
Abstract
The European grape berry moth is an important pest in vineyards. Males respond to the female-produced sex pheromone released from a piezo nebulizer in a dose-dependent manner in a wind tunnel: <50% arrive at the source at 5–50 pg/min (underdosed), 80% arrive at 100 pg/min to 10 ng/min (optimal) and <20% arrive at 100 ng/min (overdosed). Males responding to overdosed pheromone show in flight arrestment at 80 cm from the source. Host plant chemostimuli for Eupoecilia ambiguella increase the responses of males to underdosed and overdosed pheromone. (Z)-3-hexen-1-ol, (+)-terpinen-4-ol, (E)-β-caryophyllene and methyl salicylate released with the underdosed pheromone cause a significant increase in male E. ambiguella flying to the source. Time–event analysis indicates a positive correlation between faster activation and probability of source contact by the responding males. The four host plant compounds added to the overdosed pheromone permitted males to take off faster and with a higher probability of flying to the source. This suggests that perception of host plant products with the sex pheromone facilitates male E. ambiguella to locate females on host plants, lending credence to the hypothesis that plant products can signal rendezvous sites suitable for mating.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Olfaction is an important modality for insects serving inter- and intra-specific communication (Schoonhoven et al. 2005). Insects perceive a range of volatile products emitted by plants that serve them as signals in the search for food, oviposition sites and for shelter against desiccation and enemies (Syed and Guerin 2004; Schoonhoven et al. 2005). Host plant products can also serve phytophagous insects to locate rendezvous sites for mating (Landolt and Phillips 1997; Reddy and Guerrero 2004) whereby the probability to encounter the opposite sex may be enhanced. In this manner, host plant choice and mate choice are tightly linked. In addition, preferences for host plant odours can serve as a driving force for sympatric speciation (Bush 1969; Linn et al. 2003, 2005).
To date, numerous studies have reported on interactions between insect pheromones and host plant chemicals (for a review see Landolt and Phillips 1997). In one of the first studies Rahn (1968) showed that in the leek moth, Acrolepiopsis assectella, female-baited traps are most efficient to recapture released males in the presence of the host plant than with non host plants. Plant products can influence pheromone production, its release or its perception; sequestration of plant compounds as pheromone products is known, for example, in the moth species Utetheisa ornatrix and Grapholita molesta (Conner et al. 1981; Boppré 1986; Löfstedt et al. 1989). In female corn earworm moths, Helicoverpa zea, the production of sex pheromone is enhanced by a factor of 20–30 in the presence of host plants or the host plant volatiles ethylene, 3-methyl-butan-1-ol or phenyl acetaldehyde (Raina et al. 1992). However, for the perceiver of the pheromone, which in the case of moths is generally a male, enhancement and synergistic effects of plant volatiles on male attraction to pheromones have been reported in Cydia pomonella (Yang et al. 2004), Spodoptera exigua (Deng et al. 2004), H. zea (Light et al. 1993) and Heliothis virescens (Dickens et al. 1993). Such effects have only been investigated to date in a few moth species and knowledge about pheromone–host plant volatile interactions in moths is still scanty.
The European grape berry moth, Eupoecilia ambiguella Hb. (Lepidoptera, Tortricidae), is together with the grape vine moth, Lobesia botrana Den. & Schiff. (Lepidoptera, Tortricidae), one of the most important pests in European vineyards. On vine, this polyphagous species exhibits two flight periods per year. Larvae of the second generation feed on grapes and facilitate secondary infection of the fruit (Galet 1982). As in other moth species, female E. ambiguella release a sex pheromone to attract males (Arn et al. 1976, 1986). The sex pheromone is used in some pheromone-based integrated methods to control this pest (Rauscher and Arn 1979; Charmillot et al. 1995; Charmillot and Pasquier 2004). Analysis of the headspace odours over six host plant species lead to the identification of several host plant volatiles that elicit electroantennogram responses from male E. ambiguella antennae, i.e. (E)-2-hexenol, (Z)-3-hexenol, (Z)-3-hexenyl acetate, 1-octen-3-ol, R(+)-limonene, linalool, (E)-β-caryophyllene, (+)-terpinen-4-ol, 4,8-dimethyl-1(E),3,7-nonatriene (DMNT), benzaldehyde and methyl salicylate (Schmidt-Büsser et al. unpublished data). Here, we show how these host plant volatiles can affect the behavioural response of male E. ambiguella to its sex pheromone. Our hypothesis is that host plant compounds are used by E. ambiguella to mediate sexual communication as a strategy for optimised reproduction.
Materials and methods
Insects
Insects were held in a climate chamber at a 16:8 h L/D cycle at 65% RH and 25°C during the photophase and 85% RH, 18°C during the scotophase. The larval stages were reared on a semiartificial medium (Rauscher et al. 1984). Pupae were sexed and put on a gauze mesh over a dish filled with water for emergence. Each day males emerged into cages (BugDorm, 30 × 30 × 30 cm, MegaView Science Education Services Co., Taiwan) which were sprayed once a day with water and enclosed in plastic bags. A sugar solution (10% sucrose) in two glass vials closed with cotton wool wicks was offered to the males by placing two of them on the top of the cage.
Compounds tested
The synthetic pheromone compounds used were (Z)-9-dodecenyl acetate (Z9–12:Ac; 99.9%, Plant Research International, The Netherlands), dodecenyl acetate (12:Ac; >96%, BASF, Germany) and octadecyl acetate (18:Ac; >97%, TCI Europe N.V., Belgium). The commercially available plant volatiles hexanol (99%), (E)-β-caryophyllene (99%), methyl salicylate (>99%), R(+)-limonene (>98%), ±linalool (racemic mixture, >97%), (Z)-3-hexen-1-ol (>98%) and (+)-terpinen-4-ol (99%) were purchased from Fluka (Switzerland), 1-octen-3-ol (racemic mixture, >97%) from Merck (Germany), benzaldehyde (99%) and (E)-2-hexenal (>95%) from Sigma-Aldrich (Germany), and 4,8-dimethyl-1(E),3,7-nonatriene (DMNT) was supplied by Givaudan (Switzerland).
