that pointed to their nest.

The choice of gaps on training runs before and after the nest entrance moved from very slightly to the right of the y-axis (left panel) to slightly to the left of the y-axis (right panel) in the course of experimentation. The thickness of arrows represents the proportion of choices through that gap. Dashed lines represent the straight line connecting the feeder and the nest, represented by the black dots
Discussion
The results show in some detail the important link between the distant panorama and route-following behaviour through an obstacle course. What is still not clear from the results is whether the distant panorama functions more as a navigational cue directly guiding route-following, or more as contextual cues triggering appropriate behaviour with respect to the individual landmarks forming the obstacle course. We focus on these two major points in this section.
Link between the distant panorama and route-following
In training, many ants developed stereotypical courses through the obstacles between feeder and nest. All the ants that participated in tests had followed a habitual route through the same gaps a number of times before being tested. Changing the visual scene in any way changed the habitual behaviour, whether landmarks exchanged positions, shifted locations, or were absent altogether. The ants were then less likely to take a route through the habitual gaps defined by the positions of the landmarks. And they turned back and forth (meandered) more in their travel. In the case of shifting the entire set of landmarks along with the starting position of the ants (translations), a ‘dose-dependent response curve’ was evident: the larger the translation, the bigger the effect (Fig. 4 for gap choices, Fig. 5 for Meander).
These ‘dose response’ characteristics might indicate probabilistic contextual modulation in individual ants, meaning that the probability that the context will trigger the usual path decreases gradually with increasing change of context. But contextual modulation might also be all-or-none, with each ant having a sharp threshold of tolerance for change of context, beyond which the context simply does not trigger the usual behaviour and ‘all bets are off’. While the group data show a ‘dose response curve’ and not a step function, we tested each ant only once. It is possible to obtain a dose-response curve for a group if individual ants vary in step-like tolerance thresholds. As an analogy, individual animals might learn in an all-or-none basis as a function of trials of training while the group data show a smooth learning acquisition curve (Gallistel et al. 2004).
On the transposition test, more ants charted the habitual route in Earth-based coordinates in Experiment 1 than in Experiment 2. Several reasons might account for this difference. In Experiment 1, the ants encountered one landmark at a time as they traversed the y-axis towards the nest, whereas in Experiment 2, multiple landmarks needed to be negotiated at each row. The obstacle avoidance requirements were thus simpler in Experiment 1. Transpositions in Experiment 1 also meant that the same gap would often be chosen on the basis of either the landmark or the Earth-based route. In addition, Experiment 2 contained larger landmarks dominating more of the view on the homeward route.
Some weak evidence from this study suggests that the ants might have also encoded the compass direction to travel to the first gap, presumably based on a sky compass. This is called a local vector (Collett et al. 1998; Collett and Collett 2002; Cheng 2006). On the Panorama test in Experiment 2, with the training landmarks missing, many ants still headed to the usual landmarkless ‘gap’ at the first row. Also in Experiment 2, more ants in the Translation 1 test followed the usual landmarks than ants in the Transposition test, and they also exhibited less Meander. The transposition switched landmark positions and required the ants to travel in a different compass direction to the usual landmarks, creating a possible conflict between landmarks and the local vector. On translation tests, the starting position was also translated, preserving the compass direction and local vector to the usual gap. Some ants reached the usual gap at the first row of landmarks under diverse conditions, even on the distal test field (Translation Distal test, Fig. 4a). But no ants reached the usual gap at the first row when the array of landmarks was rotated on the distal test field (Distal rotated test, Fig. 4a), and few reached the first row at all, despite the fact that the starting alley pointed towards the landmarks. A local vector on emerging from the starting alley might have oriented the ants towards the ‘correct’ landmarks when they were in the direction found during training. Definitive evidence for the use of local vectors was found recently by testing the ants in a round arena devoid of skyline cues (Legge et al. 2010). Even with conflicting landmark cues, the ants showed a strong tendency to head in the trained compass direction to find an exit out of the arena. Other ants too can learn local vectors and motor sequences (Collett et al. 1998; Macquart et al. 2006, 2008).
