
Abstract
We recorded the body axis orientation of free-living black wildebeest relative to incident solar radiation and wind. Observations were made on three consecutive days, on six occasions over the course of 1 year, in a treeless, predominantly cloudless habitat. Frequency of orientation parallel to incident solar radiation increased, and perpendicular to incident solar radiation decreased, as ambient dry-bulb temperature or solar radiation intensity increased, or wind speed decreased. We believe these changes were mediated via their effect on skin temperature. Parallel orientation behavior was more prominent when the wildebeest were standing without feeding than it was when they were feeding. We calculate that a black wildebeest adopting parallel orientation throughout the diurnal period would absorb 30% less radiant heat than the same animal adopting perpendicular orientation. Parallel orientation was reduced at times when water was freely available, possibly reflecting a shift from behavioral to autonomic thermoregulatory mechanisms. The use of orientation behavior by black wildebeest is well developed and forms part of the suite of adaptations that help them to maintain heat balance while living in a shadeless, often hot, environment.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Although many conservators, farmers, and ecotourists will discuss the topic authoritatively, our formal knowledge of behavioral thermoregulatory strategies used by large mammals to avoid or employ directly the energy in solar radiation is based on just a few empirical studies. These studies document shade-seeking and postural adjustments as the principal responses of some species to excess radiant heat load (Ben Shahar and Fairall 1987; Berry et al. 1982; David 1973; Hofmeyr and Louw 1987; Jarman and Jarman 1973; Mitchell 1977), but as far as we are aware no attempt has been made to put thermoregulatory behavior, under solar radiation, into the context of overall behavior patterns. For example, shade-seeking may preclude foraging by grazing animals, which may partly explain the decrease in feeding time observed at high ambient temperature (Lewis 1978). There is therefore, likely, to be a trade-off between thermoregulatory and other behaviors, especially when conflicting demands are placed on the animal’s behavior (Rautenberg et al. 1980; Torres-Contreras and Bozinovic 1997).
An animal reducing the surface area presented to incident solar radiation will reduce radiant heat gain (Clapperton et al. 1965; Riemerschmid 1943). In hot conditions, any reduction in heat gain will reduce the requirement for evaporative cooling. Conversely, in cool conditions an increase in radiant heat gain will reduce the requirement for metabolic heat production. So orientation with respect to the sun can be employed to reduce requirements for water and energy. How birds use solar orientation for thermoregulation is relatively established, heat gain being augmented by wing spreading (Clark and Ohmart 1985; Sellers 1995), but only three studies have reported quantitative data on solar orientation in large mammals. Hofmeyr and Louw (1987) reported that most springbok oriented parallel to incident solar radiation, but under cloud cover, the proportion of the herd orienting parallel to incident solar radiation decreased to no more than expected by chance. Their study, though, was confined to 5 days of observation in one season. Berry et al. (1984) concluded that blue wildebeest oriented parallel to incident solar radiation significantly more than expected by chance, but that the probability of parallel orientation was highest early and late in the day. However, during the warmest period of the day the blue wildebeest were walking to water, and so the orientation would have been toward the water source, independent of the thermal environment. As an incidental observation during a study concerned with territoriality, David (1973) noted that on a single “very hot” day, 250 of 471 bontebok under scrutiny were oriented parallel to incident solar radiation. He also noted that wind direction did not influence orientation.
The black wildebeest (Conochaetes gnou) is endemic to plains in the temperate grasslands (highveld) and arid shrubland (Karoo) of South Africa (Skinner and Smithers 1990). During summer the ambient dry-bulb temperature can approach or exceed mammalian core temperature. In their treeless habitats, black wildebeest have little access to shade, but even if they do have access to shade they typically do not seek it (Estes 1991), despite the combination of hot days, clear skies and high altitude imposing radiant heat loads approaching 1,000 W m−2. Nevertheless, free-ranging black wildebeest exhibit very narrow (<1°C) nychthemeral variation in body temperature during summer (Jessen et al. 1994). They achieve excellent homeothermy in the heat, presumably without incurring large evaporative losses of water because they drink infrequently (personal observations). The strategy that these wildebeest appear to employ to reduce solar heat load is solar orientation (Jessen et al. 1994), but that has not been quantitatively analyzed.