Behavioural experiments
The wind tunnel (200 × 60 × 60 cm) is constructed of non-reflecting glass (Syed and Guerin 2004). Two centrifugal ventilators at either end operate simultaneously to move the air across the tunnel at 30 cm/s through active charcoal cartridges and laminar flow screens (Rauscher et al. 1984). Overhead illumination is provided by high frequency fluorescent daylight tubes of 36 W (>1 kHz, Philips) running the length of the tunnel (≈5 lux on the floor). Below the tunnel floor, a white sheet with black patches of irregular sizes and shapes fixed at random provided visual cues. Humidity and temperature were the same as in the rearing chamber during the scotophase (above).
Three-day old male E. ambiguella were transferred individually into glass tubes (o.d. 2.5 cm, i.d. 2.1 cm; length 12.5 cm) closed with plastic caps at both ends. The glass tube was put on a stand 30 cm high and 30 cm from the downwind end of the wind tunnel, and the caps were removed on exposure to the odour source. Observations on the behaviour of individual males were quantified with a data logger (The Observer Version 5.0, Noldus, NL). The following behavioural elements were recorded following exposure to a treatment: duration of no activity, activation, take off, upwind flight, flying half the length of the wind tunnel (midline), close-in to within 10 cm of the source, and contact with the source. The observation of a male terminated either after 1 min when it was not activated or, otherwise, after 2 min.
A piezo nebulizer was used to deliver different treatments in the air as described by El-Sayed et al. (1999). Briefly, a glass capillary (5–6 cm long, o.d. 1 mm, i.d. 0.46 mm) with a drawn-out tip (2-3 cm) and an opening of 10–50 μm was connected via a PTFE microtube (1.5 m long, 1.02 mm o.d., 0.56 mm i.d., Hamilton, Switzerland) to a syringe containing an alcohol solution with known amounts of the test products. The solution was pumped to the glass capillary at a defined rate with a syringe pump (CMA 400, Microdialysis, Sweden). A piezo disc (Philips PXE5 25/2.0, Megatron AG, Switzerland) was connected by a clip (a modified piano chord) to the capillary. A frequency generator (FG-5000A Wavetex, Germany) at an amplitude of 40 V and a frequency of 90 kHz linked to the piezo disc caused the capillary tip to oscillate. In this manner, test solutions were released as an aerosol into the air stream. The capillary tip was protected from the arriving moths with a small metal mesh cage that was cleaned after exposure to each treatment. Ethanol (pro Analysi, Merck, Germany) was used as solvent for all volatiles tested as it had been shown by El-Sayed et al. (1999) to have no influence on the behaviour of E. ambiguella males in a wind tunnel.
The pheromone mixture used was a ternary mixture of Z9–12:Ac, 12:Ac and 18:Ac at a ratio of 1:1:2, as described by Arn et al. (1986). Seven different release rates (5 pg/min, 10 pg/min, 50 pg/min, 100 pg/min, 1 ng/min, 10 ng/min and 100 ng/min) of this ternary mixture were tested. The release rates of the pheromone referred to hereafter are with reference to the main pheromone component Z9–12:Ac. Plant volatiles were added either to an underdosed (5 or 50 pg/min) or to an overdosed (100 ng/min) level of the pheromone and released from the same piezo nebulizer.
To visualise the flight of male E. ambiguella in the wind tunnel we developed a 3D tracking system. An algorithm applied to the synchronised images obtained from two high speed cameras (Basler eXcite exA640-120m, Altrona Vision, Switzerland; with f = 6 mm objectives, CL-Electronics, Switzerland) determines the coordinates of a flying moth in 3D at a given time (Halcon, MVTec Software GmbH, Germany). Software (Crow, University of Neuchâtel) permits adjustment for parameters such as the initial calibration, luminosity and insect size. The two cameras were mounted on a stand at a distance of 60 cm from each other on one side of the wind tunnel. As the light settings of the wind tunnel were too low for the cameras to detect the flying moth we attached two lamps equipped with 25 W tungsten bulbs, reduced by a transformer to ≈11 W at the same angles as the cameras. These lamps shone on a retroreflective paper (Scotchlite 680-CR, 3 M, Switzerland) mounted on the back wall of the wind tunnel. Due to the reflected light the flying insect contrasted with the back wall permitting the cameras to record the flight coordinates at only marginally altered light settings for the moths. The flight of male moths was reconstructed from the recorded coordinates using the R package rgl (Version 2.4.1, http://www.r-project.org).
Statistical analysis
The responses of male E. ambiguella under each behavioural criterion recorded for different treatments, assumed to be binomially distributed, were compared by fitting a generalised linear model (GLM) with a logit link function (logistic regression) using the statistical package R (Version 2.4.1, http://www.r-project.org). Analysis of deviance based on the asymptotic distribution was used to test whether the flight responses were significantly dependent on the treatments. When the GLM showed significance (P < 0.05) multiple comparisons (R-Package: Multcomp, http://www.r-project.org) were made using Tukey-contrasts. Time–event analyses were performed using survival statistics, including proportional hazards (Cox) regression (Cox 1972), using the R-package survival (http://www.r-project.org). The flight times for defined distances in the wind tunnel in response to different treatments were compared by ANOVA (P < 0.05).
