
Abstract
Among lampyrids, intraspecific sexual communication is facilitated by spectral correspondence between visual sensitivity and bioluminescence emission from the single lantern in the tail. Could a similar strategy be utilized by the elaterids (click beetles), which have one ventral abdominal and two dorsal prothoracic lanterns? Spectral sensitivity [S(λ)] and bioluminescence were investigated in four Brazilian click beetle species Fulgeochlizus bruchii, Pyrearinus termitilluminans, Pyrophorus punctatissimus and P. divergens, representing three genera. In addition, in situ microspectrophotometric absorption spectra were obtained for visual and screening pigments in P. punctatissimus and P. divergens species. In all species, the electroretinographic S(λ) functions showed broad peaks in the green with a shoulder in the near-ultraviolet, suggesting the presence of short- and long-wavelength receptors in the compound eyes. The long-wavelength receptor in Pyrophorus species is mediated by a P540 rhodopsin in conjunction with a species-specific screening pigment. A correspondence was found between green to yellow bioluminescence emissions and its broad S(λ) maximum in each of the four species. It is hypothesized that in elaterids, bioluminescence of the abdominal lantern is an optical signal for intraspecifc sexual communication, while the signals from the prothoracic lanterns serve to warn predators and may also provide illumination in flight.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Over the last two decades, neuroethological investigations of optical signaling in fireflies have demonstrated spectral tuning between visual receptors and bioluminescence emission in a number of species (Lall et al. 1980a, b, 1982; Eguchi et al. 1984). A mathematical optimization model based on bioluminescence signal-to-environmental photon noise ratio was developed to explain the presence of different colors of bioluminescence among North American fireflies (Seliger et al. 1982a, b). The predictions of the model with respect to different combinations of visual pigments and screening pigments resulting in broad or narrow visual spectral sensitivity functions [S(λ)] among night- and twilight-active fireflies, respectively, have now been verified by microspectrophotometry (MSP) (Cronin et al. 2000). Since the bioluminescence signals are spectrally narrow and limited in photon content and appear against a spectrally broad background, the spectral correspondence between the S(λ) functions of the visual receptors (i.e., the detectors for the bioluminescence optical signals) and the emission spectra of the bioluminescence optical signals significantly enhances the detection of species-specific bioluminescence optical signals. These signals are utilized for sexual communication (Seliger et al. 1982a, b) and represent a significant dimension of the evolutionary adaptation in species inhabiting different habitats and photic niches.
The lampyrids (fireflies) possess only one lantern, whereas the bioluminescent elaterids (click beetles) have three: a pair of dorsal lateral prothoracic lanterns and a single ventral abdominal lantern. In various species of elaterids, the dorsal lanterns emit green light (range 525–559 nm), while the ventral lanterns emit lime-green to bright yellow or orange (range 562–584 nm) (Bechara 1988; Colepicolo-Neto et al. 1986). In a recent paper (Lall et al. 2000), the spectral correspondence between the visual sensitivity and bioluminescence emission was described in Pyrophorus punctatissimus.
For the present investigation, four species of click beetles were selected representing three genera of sympatric and disjunct occurrence. P. divergens and P. punctatissimus belong to a group of species of the genus Pyrophorus, which occur in the Amazon and in the Atlantic forests (Costa 1971). These two species are only sympatric in openfields (“cerrados”) of the state of Mato Grosso (MT) where they were collected for this study. Another Pyrophorinid species, Fulgeochlizus bruchii, is known to occur only in Argentina and in the cerrados of central Brazil (Costa 1975, 1991). The genus Pyrearinus includes over 50 species found throughout South America. P. termitilluminans is endemic to the Brazilian cerrados, where their larvae and pupae live in shallow tunnels excavated into termite mounds (Costa 1982). During the rainy season, the larvae expose their heads and prothoracic green lanterns in the exits of the tunnels to attract and catch flying prey, mostly termites and ants. Their habitat is scattered with “luminous termite mounds” (Bechara 1988). All pyrophoronids are vespertine and nocturnal and their nuptial flights last less than 30 min. When male elaterids emit yellow/orange light from their abdominal lanterns, females walking on the leaves and trunks of the trees respond with their abdominal light, and finally they copulate.
There are three aspects to the present study: (1) to obtain in situ MSP absorption spectrum of visual and screening pigments in P. punctatissimus and P. divergens; (2) to determine electroretinographic (ERG) spectral sensitivity of the compound eyes of four elaterid species (F. bruchii, Pyrearinus termitilluminans, Pyrophorus punctatissimus, and P. divergens); (3) to obtain in vivo bioluminescence emission spectra for the abdominal lanterns in both the male and the female of the four species. We report here the spectral correspondence between the bioluminescence emission and vision in each of the four click beetle species (three genera) under investigation. These species were selected by virtue of their easy collection and abundance in the Brazilian cerrados. The hypothesis being tested in these experiments is whether in different species of click beetles inhabiting Brazilian cerrados, there exists spectral correspondence between visual sensitivity and bioluminescence emission for courtship signaling between the sexes.