Panorama and/or contextual cues
Our results confirm once again that M. bagoti follows stereotypical routes, in our case through an artificially constructed obstacle course. A common view of this route-following behaviour is that what to do with respect to a landmark object is conditioned upon or triggered by contextual cues (Collett et al. 1998; Collett and Collett 2002; Cheng 2006). The panoramic cues function as contextual cues to facilitate the use of particular landmarks, helping the animal to identify landmarks, providing signposts for behaviour (Collett and Collett 2002), or setting the occasion for the use of servomechanisms based on particular landmarks (Cheng 2006). But contextual cues do not control behaviour directly. That control is placed in the landmark object around which the insect is navigating. Plenty of evidence links contextual cues of all kinds to memory retrieval and behaviour (review, Collett et al. 2003). The physical setting in which the animal is navigating can serve as a contextual cue (Collett and Kelber 1988; Colborn et al. 1999; Cheng 2005), in one case even after the animal has entered a test apparatus that blocked the view of the surroundings (Collett et al. 1997). Other contextual cues include the motivation to travel (for example, having food to take home vs. going out to seek forage; Dyer et al. 2002; Beugnon et al. 2005), the time of the day (Koltermann 1971), the encounter of a particular familiar landmark (Collett et al. 2002), and possibly the distance the insect has already travelled (Srinivasan et al. 1999) and sequential cues (Chameron et al. 1998; Zhang et al. 1999), in which a step in the sequence provides the context for appropriate memory retrieval.
Recent evidence shows that M. bagoti sometimes uses a panoramic snapshot directly for orientation (Graham and Cheng 2009a, b; for suggestive results on honeybees, see Towne and Moscrip 2008). Replacing the natural skyline (a record of how elevated the tops of terrestrial objects are, without identifying the objects) with an artificial skyline made of black cloth forming an arena, is sufficient for the ants to chart an initial direction home (Graham and Cheng 2009a). This suggests that the context/landmark separation may not be necessary for explaining all route-following behaviour, although some contextual cues would still play a role in guiding navigation (for example, providing the motivation to home). Individual landmarks need not be identified at all (see also Macquart et al. 2006). Instead, the entire panorama, encompassing near and far landmark cues, drives behaviour. Matching of global panoramic views can explain both route-following and homing from new release points (Zeil et al. 2003; Wystrach 2009; Wystrach and Beugnon 2009). Might a route actually consist of a series of matches to panoramic skylines, without segregation of distant contextual cues and nearby signposts?
Despite the sizeable set of manipulations, the data presented here can be accommodated equally well on either view. A dose-dependent degradation in following the usual route through the obstacle course after translations of the landmark array (and the ant’s starting position) may reflect an increasing probability of failure of the degraded context to trigger the usual route. Or else it may reflect a degradation in the overall match with the panorama, a panorama including both the landmarks and the distant trees and bushes. The transposition likewise changes both the overall panorama and the link between the panoramic context and individual landmarks. The success of the ants on the Panorama test, with all experimental landmarks absent, should not be seen as a triumph for the hypothesis of the direct control by the panorama. It is possible that beyond row 4, the ants had identified and used other landmarks for homing.
To take two specific outcomes that both major hypotheses support, consider the transposition and Translation 1. With a transposition, the panorama appears to match the training conditions better on the landmark-based route (Fig. 7c compared with Fig. 7a) rather than the Earth-based route (Fig. 7b compared with Fig. 7a). But while Fig. 7c looks to match the training view better, the advantage of following this landmark-based route may well be balanced by the tendency of the ants to head to row 1 in a habitual compass direction. For Translation 1, the match in heading towards the usual route is much better than any other (data not shown), the distant panoramic cues having little changed with the 1.2 m translation. Both hypotheses would predict a high degree of adherence to the usual route, which is what the ants did (Fig. 4).

Panoramic views taken between row 1 and row 2 in the training condition (a) and in the transposition condition (b, c) in Experiment 2. a Panoramic view on a training run. The photo was taken with a curved mirror (GoPano Plus) attached to the front of the lens of a digital camera placed lens down on the ground, and then unwarped using software (PhotoWarp 2.5) to form a cylindrical view, in which the right edge is coincident with the left edge. The camera location was along the usual training route of one ant, between row 1 and row 2. The photo has been blurred to simulate approximately a 5° visual resolution. b A panoramic photo from a transposition test, taken at the same location as in a. The photo was obtained and treated in the same fashion. c A panoramic photo from a transposition test, taken at a location 1.2 m to the left of the camera location in a and b, the location of the best matching gap based on the landmarks along the route. Otherwise, the photo was obtained and treated in the same fashion
We do not believe that the two broad hypotheses are indistinguishable. Nor do we think that they are equivalent models formulated in different words. More detailed modelling of a range of experimentally transformed conditions might well provide discriminating evidence. It is likely that both models are correct, but in different circumstances. Characterising these different circumstances forms an important research agenda.