To compare the effect of plant volatiles on the attractiveness of underdosed pheromone release rates, odds ratios were calculated. An odd is the ratio of the probability that an event of interest, e.g. upwind flight or source contact, occurs divided by the probability that the event does not occur for a given treatment. The odds ratio is then established by comparison of the odds of two groups (Cox and Snell 1989; Bland and Altman 2000), for example, the ratio between the odds of males flying upwind in the presence of pheromone plus a plant volatile compared to the odds of males flying upwind to the pheromone alone. The odds ratio is a measure of how likely male E. ambiguella that are exposed to pheromone plus plant volatiles mixtures will fly upwind compared to males that are exposed to the pheromone alone. Odds ratios can have values between zero and infinity, where an odds ratio of one indicates no difference between two treatments, values lower than one a negative and values higher than one a positive effect of the plant volatile on the sex pheromone.
Results
Flight responses to different release rates of the ternary pheromone blend
The pheromone release rate of the ternary pheromone blend significantly influenced the different behavioural elements recorded for male E. ambiguella. The percentage of males responding increased to over 90% with dose to reach a plateau at 50 pg/min pheromone for activation and to between 70 and 80% at 100 pg/min for upwind flight (Fig. 1). For close-in and source contact, i.e. touching the metal mesh protecting the glass capillary, the dose-response curve followed an inverted U-profile in that the percentage of males flying close to and contacting the pheromone source increased with pheromone dose to plateau at 100 pg/min; at 10 ng/min pheromone 82% of the males still flew close to the source but only 34% did so at 100 ng/min. At this dose many males ceased upwind flight before arriving in the source vicinity and showed in-flight arrestment behaviour, indicating pheromone overdose. Flight tracks of two male E. ambiguella one flying to an optimal (10 ng/min) and the other to an overdosed (100 ng/min) pheromone level were recorded in 3D to visualise this phenomenon (Fig. 2; see the animation of the flight in 3D in supplementary material). This recording method only became available in this laboratory when this research project was due to finish.

Percentage of male E. ambiguella responding to different release rates of the ternary pheromone blend (Z9–12:Ac, 12:Ac and 18:Ac, 1:1:2) released from a piezo nebulizer. Most males flew to the ternary pheromone blend when released at between 100 pg/min and 10 ng/min. Different letters assigned within a behavioural element indicate statistically significant differences (logistic regression, P < 0.05)

3D reconstructions of male E. ambiguella flights to an optimal pheromone release rate of Z9–12:Ac, 12:Ac and 18:Ac at 1:1:2 of 10 ng/min (a) and to an overdosed level of the same ternary blend of 100 ng/min (b). The box represents the outline of the wind tunnel. In the presence of a high pheromone dose (b) the moth showed in-flight arrestment in the middle of the wind tunnel and was inhibited from flying to the source
As the behavioural response of male E. ambiguella under all behavioural criteria was already high at pheromone levels between 100 pg/min and 10 ng/min, we decided to study the effect of plant volatiles on the response to the pheromone at suboptimal doses, i.e. at the underdosed 5 and 50 pg/min and overdosed 100 ng/min levels.
Effects of plant volatiles added to an underdosed pheromone level
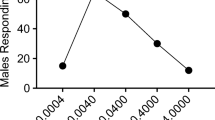
Initial experiments were made to determine levels at which plant volatiles could be effective when combined with the ternary pheromone blend. To this end, (+)-terpinen-4-ol was released at four levels ranging from 0.5 pg/min to 500 ng/min with the pheromone at 50 pg/min, i.e. a level of pheromone release that was suboptimal for upwind flight and source contact (above). Increasing doses of (+)-terpinen-4-ol induced higher numbers of upwind flights towards the pheromone source: at the pheromone to (+)-terpinen-4-ol ratio of 1:10,000 significantly more males flew half the length of the wind tunnel towards the source, closed-in on and contacted the source compared to the pheromone alone (Fig. 3a; logistic regression, P < 0.05).

Percentage of male E. ambiguella responding to an underdosed pheromone (phero) level (50 pg/min) presented alone and with (+)-terpinen-4-ol added at different release rates (a) and with either (Z)-3-hexen-1-ol, (E)-β-caryophyllene, methyl salicylate or benzaldehyde added at a pheromone–plant volatile ratio of 1:10,000 (b). Asterisks indicate a significant difference from the pheromone alone (logistic regression, P < 0.05)
(Z)-3-hexen-1-ol, (E)-β-caryophyllene, methyl salicylate and benzaldehyde were subsequently tested at the pheromone:plant volatile ratio of 1:10,000 with each host volatile released at 500 ng/min (pheromone at 50 pg/min). In presence of (E)-β-caryophyllene the response level of the whole behavioural sequence from activation to source contact by males was significantly increased compared to the pheromone alone and source contacts by males rose from 13 to 41% (Fig. 3b; logistic regression, P < 0.05). Both (Z)-3-hexen-1-ol and methyl salicylate released at 500 ng/min significantly increased the number of male E. ambiguella closing in on and contacting the pheromone source from 13 to 30 and 27%, respectively (Fig. 3b). With the pheromone:benzaldehyde mixture slightly fewer males were activated and made upwind flights compared to the pheromone alone, but more of these males arrived at the source; these differences were not significant (Fig. 3b).
The attractiveness of the pheromone presented alone was regularly controlled for these experiments. In later experiments, however, male responses to the 50 pg/min release rate increased significantly to almost 60% of upwind flights compared to 28% obtained in the earlier experiments. The release rate of the pheromone was decreased to 5 pg/min for subsequent tests with the plant volatiles (E)-2-hexen-1-al, 1-octen-3-ol, linalool and limonene each released at 50 ng/min to maintain the pheromone:plant volatile ratio at 1:10,000 as for experiments with the other five plant volatiles (above). With the exception of 1-octen-3-ol, where all behavioural steps dropped, all the other plant compounds served to increase the percentage of males contacting the source but not significantly (Table 1). To compare the different plant volatile treatments based on a pheromone release rate of either 5 or 50 pg/min, odds ratios for the pheromone-plant compound mixture and the pheromone blend alone were calculated (Table 1). A general pattern emerged, namely, that all plant volatiles tested in combination with the pheromone, with the exception of 1-octen-3-ol, had a positive effect on the percentage of males flying up the wind tunnel, closing in on and contacting the source, with (E)-β-caryophyllene and (+)-terpinen-4-ol having the strongest effects.