Materials and methods
Insects: Coleoptera
Adults of Pyrophorus punctatissimus, P. divergens, F. bruchii, and Pyrearinus termitilluminans were collected during the early hours of the night in the Fazenda Santa Cruz (municipality of Costa Rica) located in the state of Mato Grosso do Sul, Central Western Brazil, during the month of October from 1996 to 1998. Pyrophorus divergens was also collected in a remnant of mesophyll tropical forest in the municipality of Cotia, located in the state of São Paulo, southwestern Brazil, about 40 km from São Paulo city in 1997 and 1998.
These specimens were maintained in the laboratory in plastic containers at room temperature. Periodically, the insects were fed sugar water. Species identification was made by Professora C. Costa, Universidade de São Paulo, Museu de Zoologia. Live specimens of both sexes were shipped from São Paulo to Baltimore, MD, USA.
Microspectrophotometry
Procedure
The insects were dark adapted for several days before being prepared for MSP. Dark-adapted animals were decapitated, heads were quick frozen using cryogenic spray, and the frozen heads were mounted on a cryostat (−25 to −30°C) for sectioning at a thickness of 14 μm. Sections were mounted in insect Ringer’s (Muri and Jones 1983) for scanning. Scans were taken as described in Cronin et al. (2000), from 400 to 700 nm at 1 nm intervals, using a circular spot 5 μm in diameter.
The spectrum of each scanned spot, chosen from a location without any pigmentation in the proximal region of the retina (where the inner rhabdomeres are located), was taken in the dark-adapted state and again after a saturating exposure to red light (passed through a Corning CS2-61 filter) to maximally convert rhodopsin to metarhodopsin. The resulting difference spectra for photoconversion were fitted to model rhodopsin/metarhodopsin photopigment pairs, using standard templates (Palacios et al. 1996). Any baseline drift was first removed by subtracting the average spectral value from 651 to 700 nm (where the visual pigments had minimal absorbance). Corrected spectra were fitted from 425 to 650 nm, using all combinations of the model pigments (rhodopsin maximum from 500 to 600 nm; metarhodopsin maximum 450–550 nm, both at 1 nm intervals; extinction ratio, M max/R max, 1.0–2.25, intervals of 0.05). The test spectrum producing the minimum sum of squares of deviations from the data was selected as the best fit.
The absorbance of the yellow screening pigment was measured in retinal sections, placing the beam of the microspectrophotometer in locations within the clear zone where the yellow pigment was visible to the eye. The MSP data were obtained in the Department of Biological Sciences, University of Maryland, Baltimore County.
Electrophysiology
Optical system
A double-beam optical stimulator was constructed in São Paulo, Brazil. The test beam was obtained from a 15 W tungsten microscopic light source; broadband colored filters could be placed in this beam to provide chromatic adaptation light. Each of a set of 18 interference filters (half bandwidth ~10 nm), covering the spectrum from near-UV (360 nm) to red (683 nm), was placed in this beam. A second system was used in Washington, DC, with the test beam originating from a 150 W xenon arc lamp (Quick Illumination Industries, Albany, NY, USA) and passed through a high intensity grating monochromator (600 lines mm−1, Bausch and Lomb, Inc., Rochester, NY, USA), which isolated narrow wavelength bands (half bandwidth of 9.9 nm) from 320 to 680 nm (for details see Lall et al. 1988). A series of glass filters (CS7-54, >70% transmission at 250–390 nm; CS4-97, >40% transmission from 355 to 605 nm; and CS3-72, 40% transmission at >450 nm, Corning Glass Co., Corning, NY, USA) was placed in the light beam to eliminate stray light and the second-order spectrum. The light beam was focused by a series of quartz lenses onto one arm of a bifurcated quartz light guide (3 × 750 mm). A tungsten light source provided the adaptation beam that was focused on the second arm of the bifurcated light guide. The green and red adaptation beams were provided by CS4-64 and CS2-59 colored glass filters (Corning Glass Co., Corning, NY, USA). The emerging light beam from the light guide was oriented so as to be on-axis for facets on the dorsal medial equatorial region. A series of quartz neutral-density filters attenuated the intensity of the test beam over 5 log units. The stimulus beams were interrupted by electronic shutters (Uniblitz, Vincent Associates, Rochester, NY, USA), which controlled the duration of the stimulus flash.