We studied the orientation behavior of free-ranging black wildebeest, at a single habitat, on six separate occasions over the course of 1 year, and addressed several questions. Firstly, do wildebeest orient preferentially with respect to incident solar radiation? Secondly, do other factors in their thermal environment affect solar orientation behavior, and is there a seasonal and/or nychthemeral pattern in solar orientation? Thirdly, do other homeostatic behavior (e.g., feeding) over-ride or influence solar orientation behavior?
Materials and methods
Behavioral observations were made on black wildebeest on Benfontein game farm (111 km2) near Kimberley, South Africa (28°50′S, 24°50′E), approximately 1,200 m above sea level. The climate is semi-arid with dry winters (June to August) and wet summers (December to February). Mean annual rainfall at Kimberley between 1876 and 1994 was 422±132 mm. The vegetation of the area is described as Kalahari thornveld invaded by Karoo (Acocks 1975) and the farm consists of open thorn tree savannah surrounding a large treeless pan or calcareous tufa (approximately 6 km2). Herds of black wildebeest tended to concentrate on and around the pan, which was covered in shrubs and short perennial grasses.
We conducted surveys from sunrise to sunset on three consecutive days in each of May, August, November, and December 1994, and January 1995. Conditions were overcast on the first 3 days of observations in November and 3 further days of data were collected after the weather had cleared. These periods were analyzed separately (see below). Observations commenced before sunrise and continued until after sunset.
A herd was observed from a stationary vehicle parked between 50 and 700 m (typically about 200 m) away, using 10×50 binoculars (Nikon). Data were collected by the scan sample method (Altmann 1974). Every 15 min the behavior and compass orientation of each member of the herd under observation were recorded. Behaviors were recorded in seven categories but reduced to four categories for analysis (lying, standing, feeding, other). How these behaviors changed over the course of the year is discussed in a separate paper (Maloney et al. 2005). Orientation was classified into eight 45° arcs, relative to magnetic north. Data were recorded onto audiocassette and transcribed later. Every 30 min the compass direction of the shadow cast by a gnomon protruding vertically from a horizontal board was used to calculate the compass bearing of incident solar radiation.
The herd under observation usually consisted of a territorial bull with adult females and a few calves (Skinner and Smithers 1990). We analyzed the orientation of the adult females only.
Weather data were collected near to where behavioral observations were made. Sensors measuring dry bulb temperature, relative humidity, solar radiation intensity, wind speed, and wind direction were connected to a data logger (MC Systems, South Africa) which read each sensor every minute and recorded 15-min averages. Weather data for day 2 in August were lost.
Data analysis
For each observation period, the orientation of each animal, according to its compass arc, was expressed relative to the incoming solar radiation; for example, if an animal was facing the sun it was placed in the 0°C arc, if it was facing directly away from the sun in the 180°C arc, etc. Similar calculations were performed to place the animals into eight arcs relative to the prevailing wind.
Since the number of animals observed at each scan was not constant, we calculated summary statistics in two ways. Firstly the number of animals oriented in each arc at each scan was summed for each hour, day and for the entire data set. The proportion of the time spent oriented in each arc then was calculated as the total number in each arc divided by the total observations. The summary statistics calculated in this manner will be biased toward scans made when the herd was large. Secondly, whatever the prevailing herd size, the proportion of the herd in each arc at each scan was calculated and the average proportion of animals oriented in each arc was calculated for each hour, day and for the entire data set. The summary statistics calculated in this second manner will be biased toward scans when the herd was small. The estimated proportion of time spent oriented in each arc by the two methods differed by an average 0.72±1.5% (mean±SD) for calculations covering hours, by 0.77±0.9% for calculations covering days, and 0.77±0.6% for calculations covering seasons. So in spite of the potential distortions, the two approaches to the calculations arrived at essentially the same answer, and we based our further analyses on the first method of calculation.
Meteorological conditions prevailing at each survey were specified by reducing dry-bulb temperature, wind speed, solar radiation, and black globe temperature to hourly averages for each day and analyzing with two-way (survey [factorial] by time [repeated measure]) ANOVA. A Student–Newman–Keuls test was performed for post-hoc comparisons.
On the basis of preliminary analysis of the meteorological conditions, we consolidated our 3-day surveys into “seasons”, the surveys in May and August (Southern Hemisphere autumn and winter) were pooled into a “Cool” season, those in the clear, dry days in November and in December into a “Warm” season, and those made on overcast days in November into a “Summer overcast” season. While most of the measured meteorological conditions were similar between the Warm season and those in January, the behavior in January proved to be quite different. Since the main difference between January and the previous surveys was the occurrence of the first significant rains of the wet season just prior to the January survey, we have analyzed these data separately and refer to the January data as the “Summer post-rain” season.