To investigate effects of plant volatile mixtures on male E. ambiguella pheromone responses a mixture of (Z)-3-hexen-1-ol, (E)-β-caryophyllene and methyl salicylate each released at 50 ng/min and the pheromone at 5 pg/min was tested. No significant difference in the response to the pheromone alone and the pheromone mixed with these three plant volatiles was recorded (Table 1; logistic regression). As this plant volatile mixture did not improve the attractiveness of the pheromone to E. ambiguella males a different approach to test plant volatile mixtures was taken. First, the response to (E)-β-caryophyllene released at different levels with the pheromone was recorded in order to establish if a particular ratio was critical (this compound performed best in combination with the pheromone; Table 1). Pheromone:(E)-β-caryophyllene ratios of 1:1, 1:1,000 and 1:100,000 were tested at a pheromone release rate of 5 pg/min and compared to the 1:10,000 ratio tested above at 50 pg/min pheromone using odds ratios. (E)-β-caryophyllene released at the same rate as the pheromone induced no significant difference in the response of male E. ambiguella compared to the pheromone alone (Table 1). Increasing the release rate of (E)-β-caryophyllene 1,000-fold, significantly more males flew upwind to the source (35%) compared to the pheromone alone (20%; P < 0.05) and the numbers of males contacting the source also increased but not significantly (19% compared to 9%; Table 1). At the highest release rate of 500 ng/min (E)-β-caryophyllene, more males flew upwind and contacted the source compared to the pheromone alone, but only the number of males flying over half the length of the wind tunnel differed significantly from the pheromone alone (Table 1). The pheromone:(E)-β-caryophyllene ratio of 1:10,000 had the strongest effect in attracting males to the source although as many males also flew upwind to the 1:1,000 ratio (Table 1). However, the 1:10,000 ratio proved most effective in inducing source contacts.
(Z)-3-hexen-1-ol was then released with the 1:1,000 pheromone:(E)-β-caryophyllene combination (at 5 pg and 5 ng/min, respectively) at four different release rates (50 pg/min, 500 pg/min, 5 ng/min and 50 ng/min). Best attraction of E. ambiguella males was achieved at a pheromone:(E)-β-caryophyllene:(Z)-3-hexen-1-ol ratio of 1:1,000:100 (Fig. 4; Table 1) where 48% of the males flew upwind, more than to the pheromone alone (20%, P < 0.05) or to the 1:1,000 pheromone:(E)-β-caryphyllene mixture (35%). When (Z)-3-hexen-1-ol was released at 5 ng/min in the mixture the response was the same as the response to the pheromone:(E)-β-caryophyllene mixture of 1:1,000. The lowest and highest (Z)-3-hexen-1-ol release rates tested in these mixtures (50 pg/min and 50 ng/min, respectively) did not cause significant differences in male responses compared to the pheromone alone. In summary, comparing the odds ratios of all plant volatile–pheromone combinations tested (Table 1) the 1,000:100 mixture of (E)-β-caryophyllene and (Z)-3-hexen-1-ol had the strongest effect on the percentage of male grape berry moths flying to an underdosed pheromone source in the wind tunnel.

Percentage of male E. ambiguella responding to an underdosed pheromone (phero) level (5 pg/min), in mixtures with (E)-β-caryophyllene (β-caryo) released at 5 ng/min and to this binary mixture with (Z)-3-hexen-1-ol (Z3hex) released at different levels. Different letters within a behavioural element indicate statistically significant differences between treatments (logistic regression, P < 0.05); n = 80 for each treatment
Time–event analysis of behavioural responses
In the experiments conducted with the pheromone released at 50 pg/min and the plant volatiles at 500 ng/min, in addition to recording the percentage of males responding, the time at which each behavioural element occurred was also recorded. A time–event analysis of male activation showed that they were activated significantly faster with either (+)-terpinen-4-ol or (E)-β-caryophyllene released at 500 ng/min with the pheromone compared to the pheromone alone (proportional hazards (Cox) regression, P < 0.05; Fig. 5). Furthermore, a correlation between the time to activation and probability of source contact was found as earlier activated males were more likely to fly upwind and contact the source than later activated ones (logistic regression, P < 0.01). However, this is a general effect and no direct interaction between the time to activation and odour treatment on the probability of source contact was found.

Cumulative percentage of male E. ambiguella activated over a time period of 60 s in response to its underdosed pheromone (phero) presented alone at 50 pg/min, with (+)-terpinen-4-ol added at different release rates (a) and with (Z)-3-hexen-1-ol, (E)-β-caryophyllene, methyl salicylate or benzaldehyde released at 500 ng/min (b). Males were activated significantly faster with (+)-terpinen-4-ol and (E)-β-caryophyllene released at 500 ng/min and in higher numbers compared to the pheromone alone (proportional hazards (Cox) regression, P < 0.05)
Plant volatiles also influenced flight time. The mean flight time from the downwind quarter to different points upwind in the wind tunnel, i.e. midline, third upwind quarter, 10 cm before the source and source contact, was compared for the pheromone alone with plant compounds added to the pheromone. In general, the males flew in a shorter time to the source with (Z)-3-hexen-1-ol, benzaldehyde, methyl salicylate and (E)-β-caryophyllene added at 500 ng/min, and this effect was significant for the latter two plant compounds (ANOVA, P < 0.05; Fig. 6). Such an effect on the flight time was not found with (+)-terpinen-4-ol (data not shown).