Intensity calibration
The quantum flux of the photic stimulus at different wavelengths across the spectrum was determined by placing a calibrated photodiode (SED 033, International Light, Inc., Newburyport, MA, USA) at the spot where the experimental eye was placed. The output of the photodiode was read on an electrometer (Model # IL 700, International Light, Inc., Newburyport, MA, USA).
Electrical recordings
The ERGs were recorded from dark-adapted intact beetles with a glass microelectrode (tip diameter ~10 μm) filled with insect Ringer’s (Muri and Jones 1983) and inserted underneath the corneal surface of the compound eye. The reference electrode was another Ringer-filled microelectrode inserted in the antenna. An Ag–AgCl pellet connected the microelectrodes to the input grid of a P16 Grass DC/AC preamplifier (Astro Instruments, Providence, RI, USA), the output of which was monitored on an oscilloscope (Model 5113, Tektronix, Inc, Beaverton, OR, USA) and displayed on an ink writer (RS 3200, Gould, Inc., Cleveland, OH, USA) for later evaluation and measurements. It is assumed that the responses we obtained arose primarily from the dorsal medial equatorial ommatidia, since that was the direction of the cone of the stimulating light beam.
Experimental procedures
Intact insects were immobilized on a cork with an electrical tape and the head was attached to the cork using a mixture of equal amounts of beeswax and rosin. The preparation was mounted on the recording stage and dark adapted for at least an hour prior to experimentation. Test flashes of 0.1 s were delivered at 0.5, 1 or 2 min intervals, permitting a complete recovery of the sensitivity of the eye after each flash. A standard flash of a selected wavelength (540 nm) and intensity (Neutral Density 2.0 filter) was periodically presented to monitor any overall changes in the sensitivity of the eye. Test flashes were delivered at 17 different wavelengths, which covered the spectrum from 360 to 683 nm in ~20 nm steps. The amplitude of the ERG provided an index of the sensitivity of the eye to the quantum flux and the spectral composition of the light stimulus.
In a few cases, ERGs were recorded over the entire range of available intensities for selected stimulus wavelengths, and intensity–response curves (i.e., V/log I functions) were obtained. The spectral sensitivity of the eye was obtained by determining the number of photons (Q) needed to elicit a criterion response across different stimulus wavelengths. The criterion amplitude chosen to construct spectral sensitivity varied from 0.5 to 2 mV. Precautions were taken such that the criterion amplitude was in the linear range of V/log I functions for each specific run. A plot of 1/Q as a function of wavelength gave the S(λ) function. All recordings were obtained during the animals’ photophase. All experiments were conducted at room temperature (21–23°C). These data were obtained during the months of November to February in 1996–1999 in the Departamento de Psicologia Experimental, Universidade de São Paulo, and in the months of November and December 1998–2000 and January 2001 in Washington, DC.
Bioluminescence
Specimens of P. divergens, P. punctatissimus, F. bruchii, and Pyrearinus termitilluminans were anesthetized with ethyl acetate for 1 min to obtain constant bioluminescence emission from both pro-thoracic and abdominal lanterns. The anesthetic procedure significantly minimizes the light fading effect associated with fluctuating O2 uptake levels through the spiracles and tracheal tubes. Spectra were acquired using a Spex Fluorolog 1681 with photomultiplier high voltage set to 750 V and 4.0 mm slit width of the monochromator. Prothoracic lantern spectra were measured by pointing the lantern segments toward the photomultiplier aperture. Prior to obtaining data, the insects were conditioned in the cuvette compartment with tape to minimize the fluctuations in emissions due to handling. The same procedure was used to efficiently expose the segment’s lantern to obtain the spectra of the abdominal lantern. All spectra were normalized to the maximum intensity of each scan and the points were fitted with an asymmetric double sigmoidal function using Mirocal Origin 4.1.
Results
Visual and screening pigments
In situ MSP scans were taken in the proximal regions of rhabdoms as described in “Materials and methods”. After actinic exposure of the dark-adapted retina to saturating red light, there was a decrease in absorbance at wavelengths above ~525 nm and a somewhat larger increase in absorbance between 400 and 525 nm as shown for P. divergens in Fig. 1a (jagged trace; average of 7 scans). The change suggests that the peak of a metarhodopsin is shifted 50 nm toward the shorter wavelengths from the original peak for a rhodopsin in the middle-wavelength region. Analysis of the difference spectrum for photoconversion (Fig. 1a, solid trace) indicates that the best-fitting pigment pair consists of a P539 rhodopsin and an M488 metarhodopsin with peak absorbance 1.85 times that of the rhodopsin. The data for Pyrophorus divergens (Fig. 1a) and P. punctatissimus (Fig. 1b) are similar. In P. punctatissimus, the analysis suggests the presence of a P543 rhodopsin and an M486 metarhodopsin (N = 9). Thus, it can be inferred that visual pigments, absorbing maximally near 540 nm (Fig. 1c), mediate visual sensitivity in the green part of the light spectrum in these species.