Statistics
Orientation was analyzed using several methods to address specific hypotheses. Mean angle relative to the sun or wind was calculated using circular statistics (Batschelet 1981). Whether orientations differed significantly from randomness was assessed using Rayleigh’s test (Batschelet 1981). Orientation was noticeably bimodal, so the procedure of doubling angles, which corrects for that bimodality, was used. Correction was made for categorical data since the orientations were categorised into eight distinct compass arcs (Batschelet 1981). Data reflecting orientation to wind were omitted from analyses if wind speed was less than 1 m s−1.
Because of the bimodal nature of the data, we further consolidated the 0° and 180° arcs as orientation parallel to incoming radiation (called “Par” for parallel on figures), the 45, 135, 225, and 315° arcs as orientation oblique to incoming radiation (called “Obl” on figures), and the 90°, and 270° arcs as orientation perpendicular to incoming radiation (called “Prp” for perpendicular on figures). The expected values for random orientation in the oblique category are twice that expected for either the parallel or perpendicular category, and so to simplify comparisons on figures, the values for observed and expected in the oblique category are halved. The proportion of each day spent in each orientation category was used to calculate a mean and standard deviation for overall orientation relative both to incoming radiation and to wind, for all days (n=18), as well as for each season (Cool n=6 days, Summer overcast n=3 days, Warm n=6 days, Summer post-rain n = 3 days). These means were checked for deviation from randomness using a Students t test (random=0.25 proportion for each category). Bonferroni correction for multiple comparisons was applied to account for tests being conducted simultaneously. At a finer time scale, selection by animals of orientation in parallel and perpendicular categories, averaged over each hour, was calculated for each season and compared to that expected if the orientation was random, again with Bonferroni correction for multiple comparisons.
We also examined the relationship between orientation and behavior. For each day (n=18), we calculated the proportion of observations in which the animals engaged in each behavior (Lie, Stand, Feed, Other) fell into each of the orientation categories. From these daily values we calculated a mean and standard deviation for the orientation adopted when the animals were engaged in each behavior over the 18 days. This mean was checked for significant deviation from randomness, again with implementation of the Bonferroni correction to account for multiple comparisons.
Comparisons across seasons and behavioral categories were performed by testing for differences in the frequency distribution of orientation using hierarchical-loglinear analysis for multidimensional contingency tables (SPSS v. 5.01, SPSS Inc., IL, USA). ORIENTATION (parallel, oblique, perpendicular) was the dependent variable, BEHAVIOR (lie, stand, feed, other) and SEASON were main effects, with an interaction term of SEASON × BEHAVIOR. Loglinear analysis essentially compares observed orientation frequencies to those expected based on the interaction between seasons and behavior. We carried out the hierarchical loglinear analysis in two stages. First we used a saturated hilog-linear model to determine the strength of the associations within the main effects and interaction term relative to the ORIENTATION variable. Then any significant main effects and interaction terms were fitted into a logit model with ORIENTATION used as the dependent variable. The logit model that could be interpreted most easily proved to be a saturated logit model (likelihood ratio and Pearson’s chi-square was <0.001, df=0, P=1.0).
Standardised residuals (SR) were calculated for each orientation category within the ORIENTATION variable for each main effect as well as the interaction term as part of the logit process. The SR may be negative or positive depending on the magnitude of the observed frequency relative to the expected frequency (Norusis 1992). A SR greater than 3.27 for an orientation category corresponded to a probability of less than 0.001 that the observed frequency was the same as the expected frequency within the main effect or interaction. Similarly 2.56<SR<3.27 corresponded to a P<0.01; and 1.96<SR<2.56 corresponded to a P<0.05.