Mean flight time of male E. ambiguella in different segments of the wind tunnel in response to the pheromone (phero) alone (n = 160) and with either (Z)-3-hexen-1-ol (Z3hex; n = 80), methyl salicylate (ms; n = 80), (E)-β-caryophyllene (β-caryo; n = 160) or benzaldehyde (benzald; n = 80) released at a pheromone:plant compound ratio of 1:10,000. First segment flight from the first quarter to half the length of the wind tunnel distance; second segment flight from the middle to the third quarter; third segment flight from the third quarter to within 10 cm of the source; fourth segment flight time from close-in at 10 cm from the source to source contact. In the presence of methyl salicylate and (E)-β-caryophyllene males needed significantly less time to fly from the first quarter of the wind tunnel to the source compared to the pheromone alone (ANOVA, P < 0.05)
Effects of plant volatiles on behavioural responses to an overdosed pheromone level
At high pheromone levels (100 ng/min) most males flew upwind to within a certain distance of the source, but then showed in-flight arrestment or flew out of the odour plume (Figs. 1, 2). Only a few males flew to within 10 cm of the source and made contact (Fig. 1). To investigate a possible influence of host volatiles on in-flight arrestment a headspace extract of the rearing medium, (+)-terpinen-4-ol, methyl salicylate, 1-hexanol, DMNT, (Z)-3-hexen-1-ol and 1-octen-3-ol were released at 100 pg/min, 10 ng/min and 1 μg/min and (E)-β-caryophyllene at only 100 pg/min with the overdosed pheromone at 100 ng/min. In general, the plant volatiles released at 1 μg/min had no influence on the number of males that flew to within 10 cm of the overdosed pheromone (Fig. 7). Four of the plant volatiles, namely methyl salicylate, 1-hexanol, DMNT, 1-octen-3-ol, and the headspace extract of the rearing medium had no significant effect on the percentage of male grape berry moths flying to within 10 cm of the source at any of the release rates tested. However, addition of (+)-terpinen-4-ol or (Z)-3-hexen-1-ol released at either 10 ng/min or 100 pg/min and (E)-β-caryophyllene released at 100 pg/min with the overdosed pheromone changed male flight behaviour by rendering the overdosed pheromone dose attractive over the full length of the wind tunnel. This resulted in significantly more males flying to within 10 cm of the overdosed pheromone source at release levels of these three host plant volatiles of 0.1 and 0.001 times the pheromone (logistic regression, P < 0.05).

Behavioural elements performed by male E. ambiguella to an overdosed pheromone release rate (100 ng/min) presented alone and in mixtures with different plant volatiles at pheromone to plant volatile ratios of 1:10, 1:0.1 and 1:1:0.001. (+)-Terpinen-4-ol and (Z)-3-hexen-1-ol released at 10% of the pheromone release rate and these two compounds or (E)-β-caryophyllene released at 0.1% of the pheromone served to restore close-in and contact with the source (logistic regression, P < 0.05; asterisks indicate a significant difference from the pheromone alone)
Since the males were activated immediately on exposure to the overdosed pheromone level, we performed a time–event analysis starting with the next behavioural element, i.e. “take-off”, for males exposed to the pheromone alone and for pheromone:plant volatile mixtures at a 1:0.001 ratio. In the presence of (Z)-3-hexen-1-ol, 1-octen-3-ol, (E)-β-caryophyllene, (+)-terpinen-4-ol and methyl salicylate the males took off faster compared to the pheromone alone, and this difference was significant for (+)-terpinen-4-ol and methyl salicylate (Fig. 8; proportional hazards (Cox) regression, P < 0.05).

Cumulative percentage of male E. ambiguella taking off over a time period of 120 s in response to the overdosed pheromone alone (100 ng/min) and to pheromone:plant volatile mixtures of 1:0.001. Males took of significantly faster in the presence of (+)-terpinen-4-ol and methyl salicylate compared to the pheromone alone (Cox proportional hazards model, P < 0.05; asterisks indicate a significant difference from the pheromone alone)
Discussion
Dose-dependent response to the ternary pheromone blend
Male E. ambiguella responded to the ternary pheromone blend of Z9–12:Ac, 12:Ac and 18:Ac at a 1:1:2 ratio in a dose-dependent manner. For all behavioural elements recorded, the percentage of males responding increased with increasing dose before reaching a plateau of optimal attraction, as already described by Arn et al. (1986). The optimal pheromone loads on rubber septa reported by these authors at between 100 and 500 μg, resulting in a release rate of between 1 and 10 ng/min (after McDonough et al. 1989), correspond well to the optimal pheromone release rates of between 0.1 and 10 ng/min we found here using the piezo nebulizer to release the pheromone. While the percentage of activated males and those undertaking upwind flight remained at high levels even at the highest pheromone release rate of 100 ng/min, the percentage undertaking “close in” and “source contact” dropped significantly. These males flew upwind only for a certain distance and then showed in-flight arrestment behaviour (Fig. 2b; for animation see supplementary material). In-flight arrestment behaviour in response to very high pheromone doses is probably linked to the plume structure and dose. As an odour plume is not continuous but intermittent, upwind flying males encounter bursts of odour alternating with clean air (Murlis and Jones 1981; Kaissling 1997). The density of such bursts will be higher near the source than farther downwind such that males can encounter concentrations that are overdosed, and, consequently, show in-flight arrestment behaviour. In addition, the antennae and the body surface of the male probably pick up pheromone at high release rates causing a constant release and excitation of olfactory receptor neurones, further reducing the male’s ability to detect differences in pheromone levels in the air.