Difference spectra for photoconversion of visual pigment in the retinas of Pyrophorus (male) species. a P. punctatissimus. The jagged trace is an average of seven photoconversions, with absorption in the dark-adapted retina subtracted from that after a saturating actinic treatment with bright red light (Corning CS2-61 filter). The smooth trace is the best-fit difference spectrum, obtained using templates of a 539-nm rhodopsin and a 488-nm metarhodopsin with peak absorbance 1.85 times that of the rhodopsin. b P. divergens. The jagged trace is an average of nine photoconversions, but otherwise as in part a. The smooth trace is based on a 543-nm rhodopsin interconverting with a 486-nm metarhodopsin absorbing at 2.0 times that of the rhodopsin at the peak. c Absorption spectrum of the best-fit rhodopsin from P. punctatissimus (λ max at 539 nm) (dark trace) and the corresponding metarhodopsin (λ max at 488 nm) (light trace)
In the ventral sectors of the eyes of both P. punctatissimus and P. divergens, yellow screening pigments overlie the rhabdoms. The absorbance spectra of the screening pigments, while absorbing generally in the same part of the spectrum, differ somewhat between the two species (Fig. 2).

In situ absorbance spectrum of the screening pigment obtained from the ventral sector of the compound eyes in P. punctatissimus (thick line) and P. divergens (thin line) males. Each curve represents an average of five scans
Electroretinogram and visual spectral sensitivity
The electroretinogram recorded from the dorsal medial region of the compound eyes in Pyrophorus punctatissimus, P. divergens, F. bruchii, and Pyrearinus termitilluminans was an on-negative response similar to that described for fireflies Photuris versicolor (Lall 1981) and Photuris lucicrescens (Lall et al. 1982). There was no off-transient response.
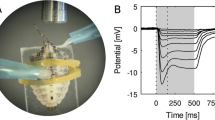
In a number of experiments, the amplitude of the ERGs was recorded for all stimulus wavelengths over the entire range of stimulus intensities. The amplitude of the response varied over a range from a low of 0.2 mV to a high of 5–6 mV. The data from one such experimental recording in F. bruchii is presented in Fig. 3. The Naka–Rushton function (Naka and Rushton 1966) as shown in the form below:
was fitted to our V/log I data, where I, R are an intensity–response ordered pair, R max is the asymptotic amplitude, K is the semi-saturation constant, and n is related to the slope. Our data (Fig. 3) were fitted to a single function where R max = 5.5 mV and n = 0.75. These data support the principle of univariance as postulated by Naka and Rushton (1966).

V/log I curves for selected wavelengths from one dark-adapted compound eye in Fulgeochlizus bruchii male. Abscissa log10 relative attenuation of the test intensity. Ordinate peak mV responses to 100 ms test flash. The curves are arbitrarily displaced along the x-axis for visibility. The numbers above the curves indicate the respective stimulus wavelengths. In each curve, the uppermost data point represents response to an unattenuated intensity of the stimulus. The subsequent data points are responses to different attenuation of stimulus intensity. Inset is the scale for the abscissa
The average S(λ) curves for P. punctatissimus, P. divergens, F. bruchii, and Pyrearinus termitilluminans are presented in Figs. 4 and 5. These S(λ) functions have a broad sensitivity peak in the green (λ max range 540–560 nm) with a shoulder in the near-UV. Since the recording electrode was placed in the dorsal medial region of the compound eye, the curves in Figs. 4a and 5 represent the average response from that region of the eye, except in P. termitilluminans (Fig. 4c) where the electrode placement was closer to the dorsal margin.