Standard linear regression was then used to determine if relationships existed between ambient conditions and orientation. Proportions of animals oriented parallel or perpendicular to incident solar radiation over hours (and days) were arcsine transformed and regressed on hourly (and daily) ambient dry-bulb temperature, solar radiation intensity, wind speed, and black globe temperature. Black globe temperature proved to show the strongest relationships to orientation behavior and so was used in subsequent analyses. The proportion of hours spent oriented parallel to incident solar radiation when the animals were performing different behaviors was also analyzed. Hours where fewer than five animals had a particular behavior were omitted. The proportions of observations in each hour when the animals were lying, standing, or feeding parallel to incident solar radiation were regressed on black globe temperature for that hour. The arcsine transform was performed whenever proportions were analyzed. Because more than one variable was regressed on daily or hourly globe temperature, the Bonferroni correction was applied. When proportion of hours oriented parallel to incident solar radiation proved to be significantly related to black globe temperature while the animals were standing or feeding, we determined whether the regression equations were the same by comparing the slope and intercepts (Zar 1996). Similarly, orientation to the wind was analyzed by regressing the proportion of observations parallel and perpendicular to the wind against wind speed for days and hours. Data where wind speed was less than 1 m s−1 were omitted.
When black globe temperature (which integrates dry bulb temperature, solar radiation intensity, and wind speed) proved to influence orientation (see Results) a forward stepwise multiple regression was performed to determine which components of ambient conditions (dry-bulb temperature, solar radiation intensity, or wind speed) contributed to the influence.
The magnitude of the solar heat load intercepted by wildebeest in parallel and perpendicular orientations was estimated based on previous studies on cattle (Hogan and Skouby 1923; Riemerschmid 1943). Details are given in Appendix.
Results
Eighteen days of data were collected, consisting of scans of the herd every 15 min for 11–12 h day−1. Over the 819 separate scans, the median number of adult females in the herd under observation was 18 and the 25th and 75th percentiles were 10 and 24, respectively. There was one scan where only two animals were visible, and the minimum number of animals observed in any other scan was five. The maximum number of animals observed in any scan was 58.
Weather
Dry-bulb temperature, solar radiation intensity, and black globe temperature differed significantly between the seasons (main effect of season P<10−4 in each case), while wind speed did not differ. Post-hoc comparisons showed that dry-bulb temperature and solar radiation intensity did not differ between the Warm and the Summer post-rain season, but was higher in both these seasons than in the Cool and the Summer overcast seasons (Fig. 1). Mean dry-bulb temperature was significantly higher, and solar radiation significantly lower, in the Summer overcast than in the Cool season. Black globe temperature did not differ between the Cool and the Summer overcast seasons, or between the Warm and the Summer post-rain seasons, but was significantly higher in the latter two seasons than in the former two.

Average hourly meteorological data for the four seasons. For clarity, error bars have been omitted. Average SEM for the cool, overcast, warm, and Summer postrain seasons were, respectively, 2.5, 2.5, 3.8, and 1.3 for dry-bulb temperature, 37, 81, 38, and 38 for solar radiation intensity, 0.7, 1.1, 2.8, and 3.1 for wind speed, and 2.5, 4.7, 1.3, and 2.8 for globe temperature
Orientation
Orientation selection relative to incident solar radiation departed significantly from random (Fig. 2a) with a mean bimodal angle of 2±37°. The proportion of orientation in the parallel category was significantly greater than expected by chance, and proportions in the oblique and perpendicular categories were significantly less than expected by chance. Orientation selection relative to wind also departed significantly from random (Fig. 2b) with a mean bimodal angle of 31±40°. However, the proportion of orientations in the three categories, relative to wind direction, did not differ from that expected for a random orientation, probably because the preferred orientation indicated by the mean angle lay close to a category boundary. On average, the wind was incident at 321±39° relative to the incident solar radiation (Fig. 2c).

a Orientation to incident solar radiation pooled across all surveys. Right panels are radar plots showing relative frequency of observations in each of the eight orientation arcs and the mean bimodal angle (solid dark line). Left panels show proportion of observations (mean±SD for n=18 days) in the three pooled bimodal orientation categories (Par parallel, Obl oblique, Prp perpendicular). Significant deviations from a random distribution of orientations (dotted line) are identified above each bar (*** P<0.001). b Orientation of wildebeest to incident wind pooled across surveys. Detail of plot as for (a). c Wind direction relative to incident solar radiation pooled across surveys. Radar plot shows relative frequency of wind direction in the eight arcs relative to incident solar radiation and the mean bimodal angle for these data (solid dark line)
Orientation to incident solar radiation departed from random in each of the Cool, Warm and Summer post-rain seasons (Fig. 3; mean bimodal angles of 17±39, 355±32, and 18±38°, respectively), but was random in the Summer overcast season. When orientations were pooled into the three broader categories, those in the Warm season departed significantly from the expected random proportions (Fig. 3c), in that orientation in the parallel category occurred significantly more often, and in the oblique and perpendicular categories less often, than expected by chance. Comparisons across seasons showed that parallel orientation occurred significantly more in the Warm season than in the other seasons, and less in the Cool and the Summer overcast season than in the Summer post-rain season. Orientation in the perpendicular direction occurred more during the Cool and the Summer overcast seasons than the other seasons, and less during the Warm season than during the Summer post-rain season.