References
Benhamou S (1996) No evidence for cognitive mapping in rats. Anim Behav 52:201–212
Bennett ATD (1996) Do animals have cognitive maps? J Exp Biol 199:219–224
Beugnon G, Lachaud J-P, Chagné P (2005) Use of long-term stored vector information in the neotropical ant Gigantiops destructor. J Insect Behav 18:415–432
Calhoun JB (1963) The ecology and sociology of the Norway rat. US Department of Health, Education, and Welfare, Bethesda, MD
Chameron S, Schatz B, Pastergue-Ruiz I, Beugnon G, Collett TS (1998) The learning of a sequence of visual patterns by the ant Cataglyphis cursor. Proc R Soc Lond B 265:2309–2313
Cheng K (2005) Context cues eliminate retroactive interference effects in honeybees (Apis mellifera). J Exp Biol 208:1019–1024
Cheng K (2006) Arthropod navigation: ants, bees, crabs, spiders finding their way. In: Wasserman EA, Zentall TR (eds) Comparative cognition: experimental explorations of animal intelligence. Oxford University Press, Oxford, pp 189–209
Cheng K, Narendra A, Sommer S, Wehner R (2009) Traveling in clutter: navigation in the Central Australian desert ant Melophorus bagoti. Behav Proc 80:261–268
Christian KA, Morton SR (1992) Extreme thermophilia in a Central Australian ant, Melophorus bagoti. Physiol Zool 65:885–905
Colborn M, Ahmad-Annuar A, Fauria K, Collett TS (1999) Contextual modulation of visuomotor associations in bumble-bees (Bombus terrestris). Proc R Soc Lond B 266:2413–2418
Collett TS, Collett M (2002) Memory use in insect visual navigation. Nat Rev Neurosci 3:542–552
Collett TS, Kelber A (1988) The retrieval of visuo-spatial memories by honeybees. J Comp Physiol A 163:145–150
Collett TS, Dillmann E, Giger A, Wehner R (1992) Visual landmarks and route following in desert ants. J Comp Physiol A 170:435–442
Collett TS, Fry S, Wehner R (1993) Sequence learning by honeybees. J Comp Physiol A 172:693–706
Collett TS, Fauria K, Dale K, Baron J (1997) Places and patterns: a study of context learning in honeybees. J Comp Physiol A 181:343–353
Collett M, Collett TS, Bisch S, Wehner R (1998) Local and global vectors in desert ant navigation. Nature 394:269–272
Collett M, Harland D, Collett TS (2002) The use of landmarks and panoramic context in the performance of local vectors by navigating honeybees. J Exp Biol 205:807–814
Collett TS, Fauria K, Dale K (2003) Contextual cues and insect navigation. In: Jeffery KJ (ed) The neurobiology of spatial behaviour. Oxford University Press, Oxford, pp 67–82
Dyer FC (1991) Bees acquire route-based memories but not cognitive maps in a familiar landscape. Anim Behav 41:239–246
Dyer FC, Gill M, Sharbowski J (2002) Motivation and vector navigation in honey bees. Naturwissenschaften 89:262–264
Gallistel CR, Fairhurst S, Balsam P (2004) The learning curve: implications of a quantitative analysis. Proc Natl Acad Sci USA 101:13124–13131
Gould JL (1986) The locale map of honeybees: do insects have cognitive maps? Science 232:861–863
Graham P, Cheng K (2009a) Ants use the panoramic skyline as a visual cue during navigation. Curr Biol 19:R935–R937
Graham P, Cheng K (2009b) Which portion of the natural panorama is used for view based navigation in the Australian desert ant? J Comp Physiol A 195:681–689
Graham P, Fauria K, Collett TS (2003) The influence of beacon-aiming on the routes of wood ants. J Exp Biol 206:535–541
Kohler M, Wehner R (2005) Idiosyncratic route memories in desert ants, Melophorus bagoti: how do they interact with path integration vectors? Neurobiol Learn Mem 83:1–12
Koltermann R (1971) 24-Std-Periodik in der Langzeiterinnerung an Duft- und Farbsignale bei der Honigbiene (Circadian memory rhythm after scent and colour training with honey-bees). Z Vergl Physiol 75:49–68