Upper panels electroretinographic spectral sensitivity functions in dark- (filled circles) and chromatic-adapted (filled triangles) compound eyes in males of Fulgeochlizus bruchii (a) and Pyrearinus termitilluminans (c). Error bars indicate ±1 SD. Lower panels The dark-adapted spectral sensitivity functions from a and c plotted on a percentage scale (the curve is fitted to B-spline by the Microcal Origin 4.1 computer software) and the species’ in vivo BL (dotted lines) from the ventral abdominal lantern in the male and the female for Fulgeochlizus bruchii (b) and Pyrearinus termitilluminans (d). For clarity of comparison between visual sensitivity and BL emission, the visual sensitivity (in % scale) is shown only from 460 to 680 nm in d

Upper panels electroretinographic spectral sensitivity functions in dark-adapted compound eyes in the males of Pyrophorus punctatissimus (a) and P. divergens (c). Different symbols (squares, inverted triangles and triangles) in a (diamonds, circles and triangles) and in b represent data averaged (N = 6–9) from different years. Error bars indicate ±1 SD. Lower panels A comparison between the average dark-adapted spectral sensitivity functions in upper panels plotted on a percentage scale (the curve is fitted to B-spline by the Microcal Origin 4.1 computer software) and the species’ in vivo BL from the ventral abdominal lantern in male and female P. punctatissimus (b), and P. divergens (d)
It should be pointed out that experiments were conducted during the photophase. It is known that in the nocturnal beetle Anaplognathus, the dark brown screening granules in the distal sleeve of superposition eyes remain in the light-adapted condition even when the eye is dark adapted during the day (Meyer-Rochow and Horridge 1975). The spectral absorbance of the dark brown screening pigment granules is known in firefly compound eyes (Lall et al. 1988). The extinction of these granules is very broad with a gradual rise in the short-wavelength region below 450 nm. We assume that the presence of the dark brown pigment granules during the photophase would not affect the spectral sensitivity in the long-wavelength region where the correspondence between species bioluminescence emission and S(λ) is shown in Figs. 4b, d, 5b, d, and 6.

A comparison of calculated absorbance of P540 overlaid with species screening pigment of 1.6 OD in Pyrophorus punctatissimus (a) and P. divergens (b) compared with species’ in vivo bioluminescence emission (dotted lines) from the ventral abdominal lantern of the female
Chromatic adaptation experiments were carried out to separate long- and short-wavelength (near-UV) mechanisms. Both green and red chromatic adaptation in F. bruchii (Fig. 4a) and P. termitilluminans (Fig. 4c) markedly reduced visual sensitivity in the long-wavelength region of the spectrum leaving a pronounced peak in the short-wavelength region, particularly in the near-UV.
Optical signaling from the ventral abdominal lantern in both the male and the female of the species is thought to serve in sexual communication (Colepicolo-Neto and Bechara 1984, 1986; Bechara 1988). We therefore compared the visual spectral sensitivity and the bioluminescence emission of the ventral abdominal lantern in both the male and the female. A correspondence exists between species bioluminescence emission and the broad green visual spectral mechanism, suggesting that this mechanism alone mediates the detection of the species’ bioluminescence (Figs. 4b, d, 5b, d) for mating.
Discussion
Near-UV and green spectral channels
The spectral sensitivity range from near-UV to green in click beetles (Figs. 4, 5) is a common feature of the compound eye of insects (reviews: Menzel and Backhaus 1991; Briscoe and Chittka 2001). The dark-adapted and chromatically adapted S(λ) functions strongly suggest the presence of independent green and near-UV spectral channels in the exocone eyes of F. bruchii (Fig. 4a) and P. termitilluminans (Fig. 4c). A comparable situation probably exists in P. punctatissimus and P. divergens. Similar experiments conducted on the compound eyes of North American fireflies also provided evidence for the presence of three spectral channels: near-UV, blue and green (Lall et al. 1982). The green spectral channel mediated by the P540 rhodopsin (Fig. 4) very likely is the dominant visual channel in the visible part of the spectrum for P. punctatissimus and P. divergens. The absorption spectrum of the near-UV rhodopsin was not obtained in our samples of the ventral retina, even though electrophysiologically such a receptor was isolated by chromatic adaptation (Fig. 4a, c).
Significance of the correlation between bioluminescence emissions and the spectral sensitivity functions
When we compare the ERG S(λ) functions with the peaks of species bioluminescence emissions from dorsal prothoracic and ventral abdominal lanterns, it is evident that the green spectral mechanism is an excellent detector of signals from both the prothoracic and ventral lanterns in both sexes in all four species. Thus far, the functional significance of bioluminescence of the dorsal prothoracic lanterns is unclear. They emit continuous light while the insect is in flight, so they may be used to illuminate the surroundings, as in fireflies (Lloyd 1968). They also light up when the insect is disturbed, so another function attributed to dorsal lanterns is that of a warning (aposematic) signal to potential predators. The selection of a green aposematic signal could be favored since most vertebrates and invertebrates have green receptors (Wald 1960; Briscoe and Chittka 2001); hence green bioluminescence emissions, therefore, would be detected easily by potential predators of click beetles. The selection of green rhodopsin (P540) would also maximize absolute sensitivity in green foliage habitats at night in accordance with the Sensitivity Hypothesis of Visual Ecology as was shown for North American fireflies (Seliger et al. 1982a, b).