Host plant volatiles influence male E. ambiguella flight responses to its pheromone
Since 80% of male grape berry moths already flew to the pheromone source at the optimal release rate (Fig. 1), it would have been difficult to estimate any additional effect of plant compounds. For this reason, we studied possible interactions between these two groups of compounds at suboptimal pheromone levels, i.e. at underdosed and overdosed levels. The addition of the host plant chemostimuli (Z)-3-hexen-1-ol, (+)-terpinen-4-ol and (E)-β-caryophyllene to the overdosed pheromone resulted in fewer males showing in-flight arrestment and restored the close-in behaviour on the semiochemical source. In addition, male grape berry moths took off significantly faster in the presence of (+)-terpinen-4-ol and methyl salicylate (Fig. 8). The phenomenon of restoring activity of overdosed pheromone level has already been documented in this species, where the addition of 12:Ac and 18:Ac to an overdosed level of Z9–12:Ac restored its attraction to males, thus increasing the range of doses over which the pheromone is attractive (Rauscher et al. 1984; Arn et al. 1986). Interestingly, neither 12:Ac nor 18:Ac is attractive on its own. Rauscher et al. (1984) hypothesised that compounds added to the overdosed pheromone have a dilution effect by competing with the pheromone at the receptor level leading to stimulation similar to that induced at lower levels. Plant volatiles may alter the manner in which chemostimuli arrive at the receptor cell.
Addition of the same plant volatiles that influenced male behavioural responses to the overdosed pheromone resulted in at least three behavioural effects at an underdosed pheromone level: faster activation, a shorter flight time to the odour source and an increased percentage of males flying to the pheromone source (Figs. 3, 4, 5, 6; Table 1). The latter effect of plant compounds on moth pheromones has already been reported. The addition of either linalool, (E)-β-farnesene, or (Z)-3-hexen-1-ol increased the percentage of male C. pomonella flying upwind to codlemone by 20–30% in wind tunnel studies (Yang et al. 2004). In the field, trap captures of male codling moth and H. zea were significantly increased when the pheromone was dispensed with a mixture of green leaf volatiles (Light et al. 1993). An increase in pheromone attraction by the addition of plant volatiles is also reported for H. virescens and S. exigua (Dickens et al. 1993; Deng et al. 2004). What is novel in our study is that the host plant chemostimuli are shown to reduce reaction time and flight time of responding males to the pheromone. The implication is that plant volatiles can decrease the response threshold of male E. ambiguella to its pheromone. Faster reaction to the pheromone in the presence of host plant compounds is of functional significance, since we have shown here that earlier activated males are more likely to fly to the source. Whether the shorter flight time of such males is due to increased flight velocity or to more directed flight remains open. Differences in the quality of upwind flights depending on the stimulus source were shown by Witzgall and Arn (1990), where straighter flights to calling females were recorded for male L. botrana than to an artificial pheromone source.
Despite the fact that plant compounds presented singly can affect the behavioural response of male E. ambiguella to its sex pheromone, in nature, insects deal with complex mixtures of products released from plants. Mixtures of plant compounds would therefore be expected to have stronger effects on pheromone perception by male E. ambiguella than products presented singly with the pheromone. Our results support this as a specific mixture of β-caryophyllene and (Z)-3-hexen-1-ol tends to enhance the attractiveness of the pheromone more than either of these plant volatiles added singly to the pheromone.
Overall, our results emphasise the role plant volatiles play in sexual communication between male and female moths in that mate choice in moths could be dependent on host selection, as in other insect species. Females call and oviposit on host plants (Landolt and Phillips 1997; Schoonhoven et al. 2005) and so can be located with a high probability at such places. Males could arguably be at mating sites before females start calling by following host plant cues. The fact that male E. ambiguella emerge before females is pertinent. In addition, when a female moth is calling while sitting on the host plant, her body may pick up host plant products. Plant products such as nonanal and (E, E)-farnesyl acetate were found, for example, on the scales of female C. pomonella (DeLury et al. 1999). Furthermore, the sex pheromone is accompanied by plant volatiles when it encounters the male antennal olfactory receptor cells and males may use such cues in order to optimise the chance of encountering a female to reproduce. As a strategy to locate females more efficiently male forest cockchafers Melolontha hippocastani, for example, are not only attracted to the female released sex attractant, 1,4-benzoquinone, but also to green leaf volatiles released by the leaves of deciduous trees fed on by the females (Ruther et al. 2002).
In physiological terms, plant compounds can influence the pheromone responses of E. ambiguella males at different perception steps, i.e. at the periphery, in the antennal lobe and in higher brain centres. At the periphery, an interaction between plant volatiles and pheromone products was found in H. zea (Ochieng et al. 2002) where the firing rate of sex pheromone olfactory receptor neurones in males is significantly enhanced by simultaneous stimulation with linalool and (Z)-3-hexenol. In Bombyx mori, projection neurones innervating the toroid glomerulus in the macroglomerular complex fire at an increased rate when the male antenna is stimulated by a mixture of (Z)-3-hexen-ol and bombykol compared to a stimulation with either of these products alone (Namiki et al. 2008).
Males have to deal with different pheromone levels in vineyards: low levels as emitted by females or from pheromone dispensers and detected at a distance, and higher doses in the air near dispensers carrying high pheromone loads used for mating disruption. Our results suggest that host plant chemostimuli can serve to increase the active space of a pheromone source. Underdosed pheromone is rendered more attractive, suggesting that the detection threshold can be lowered by the presence of the host plant semiochemicals. At the other extreme, high pheromone loads induce males to fly persistently to within a distance of the source but without contacting it as shown here. Adding plant semiochemicals to such sources permits males to continue upwind flight to contact the source.
Pheromone:plant volatile ratios
At a 1:10,000 pheromone to host plant volatile ratio, significantly more males were attracted to the underdosed pheromone in the wind tunnel with (Z)-3-hexen-1-ol, (E)-β-caryophyllene, (+)-terpinen-4-ol and methyl salicylate.. The plant volatile to pheromone ratio we tested here is higher compared with those found to affect responses to pheromones in other tortricids. More C. pomonella males flew to the source at a pheromone:plant volatile ratio between 1:10 and 1:100, but a higher ratio of 1:10,000 did not enhance the response in wind tunnel tests (Schmera and Guerin, in preparation; Yang et al. 2004). A codlemone:pear ester ratio of 1:10,000 on rubber septa even served to decrease male attraction in the wind tunnel (Yang et al. 2005). In L. botrana host plant volatiles affect responses to the sex pheromone at a ratio of 1:1,000 (von Arx et al., in preparation), but the effect disappeared at a higher (1:100,000) and lower (1:10) release ratio. Thus, other plant compounds tested here with no significant effect at a 1:10,000 pheromone to plant compound ratio could still contribute to better attraction of male E. ambiguella to the pheromone at other ratios.