Orientation relative to incident solar radiation for each season. Right panels show radar plots with relative frequency of observations in eight arcs and the mean bimodal angle (solid dark line) if the distribution differed significantly from random. Left panels show proportion of observations (mean±SD for n=6 days for Cool, n=3 days for Summer overcast, n=6 days for Warm, and n=3 days for Summer post-rain) in the three bimodal orientation categories (Par parallel, Obl oblique, Prp perpendicular). Proportion expected if orientation was random (0.25) is shown as broken horizontal line. Significant deviations from a random distribution of orientation (dotted line) are identified above each bar (***P<0.001). Results of LogLinear analysis for comparison across seasons are shown within each bar (+ significantly higher than expected, - significantly less than expected), number of symbols indicates significance levels + P<0.05, ++ P<0.01, +++ P<0.001). For example +++ on Warm/parallel indicates that the proportion of animals observed oriented in the parallel category during the Warm season was highly significantly greater (P<0.001) than expected
Orientation adopted when the animals were engaged in each of the four behaviors departed significantly from random (Fig. 4, Lie 21±39°, P<0.001; Stand 1±35°, P<0.001; Feed 356±38°, P<0.001; Other 354±38°, P<0.001). However, once again, the proportion of orientations in the three reduced categories, when the animals were exhibiting Lie and Other behaviors, did not differ from the proportions expected by chance. When the wildebeest were Standing they oriented significantly more in the parallel direction, and less in the oblique and perpendicular directions, than expected by chance. When the wildebeest were Feeding they oriented significantly more in the parallel direction, and less in the perpendicular direction, than expected by chance. Comparisons across behaviors showed that wildebeest adopted the parallel orientation more when Standing than during any other behavior, and less when Lying than during the other behaviors. Orientation in the perpendicular direction occurred less when Standing and Feeding than when Lying or performing the Other behaviors.

Orientation relative to incident solar radiation for each behavioral category, pooled across seasons. Details of plots as for Fig. 3. The distribution differed significantly from random in all four cases (n=18 days for each behavior)
Selected orientation varied not only with season and behavior, but also with the time of day (Fig. 5). In the Cool season, orientation parallel to incident solar radiation was significantly more frequent, and perpendicular orientation was significantly less frequent, than expected by chance at 1300 and 1400 (Fig. 5a), corresponding to the time of day when solar radiation was highest (Fig. 1). During the Summer overcast season, orientation did not deviate from random orientation at any time (Fig. 5b). In the Warm season, orientation perpendicular to incident solar radiation was avoided for most of the day, and orientation parallel to incident solar radiation was significantly more frequent than expected by chance from 0900 to 1100 and again from 1500 till sunset (Fig. 5c). Orientation parallel to incident solar radiation was no different from that expected by chance from 1200 to 1400 (Fig. 5c). In the Summer post-rain season, orientation perpendicular to incident solar radiation was avoided occasionally, but orientation parallel to incident solar radiation occurred more frequently than expected for 6 of the 12 h (Fig. 5d). Contrary to the general trend for that season, at 1100, orientation parallel to incident solar radiation occurred less frequently, and perpendicular to incident solar radiation occurred more frequently, than expected by chance.

Proportions of animals, in each hour, oriented parallel (circles) or perpendicular (squares) to incident solar radiation for each season. Proportion expected if the animals oriented randomly (0.25) is identified as a horizontal broken line on each figure. Probability of observed proportions being random is shown by symbols above or below the mean for each hour (*P<10−1, **P<10−6, ***P<10−9)
The proportion of the day spent oriented parallel to incident solar radiation increased, and the proportion of the day spent oriented perpendicular to incident solar radiation decreased, as average black globe temperature increased (Fig. 6a; parallel F 1,15=13.4, P=0.002, r 2=0.47; perpendicular F 1,15=7.2, P=0.02, r 2=0.33). Similarly the proportion of each hour oriented parallel to incident solar radiation increased significantly, and perpendicular orientation decreased significantly, as average hourly black globe temperature increased (Fig. 6b; parallel F 1,198=20.9, P<10−4, r 2=0.1; perpendicular F 1,198=17.7, P<10−4, r 2=0.08).