Among North American fireflies, we found two types of screening pigments. In night-active Photuris versicolor and twi-night active (those species that initiate flashing at twilight and continue on into the night) P. potomaca, a yellow screening pigment, which broadly absorbs maximally in the short-wavelength region, has been located in the ventral sector of the compound eyes (Lall et al. 1988; Cronin et al. 2000). P545 rhodopsin overlaid by a 2.0 optical density yellow filter can account for the ERG S(λ) function obtained in P. versicolor (Cronin et al. 2000). Those species of North American fireflies that restrict their flashing activity for a short interval at twilight, i.e., twilight-active species were found to possess species-specific magenta screening pigments with narrow absorbance maximal in the green region of the spectrum (Lall et al. 1988; Cronin et al. 2000). It was established that rhodopsins ranging from P545 to P556 overlaid by the species-specific magenta screening pigment would account for the presence of narrow yellow S(λ) functions (Lall et al. 1988; Cronin et al. 2000). It was also shown that the narrow yellow S(λ) would be selected in accordance with the Contrast Hypothesis of Visual Ecology (Seliger et al. 1982a, b). The selection of yellow bioluminescence emissions would further optimize detection by narrow yellow S(λ) functions among twilight-active fireflies (Lall et al. 1980a, b, 1988; Seliger et al. 1982b).
We modeled the spectral sensitivities based on the absorbance of P540 rhodopsin (Fig. 1c) when overlaid by 1.6 optical density of the species screening pigment (Fig. 2) in the two species (Fig. 6). The calculation was not performed for wavelengths below 400 nm because absorbance data were not available in that region. The effect of the screening pigment is to narrow the sensitivity spectrum of the P540 visual pigment-containing photoreceptor class and to shift the sensitivity peak toward longer wavelengths, 555 and 562 nm in P. punctatissimus (Fig. 6a) and P. divergens (Fig. 6b), respectively. There exists correspondence between bioluminescence emission from the ventral abdominal lantern in each species and the calculated spectral sensitivity (Fig. 6). An analogous situation is found in those North American fireflies, which inhabit a similar photic environment. The nocturnal, green bioluminescence-emitting P. versicolor and yellow bioluminescence-emitting P. potomaca also possess a yellow screening pigment in their ventral retinas, as described earlier (Lall et al. 1988; Cronin et al. 2000).
The flashing activity period in P. punctatissimus, P. divergens, F. bruchii, and P. termitilluminans lasts for about 20–30 min just after sunset during the rainy season in Brazil. Field observations suggest that only males fly during courtship and the females are stationary on the vegetation. The female’s response brings the male to her and mating takes place on the foliage (Bechara 1988). Under such circumstances, the selective pressure would be to optimize detection of the optical signal by potential mates. During this short activity period, there is an appreciable amount of short-wavelength space light (Munz and McFarland 1977; Seliger et al. 1982b), which would act as environmental photon noise. It is postulated that the yellow screening pigments located in the ventral retina of P. punctatissimus and P. divergens (Fig. 2) absorb photons of this environmental space light (Johnsen et al. 2006).
Color vision
The question arises as to whether the discrimination of the color of bioluminescence from different lanterns is possible in the click beetles. For color discrimination to take place, the optical stimulus must differentially stimulate two receptors that differ with respect to their spectral sensitivities. The spectral sensitivity data (Figs. 4, 5) presented here clearly show the presence of near-UV and green receptor systems. The presence of a third, blue receptor system, which would allow for differential stimulation of deep green (535–540 nm) to lime-green (555–560 nm) optical stimuli, was not ascertained. However, it is not uncommon in ERG recordings to obtain dual peak S(λ) functions even when blue receptors are present in the eye (Lall 1981; Lall et al. 1982, 1988). The ERG is a mass response and therefore dominated by responses from green photoreceptors since they are in the majority in the ommatidia (Briscoe and Chittka 2001). Such was the situation in firefly data (Lall 1981; Lall et al. 1980b, 1982, 1988).