Appropriate ratios are also critical when testing the effects of mixtures of host plant volatiles on the response to the sex pheromone. Best attraction was achieved with a release rate of (Z)-3-hexen-1-ol ten times lower than that of (E)-β-caryophyllene. Other (Z)-3-hexen-1-ol:(E)-β-caryophyllene ratios as with a mixture of (Z)-3-hexen-1-ol, (E)-β-caryophyllene and methyl salicylate at 10,000:10,000:10,000 added to the pheromone caused the effect of the individual compounds to disappear to a response level no different to the underdosed pheromone alone. The attractiveness of mixtures of plant volatiles has been extensively studied for female C. pomonella and L. botrana by Ansebo et al. (2004) and Tasin et al. (2005, 2006a, b, 2007) with the conclusion that the ratio between host plant volatiles is critical. A mixture of the three most abundant volatiles at the ratio occurring over grape berries, i.e. (E)-β-caryophyllene, DMNT, and (E,E)-α-farnesene at 100:78:9, was attractive to female L. botrana whereas another mixture of these three compounds at 37:17:100 found over apples was not (Tasin et al. 2006b, 2007). Although these studies and our own underline the importance of appropriate ratios, one could expect the ratio of plant volatiles needed for attraction to be less strict than the ratio of components in a species specific pheromone blend considering the range of host plants exploited. Furthermore, the ratios of plant products are highly dependent on factors such as the physiological condition of the host plant and the prevailing environmental conditions.
Abbreviations
- DMNT:
-
4,8-Dimethyl-1(E),3,7-nonatriene
- Z9–12:Ac:
-
(Z)-9-Dodecenyl acetate
- 12:Ac:
-
Dodecenyl acetate
- 18:Ac:
-
Octadecyl acetate
References
Ansebo L, Coracini MDA, Bengtsson M, Liblikas I, Ramirez M, Borg-Karlson AK, Tasin M, Witzgall P (2004) Antennal and behavioural response of codling moth Cydia pomonella to plant volatiles. J Appl Entomol 128:488–493
Arn H, Rauscher S, Buser HR, Roelofs WL (1976) Sex pheromone of Eupoecilia ambiguella—cis-9-dodecenyl acetate as a major component. Z Naturforsch C 31:499–503
Arn H, Rauscher S, Buser HR, Guerin PM (1986) Sex-pheromone of Eupoecilia ambiguella female—analysis and male response to ternary blend. J Chem Ecol 12:1417–1429
Bland JM, Altman DG (2000) The odds ratio. Br Med J 320:1468
Boppré M (1986) Insects pharmacophagously utilizing defensive plant chemicals (pyrrolizidine alkaloids). Naturwissenschaften 73:17–26
Bush GL (1969) Sympatric host race formation and speciation in frugivorous flies of the genus Rhagoletis (Diptera, Tephritidae). Evolution 23:237–251
Charmillot PJ, Pasquier D (2004) Isonet: une nouvelle gamme de diffuseurs pour la lutte par confusion contre les vers de la grappe. Revue Suisse Vitic Arboric Hortic 36:95–100
Charmillot PJ, Pasquier D, Scalco A (1995) Lutte par confusion contre les vers de la grappe eudémis et cochylis à Perroy et Allaman: résultats de 1995. Revue Suisse Vitic Arboric Hortic 27:347–358
Conner WF, Eisner T, Vandermeer RK, Guerrero R, Meinwald J (1981) Precopulatory sexual interaction in an arctiid moth (Utetheisa ornatrix): role of a pheromone derived from dietary alkaloids. Behav Ecol Sociobiol 9:227–235
Cox DR (1972) Regression models and life-tables. J R Stat Soc B 34:187–220
Cox DR, Snell EJ (1989) Analysis of binary data. Chapman & Hall, London
DeLury NC, Gries R, Gries G, Judd GJR, Khaskin G (1999) Moth scale-derived kairomones used by egg-larval parasitoid Ascogaster quadridentata to locate eggs of its host, Cydia pomonella. J Chem Ecol 25:2419–2431
Deng JY, Wei HY, Huang YP, Du JW (2004) Enhancement of attraction to sex pheromones of Spodoptera exigua by volatile compounds produced by host plants. J Chem Ecol 30:2037–2045
Dickens JC, Smith JW, Light DM (1993) Green leaf volatiles enhance sex attractant pheromone of the tobacco budworm, Heliothis virescens (Lep.: Noctuidae). Chemoecology 4:175–177
El-Sayed A, Gödde J, Arn H (1999) Sprayer for quantitative application of odor stimuli. Environ Entomol 28:947–953
Galet P (1982) Les maladies et les parasites de la vigne. Impr. du Paysan du Midi, Montpellier
Kaissling K-E (1997) Pheromone-controlled anemotaxis in moths. In: Lehrer M (ed) Orientation and communication in arthropods. Birkhauser Verlag, Basel
Landolt PJ, Phillips TW (1997) Host plant influences on sex pheromone behavior of phytophagous insects. Annu Rev Entomol 42:371–391
Light DM, Flath RA, Buttery RG, Zalom FG, Rice RE, Dickens JC, Jang EB (1993) Host-plant green leaf volatiles synergize the synthetic sex pheromone of the corn earworm and codling moth (Lepidoptera). Chemoecology 4:145–152
Linn C, Feder JL, Nojima S, Dambroski HR, Berlocher SH, Roelofs W (2003) Fruit odor discrimination and sympatric host race formation in Rhagoletis. Proc Natl Acad Sci USA 100:11490–11493
Linn CE, Dambroski HR, Nojima S, Feder JL, Berlocher SH, Roelofs WJ (2005) Variability in response specificity of apple, hawthorn, and flowering dogwood-infesting Rhagoletis flies to host fruit volatile blends: implications for sympatric host shifts. Entomol Exp Appl 116:55–64