Relationship between average daily or hourly black globe temperature and a the proportion of the day spent oriented parallel (circles) or perpendicular (squares) to incident solar radiation, b the proportion of each hour spent oriented parallel (circles) or perpendicular (squares) to incident solar radiation, and c the proportion of each hour spent parallel to incident solar radiation when engaged in the behaviors of lying (open circle), standing (+) or feeding (filled circle). Least squares regression lines in (a) and (b) are solid for parallel and broken for perpendicular. Least squares regression lines in (c) are for standing (thin) and feeding (thick); the regression for lying was not significant and is not shown. All data were arcsine transformed for analysis. Random orientation (0.25) translates to 0.52 and is indicated by horizontal broken line. Proportion of 0.7 translates to arcsine of 1.0
The proportion of each hour spent oriented parallel while standing or feeding also increased as average hourly black globe temperature increased, while there was a tendency for the proportion spent oriented parallel while lying to increase (Fig. 6c; Lie F 1.148=4.8, P=0.03, r 2=0.033; Stand F 1,190=10.9 P=0.001, r 2=0.05; Feed F 1,177=11.6 P=8×10−4, r 2=0.06). The regression lines for standing and feeding did not differ in slope (t 363=0.75, P=0.45) or intercept (t 364=1.8, P=0.7).
Surprisingly, there was no effect of wind speed on orientation to the wind. The only circumstances in which wind appeared to influence behavior was when the wind carried rain. At these times (during the Summer overcast season) the wildebeest oriented facing away from the wind and rain. However, when integrated with other variables, wind speed did prove to influence orientation with respect to the sun (see below).
Forward stepwise multiple regressions aimed at identifying which component of the thermal environment influenced orientation and behavior were similar for average day and hour data, so we only present data for hours. Orientation parallel to incident solar radiation was influenced by all three meteorological variables, namely dry-bulb temperature (T db), solar radiation (RAD) and wind speed (WS). The multiple regression equation (F 3,198=6.3, P=4×10−4, multiple r 2=0.03) was:
The standard errors of the estimates were 0.0025 for T db, 0.00007 for RAD, and 0.004 for WS. The positive coefficients for T db and RAD show that orientation parallel to incident solar radiation increased as T db and RAD increased. The negative coefficient for WS shows that orientation parallel to incident solar radiation decreased as wind speed increased. The Beta weights (a measure of the relative contribution of each independent variable to the dependent variable) for the three independent variables were T db=0.2, RAD=0.15, WS=−0.1.
Discussion
We have shown that, during the day, black wildebeest in open habitat oriented parallel to incident solar radiation significantly more than expected by chance, and so the answer to our first question (raised in Introduction) is affirmative. Parallel orientation was more likely, and perpendicular orientation less likely, as ambient dry-bulb temperature or solar radiation intensity increased, or wind speed decreased, and so we also arrive at an answer to our second question affirmatively: physical factors did affect solar orientation behavior. There were differences in the use of orientation behavior at different times of the year, but these differences appear to reflect changes in the thermal conditions rather than an intrinsic seasonal change in behavior, except perhaps after rain. The probability of adopting parallel orientation was different when the animals were lying, standing or feeding, suggesting an affirmative response to our third question: solar orientation did differ between different behaviors. However, the selection of orientation when engaged in different behaviors can be explained partly by the time of day, and therefore by the thermal stress, at which the different behaviors predominated, as well as by an intrinsic difference in orientation with behavior (see below).
A central question is whether the orientations that we observed were driven by the influence of the sun or the wind. An animal standing and facing south with the sun from the north and the wind from the east may be choosing to orient its rear to the sun or side to the wind, or some combination of the two. We found that orientation was associated significantly overall with both the sun and wind direction, with the mean angle for solar orientation being 2° and for wind orientation being 31°. Since the solar angle corresponded to selection for minimisation of surface area, but the wind angle corresponded to no apparent logical strategy, we calculated the overall wind direction relative to the sun. This angle was 321° (Fig. 2c), meaning that for wind speed above 1 m s−1 the wind generally was coming from 321° (−39°) relative to the sun. If an animal was oriented parallel to incident solar radiation, with the wind incident from 321° relative to the sun, then we would have placed its orientation at 39° relative to the wind, which is close to the mean wind orientation of 31°. Since wind orientation was not influenced by wind speed, and there was no other aspect of the behavior except orientation to incident solar radiation influenced by wind speed, we conclude that the significant orientation to wind (Fig. 2b) resulted from the animals orienting to incident solar radiation and there being a climatological bias in the wind direction relative to the incident solar radiation direction.