Kelber et al. (2002) have shown that color discrimination occurs in starlight illumination conditions for the elephant hawkmoth Deilelphila elpenor, and also in other nocturnal animals (Kelber and Roth 2006). Recently, Chuang et al. (2007, 2008) showed that nocturnal spiders, Nephila pilipes and Neoscona punctigera, attract prey (moths) with their bright yellow spots that act as visual lures at night. In a dark-adapted eye, where the screening pigment granules have moved between the corneal cones, the image of a distant object formed at the rhabdomeric segment is a summation of rays of light passing through all the facets that collect light from a given point in the field of view (Land 1981). Land estimates that superposition eyes are typically much more sensitive than apposition eyes. In click beetles, the image in the eye may well be bright enough to mediate color vision. The presence of color vision in lampyrids is postulated on the basis of behavioral evidence showing that the green–blue chromatic mechanism inhibits the male’s response to the female’s green bioluminescence light in the glowworm Lampyris noctiluca (Booth et al. 2004). In the firefly Photuris lucicrescens, near-UV, blue and green spectral mechanisms have been revealed by chromatic adaptation (Lall et al. 1982) and a blue receptor was also found in the dorsal margin of the eye in P. frontalis (Lall et al. 1988). A similar situation could exist in click beetles, where the presence of near-UV and green receptors is strongly suggested by chromatic adaptation experiments (Fig. 6), but a blue receptor is yet to be experimentally identified.
Conclusions
-
1.
In vespertine/nocturnal click beetles, the presence of a correspondence between bioluminescence emission and S(λ) functions mediated by P540 rhodopsin in conjunction with yellow screening pigment is similar to night-active fireflies.
-
2.
Both the deep green and lime-green bioluminescence emissions in the four elaterids studied here would be easily detected by the insects’ visual system. We propose that the ubiquitous green bioluminescence of the dorsal thoracic lanterns is used for illumination and as an interspecific warning signal against potential predators, whereas the ventral abdominal lantern signal is involved in intraspecific sexual communication.
-
3.
In the lampyrids, the different colors of bioluminescence represent an evolutionary selection to optimize the detection of the optical signal by the insect’s visual system under the constraints of the environmental photon noise (Seliger et al. 1982a, b). Our findings indicate that the two coleopteran families have followed similar courses in evolutionary selection for optical signaling.
Abbreviations
- BL:
-
Bioluminescence
- ERGs:
-
Electroretinograms
- MSP:
-
Microspectrophotometry
- Q :
-
Photons
- S(λ):
-
Spectral sensitivity
References
Bechara EJH (1988) Luminescent elaterid beetles: biochemical, biological, and ecological aspects. In: Baumstark AL (ed) Advances in oxygenated processes, vol 1. JAI Press Inc., London, pp 123–178
Booth D, Stewart AJA, Osorio D (2004) Color vision in the glow-worm Lampyris noctiluca (L.) (Coleoptera: Lampyridae): evidence for a green-blue chromatic mechanism. J Exp Biol 207:2373–2378
Briscoe AD, Chittka L (2001) The evolution of color vision in insects. Annu Rev Entomol 46:471–510
Chuang C, Yang E, Tso I (2007) Diurnal and nocturnal prey luring of a colorful predator. J Exp Biol 210:3830–3837
Chuang C, Yang E, Tso I (2008) Deceptive color signaling in the night: a nocturnal predator attracts prey with visual lures. Behav Ecol 19:237–244
Colepicolo-Neto P, Bechara EJH (1984) Bioluminescencia de insectos: Química e Biologia. Arq Biol Tecnol 27:439–464
Colepicolo-Neto P, Costa C, Bechara EJH (1986) Brazilian species of luminescent elaterid. Insect Biochem 16:803–810
Costa C (1971) Gênero Pyrophorus. 4. Redescrição e revalidação de Pyrophorus divergens Eschscholtz (Coleoptera, Elateridae). Pap Avulsos Zool (São Paulo) 24:65–72
Costa C (1975) Systematics and evolution of the tribes Pyrophorini and Heligimini with description of Campyloxeninae, new subfamily (Coleoptera, Elateridae). Arq Zool 26:49–191
Costa C (1982) Pyrearinus termitilluminans, sp. n., with description of the immature stages (Coleoptera, Elateridae, Pyrophorini). Rev Bras Entomol 1:23–30
Costa C (1991) New species of the genus Fulgeochlizus Costa, 1975 (Elateridae, Pyrophorinae). Rev Bras Entomol 35:567–572
Cronin TW, Järvilehto MVA, Weckström M, Lall AB (2000) Tuning of photoreceptor spectral sensitivity in fireflies (Coleoptera: Lampyridae). J Comp Physiol A 184:1–12