Legge ELG, Spetch ML, Cheng K (2010) Not using the obvious: desert ants, Melophorus bagoti, learn local vectors but not beacons in an arena. Anim Cogn 13:849–860
Macquart D, Garnier L, Combe M, Beugnon G (2006) Ant navigation en route to the goal: signature routes facilitate way-finding of Gigantiops destructor. J Comp Physiol A 192:221–234
Macquart D, Latil G, Beugnon G (2008) Sensorimotor sequence learning in the ant Gigantiops destructor. Anim Behav 75:1693–1701
Menzel R, Greggers U, Smith A, Berger S, Brandt R, Brunke S et al (2005) Honeybees navigate according to a map-like spatial memory. Proc Natl Acad Sci USA 102:3040–3045
Muser B, Sommer S, Wolf H, Wehner R (2005) Foraging ecology of the thermophilic Australian desert ant, Melophorus bagoti. Aust J Zool 53:301–311
Narendra A (2007) Homing strategies of the Australian desert ant Melophorus bagoti I. Proportional path integration takes the ant half-way home. J Exp Biol 210:1798–1803
Narendra A, Cheng K, Wehner R (2007) Acquiring, retaining and integrating memories of the outbound distance in the Australian desert ant Melophorus bagoti. J Exp Biol 210:570–577
O’Brien RG (1979) A general ANOVA method for robust tests of additive models of variances. J Am Stat Assoc 74:877–880
Rosengren R (1971) Route fidelity, visual memory and recruitment behaviour in foraging wood ants of the genus Formica (Hymenoptera, Formicidae). Acta Zool Fenn 133:1–105
Schultheiss P, Schwarz S, Wystrach A (2010) Nest relocation and colony founding in the Australian desert ant, Melophorus bagoti Lubbock (Hymenoptera: Formicidae). Psyche. doi:10.1155/2010/435838
Shettleworth SJ (2010) Cognition, evolution, and behavior, second edition. Oxford University Press, New York
Srinivasan MV, Zhang SW, Berry J, Cheng K, Zhu H (1999) Honeybee navigation: linear perception of short distances travelled. J Comp Physiol A 185:239–245
Towne WF, Moscrip H (2008) The connection between landscapes and the solar ephemeris in honeybees. J Exp Biol 211:3729–3736
Wehner R, Menzel R (1990) Do insects have cognitive maps? Ann Rev Neurosci 13:403–414
Wehner R, Boyer M, Loertscher F, Sommer S, Menzi U (2006) Ant navigation: one-way routes rather than maps. Curr Biol 16:75–79
Wystrach A (2009) Ants in rectangular arenas: a support for the global matching theory. Commun Integr Biol 2:388–390
Wystrach A, Beugnon G (2009) Ants learn geometry and features. Curr Biol 19:61–66
Zeil J, Hofmann MI, Chahl JS (2003) Catchment areas of panoramic snapshots in outdoor scenes. J Opt Soc Am 20:450–469
Zhang SW, Bartsch K, Srinivasan MV (1996) Maze learning by honeybees. Neurobiol Learn Mem 66:267–282
Zhang SW, Lehrer M, Srinivasan MV (1999) Honeybee memory: navigation by associative grouping and recall of visual stimuli. Neurobiol Learn Mem 72:180–201
Acknowledgments
The research was supported by grants from the Australian Research Council (DP0770300) and Macquarie University (graduate research funds and scholarships to S.S., P.S., and A.W.). We thank the CSIRO Centre for Arid Zone Research for letting us use their grounds for research, and two anonymous reviewers for helpful comments on the manuscript. The experiments reported in this article were conducted in compliance with the laws of Australia and the Northern Territory. The authors declare no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Wystrach, A., Schwarz, S., Schultheiss, P. et al. Views, landmarks, and routes: how do desert ants negotiate an obstacle course?. J Comp Physiol A 197, 167–179 (2011). https://doi.org/10.1007/s00359-010-0597-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00359-010-0597-2