Our conclusion that black wildebeest had no preferred orientation to wind is in contrast to the findings of Berry et al. (1984) for blue wildebeest, who concluded that wind direction did influence body orientation. Wind speed in their study was much higher than during our study, and there also a possible bias was in their data since the blue wildebeest walked to water during the day, and therefore adopted an orientation toward the water source, which was not taken into account in the analysis.
Springbok cohabit with black wildebeest at our study site and elsewhere. Hofmeyr and Louw (1987) previously showed that preference of springbok for orientation parallel to incident solar radiation was highest in full sunshine and decreased under cloud cover, indicating that transduction of incident solar intensity probably mediated the response. We have provided evidence for a similar phenomenon in black wildebeest, since the probability that an animal adopted orientation parallel to incident solar radiation increased as solar radiation intensity increased. Orientation also was influenced by ambient dry-bulb temperature. In fact, in a multiple regression analysis, dry-bulb temperature had a greater influence, as reflected in higher Beta weight, than did solar radiation, as a predictor of parallel orientation. While these factors contributed to the probability of parallel orientation, the low r 2 for the relationships suggests that there are other factors influencing individual orientation choice.
The fact that incident solar radiation intensity and dry-bulb temperature contributed independently to orientation behavior helps us to interpret some of the differences in orientation behavior between seasons. Parallel orientation was most common in the Warm season, when both dry-bulb temperature and solar radiation intensity were highest. During the Cool season, dry-bulb temperatures remained relatively low and parallel orientation was not predominant overall (Fig. 3a). However, between 1300 and 1400 there was selection for parallel orientation (Fig. 5). At this time incident solar radiation intensity already had peaked and was declining, but dry-bulb temperature was just approaching its maximum (Fig. 1). During the Summer overcast period, however, incident solar radiation intensity would appear to have been the main cue for orientation, because dry-bulb temperature was higher than during the Cool season but incident solar radiation intensity was lower (Fig. 1), and there never was selection for parallel orientation (Fig. 3b). We believe that the interplay between dry-bulb temperature and radiation intensity occurs because the wildebeest is actually responding to overall heat balance (see below), a proposition supported by our observation that the probability of parallel or perpendicular orientation correlated better with black globe temperature, an index of overall heat load, then either dry-bulb temperature or incident solar radiation intensity separately.
In the laboratory, thermoregulatory behavioral responses can be induced by altering brain temperature (Refinetti and Carlisle 1986). In humans, behavioral thermoregulation is driven approximately equally by peripheral and core temperature (Frank et al. 1999). In our black wildebeest we think that skin temperature must have been providing the main input to the brain areas responsible for the thermoregulatory behavior. Skin temperature will be increased by elevations in both dry-bulb temperature and solar radiation intensity, while elevated wind speed, at dry-bulb temperature below body temperature, will increase convective heat loss and so reduce skin temperature. Deep body temperature of black wildebeest is tightly regulated (Jessen et al., 1994), and temperature changes were more from an endogenous rhythm than from changes in environmental conditions leaving core temperature changes as an unlikely modulator of orientation. Our results imply that there is a threshold heat load for orientation responses, because they were absent in the overcast season and reduced in the cool season, but that above that threshold, the influence of dry-bulb temperature and incident solar radiation intensity combined to influence the behavior in the Warm season.
We found that preferred orientation differed with different behaviors, with parallel orientation being much more common when the animals were standing than when feeding or lying. The wildebeest also tended to stand more, and avoided feeding, at hotter times of the day (Maloney et al. 2005), so the preference for orientation parallel to solar radiation when standing likely reflects the use of parallel orientation more in hotter conditions rather than increased use for orientation when standing. It is also possible that a feeding animal may be motivated by food choice to move in a certain direction, so over-riding orientation motivated by thermal factors. But the relationship between the probability of parallel orientation and globe temperature was the same for feeding and standing, suggesting that the former is the more likely explanation. We conclude that differences in preferred orientation during different behaviors partly reflect the priority given to thermoregulation during that behavior, and partly are the passive consequence of reduced active behaviors at hotter times of the day when orientation was more likely.