Löfstedt C, Vickers NJ, Roelofs CJ, Baker TC (1989) Diet related courtship success in the Oriental fruit moth, Grapholita molesta (Tortricidae). Oikos 55:402–408
McDonough LM, Brown DF, Aller WC (1989) Insect sex pheromone: effect of temperature on evaporation rate of acetates from rubber septa. J Chem Ecol 15:779–790
Murlis J, Jones CD (1981) Fine-scale structure of odour plumes in relation to insect orientation to distant pheromone and other attractant sources. Physiol Entomol 6:71–86
Namiki S, Iwabuchi S, Kanzaki R (2008) Representation of a mixture of pheromone and host plant odor by antennal lobe projection neurons of the silkmoth Bombyx mori. J Comp Physiol A Sens Neural Behav Physiol 194:501–515
Ochieng SA, Park KC, Baker TC (2002) Host plant volatiles synergize responses of sex pheromone-specific olfactory receptor neurons in male Helicoverpa zea. J Comp Physiol B Biochem Syst Environ Physiol 188:325–333
Rahn R (1968) Effect of host plant on sexual attraction in Acrolepia assectella Zeller (Lep. Plutellidae). C R Hebd Seances Acad Sci D 266:2004–2006
Raina AK, Kingan TG, Mattoo AK (1992) Chemical signals from host plant and sexual behavior in a moth. Science 255:592–594
Rauscher S, Arn H (1979) Mating suppression in tethered females of Eupoecilia ambiguella by evaporation of (Z)-9-dodecenyl acetate in the field. Entomol Exp Appl 25:16–20
Rauscher S, Arn H, Guerin P (1984) Effects of dodecyl acetate and Z-10-tridecenyl acetate on attraction of Eupoecilia ambiguella males to the main sex pheromone component, Z-9-dodecenyl acetate. J Chem Ecol 10:253–264
Reddy GVP, Guerrero A (2004) Interactions of insect pheromones and plant semiochemicals. Trends Plant Sci 9:253–261
Ruther J, Reinecke A, Hilker M (2002) Plant volatiles in the sexual communication of Melolontha hippocastani: response towards time-dependent bouquets and novel function of (Z)-3-hexen-1-ol as a sexual kairomone. Ecol Entomol 27:76–83
Schoonhoven LM, van Loon JJA, Dicke M (2005) Insect-plant biology—from physiology to evolution. Oxford University Press, New York
Syed Z, Guerin PM (2004) Tsetse flies are attracted to the invasive plant Lantana camara. J Insect Physiol 50:43–50
Tasin M, Anfora G, Ioriatti C, Carlin S, De Cristofaro A, Schmidt S, Bengtsson M, Versini G, Witzgall P (2005) Antennal and behavioral responses of grapevine moth Lobesia botrana females to volatiles from grapevine. J Chem Ecol 31:77–87
Tasin M, Bäckmann A, Bengtsson M, Varela N, Ioriatti C, Witzgall P (2006a) Wind tunnel attraction of grapevine moth females, Lobesia botrana, to natural and artificial grape odour. Chemoecology 16:87–92
Tasin M, Bäckmann A, Bengtsson M, Ioriatti C, Witzgall P (2006b) Essential host plant cues in the grapevine moth. Naturwissenschaften 93:141–144
Tasin M, Bäckman A-C, Coracini M, Casado D, Ioriatti C, Witzgall P (2007) Synergism and redundancy in a plant volatile blend attracting grapevine moth females. Phytochemistry 68:203–209
Witzgall P, Arn H (1990) Direct measurement of the flight behavior of male moths to calling females and synthetic sex-pheromones. Z Naturforsch C 45:1067
Yang ZH, Bengtsson M, Witzgall P (2004) Host plant volatiles synergize response to sex pheromone in codling moth, Cydia pomonella. J Chem Ecol 30:619–629
Yang ZH, Casado D, Ioriatti C, Bengtsson M, Witzgall P (2005) Pheromone pre-exposure and mating modulate codling moth (Lepidoptera: Tortricidae) response to host plant volatiles. Agricul Forest Entomol 7:231–236
Acknowledgments
We thank Dr. Paul Becher and Dr. Denes Schmera (University of Neuchâtel, Switzerland) and Dr. Pierre-Joseph Charmillot, Françoise Briand and Dr. Thomas Degen (Agroscope Changins-Wädenswil, Nyon, Switzerland) for discussions and Martine Bourquin and Susana da Costa for help with the rearing of E. ambiguella. Further, we thank Dr. Jean-Luc Perret and Stéphane Donnet (University of Neuchâtel, Switzerland) for the installation and development of the 3D tracking system, Dr. Claudia Lozada-Can and Prof. Dr. Anthony Davison (Swiss Federal Institute of Technology, Lausanne, Switzerland) for their expert help with statistics. Funds for this project were provided by the Swiss Innovation Promotion Agency (CTI Project No. 7273.1 LSPP-LS2) and the National Centre of Competence in Research (NCCR) Plant Survival, a research programme of the Swiss National Science Foundation based at the University of Neuchâtel.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary material 1 (mpeg 12.4 MB)
Rights and permissions
About this article
Cite this article
Schmidt-Büsser, D., von Arx, M. & Guerin, P.M. Host plant volatiles serve to increase the response of male European grape berry moths, Eupoecilia ambiguella, to their sex pheromone. J Comp Physiol A 195, 853–864 (2009). https://doi.org/10.1007/s00359-009-0464-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00359-009-0464-1