Eguchi E, Nemoto A, Meyer-Rochow VB, Ohba N (1984) A comparative study of spectral sensitivity curves in three diurnal and eight nocturnal species of Japanese fireflies. J Insect Physiol 30:607–612
Johnsen SA, Kelber A, Warrant EJ, Sweeney AM, Widder EA, Lee RL, Jernandez-André J (2006) Crepuscular and nocturnal illumination and its effects on color perception by the nocturnal hawkmoth Deilephila elpenor. J Exp Biol 209:789–800
Kelber A, Roth LSN (2006) Nocturnal colour vision—not as rare as we might think. J Exp Biol 209:781–788
Kelber A, Balkenius A, Warrant EJ (2002) Scotopic colour vision in nocturnal hawkmoths. Nature 419:922–925
Lall AB (1981) Electroretinogram and the spectral sensitivity of the compound eyes in the firefly Photuris versicolor (Coleoptera: Lampyridae): a correspondence between green sensitivity and species bioluminescence emission. J Insect Physiol 27:461–468
Lall AB, Chapman RM, Trouth CO, Holloway JA (1980a) Spectral mechanisms of the compound eye in the firefly Photinus pyralis (Coleoptera: Lampyridae). J Comp Physiol 135:21–27
Lall AB, Seliger HH, Biggley WH, Lloyd JE (1980b) Ecology of colors of firefly bioluminescence. Science 210:560–562
Lall AB, Lord ET, Trouth CO (1982) Vision in firefly Photuris lucicrescens (Coleoptera: Lampyridae): spectral sensitivity and selective adaptation in the compound eyes. J Comp Physiol 147:195–200
Lall AB, Strother GK, Cronin TW, Seliger HH (1988) Modification of spectral sensitivities by the screening pigments in the compound eyes of twilight-active fireflies (Coleoptera: Lampyridae). J Comp Physiol A 162:23–33
Lall AB, Ventura DF, Bechara EH, de Souza JM, Colepicolo-Neto P, Viviani VR (2000) Spectral correspondence between visual spectral sensitivity and bioluminescence emission spectra in the click-beetle Pyrophorus punctatissimus (Coleoptera: Elateridae). J Insect Physiol 46:1137–1141
Land MF (1981) Optics and vision in invertebrates. In: Autrum H (ed) Handbook of sensory physiology, vol VII 6B. Springer, Berlin, pp 471–592
Lloyd JE (1968) Illumination another function of firefly flashes? Entomol News 10:265–268
Menzel R, Backhaus W (1991) Color vision in insects. In: Gouras P (ed) The perception of color–vision and visual dysfunction. CRC Press, Boca Raton, pp 262–293
Meyer-Rochow VB, Horridge GA (1975) The eye of Anaplognathus (Coleoptera, Scarabaeidae). Proc R Soc Lond B 188:1–30
Munz FW, McFarland WN (1977) Evolutionary adaptations of fishes to the photic environment. In: Crescitelli F (ed) Handbook of sensory physiology, vol VII/5. Springer, Berlin, pp 193–274
Muri RB, Jones GJ (1983) Microspectrophotometry of single rhabdoms in the retina of the honeybee drone (Apis mellifera). J Gen Physiol 82:469–496
Naka KI, Rushton WAH (1966) S-potentials from color units in the retina of fish (Cyprinidae). J Physiol 185:536–555
Palacios AG, Goldsmith TH, Bernard GD (1996) Sensitivity of cones of a cyprinid fish (Danio aequipinnatus) to ultraviolet and visible light. Vis Neurosci 13:411–421
Seliger HH, Lall AB, Lloyd JE, Biggley WH (1982a) On the colors of firefly bioluminescence. I. An optimization model. Photochem Photobiol 36:673–680
Seliger HH, Lall AB, Lloyd JE, Biggley WH (1982b) On the colors of firefly bioluminescence. II. Experimental evidence for the optimization model. Photochem Photobiol 36:681–688
Wald G (1960) The distribution and evolution of visual systems. In: Florkin M, Mason HS (eds) Comparative Biochemistry, vol 1. Academic Press, New York, pp 311–345
Acknowledgments
The electrophysiological data on the click beetles were part of a master’s degree thesis of Alexandre A. Carvalho at the Universidade de São Paulo. This investigation was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) and the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPQ), and a Guggenheim Foundation fellowship to EJH Bechara; NSF grants IBN-118793 and IBN 0235820 to TW Cronin; National Institutes of Health grant SO6GM08016 to AB Lall; and grants to DF Ventura. We are grateful to Professora Cleide Costa (Museu de Zoologia, Universidade de São Paulo) for a critical reading of this manuscript and for providing us information on the geographical distribution of the Pyrophorini studied in South America. We are also indebted to anonymous reviewers for their critical suggestions to improve this MS and to Jean Lall for her excellent editorial support.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Lall, A.B., Cronin, T.W., Carvalho, A.A. et al. Vision in click beetles (Coleoptera: Elateridae): pigments and spectral correspondence between visual sensitivity and species bioluminescence emission. J Comp Physiol A 196, 629–638 (2010). https://doi.org/10.1007/s00359-010-0549-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00359-010-0549-x