When the wildebeest were lying, their orientation departed significantly from random, with a mean angle close to parallel to solar radiation, but the preference for parallel orientation was weaker than observed during standing or feeding. This relaxation of orientation occurred despite the finding that lying tended to be the predominant behavior on hot afternoons (Maloney et al. 2005). By lying down a wildebeest, presents its entire side to the sky and it is likely that orientation makes little difference to the surface area intercepting incident solar radiation, but this hypothesis requires confirmation. When the solar angle exceeds 70° (i.e. the sun is close to overhead) the surface area presented to incident solar radiation becomes essentially independent of orientation (Riemerschmid 1943). This may explain our observation that parallel orientation was preferred in the morning and afternoon of the Warm surveys, but not around midday (Fig. 5). In November and December the sun zenithed at >80° and was above 70° angle from 1200 to 1400.
Very similar meteorological conditions were experienced in the Summer post-rain period as during the Warm season, but the preference for parallel orientation was significantly lower in the post-rain period. In general, across surveys, the diurnal time spent feeding decreased as the days became hotter, but during the Summer post-rain period the wildebeest spent more than half of the day feeding (Maloney et al. 2005). In the 8 months before that period, only 40 mm of rain was recorded at the study site but an additional 75 mm fell in the week preceding the observations. We hypothesize that the availability of free water allowed the wildebeest to rely more on evaporative cooling, and so reduce the need for thermoregulatory behaviors such as orientation. In support of that hypothesis, we frequently noted panting after the rain, but never in the Warm surveys, even though the black globe temperature was virtually identical. In the Summer post-rain survey, wildebeest did feed more and stand less, and therefore had a lower probability of parallel orientation, but that effect was not powerful enough to account for the 25% reduction in parallel orientation compared to the Warm surveys. We believe that the main difference between the equally warm pre-rain and postrain periods was the switch from behavioral heat avoidance to evaporative cooling.
We quantitatively estimated the effect of changing orientation on heat balance of wildebeest (see Appendix), as has been done previously for cattle and sheep. Riemerschmid (1943) calculated that, whatever the solar angle, absorption of direct solar radiation always was less for cattle oriented parallel to incident solar radiation than for those oriented perpendicular. The reduction in solar heat load by parallel orientation was 30% at 10° solar elevation and decreased to only 2% at 80°. These differences were important, because total solar radiation absorbed represented three times the heat generated by metabolism. Similarly Clapperton et al. (1965) calculated for sheep that parallel orientation led to a 50% reduction in absorbed radiation compared to perpendicular orientation, at 10° solar elevation.
During the Warm season wildebeest oriented parallel to incident solar radiation experienced a solar heat load only 70% of that if they oriented perpendicularly (Fig. 7). The relative difference was greater in the Cool season because the sun had a shallower zenith. How that difference in heat load impacts on wildebeest thermoregulation will depend on ambient conditions. In the Cool season, it might be possible for the animal to dissipate the entire solar heat load non-evaporatively (Clapperton et al. 1965; Finch 1972). In the Warm season, ambient dry-bulb temperatures were over 30°C after 1000 hours and so the capacity for non-evaporative heat loss was limited. We calculate that to evaporate the entire direct solar heat load, wildebeest oriented perpendicular all day would require 6.9 litres of water (assuming 2,400 J g−1 of water evaporated) while parallel orientation would require 4.8 l, a difference of 2.1 l. If water turnover rate in black wildebeest is similar, per kg, to that measured in the zebu cow, eland, goat, or awassi sheep (King 1979) then a 120 kg wildebeest will turn over about 14 l day−1, and the savings accrued by using parallel orientation would be 15% of the daily water budget. However, if water turnover is more like that of the arid-adapted oryx (King 1979), then total turnover would be 7.4 l day−1 and the use of parallel orientation throughout the day would save 30% of the daily water turnover.

Energy absorbed over 12 h from direct solar radiation for a black wildebeest, calculated for each season, for animals at a fixed orientation. Filled bars are for perpendicular orientation for the entire 12 h, open bars represent parallel orientation. Details of the calculations used to obtain these totals are given in Appendix
Our study has placed the use of orientation behavior by a large mammal species onto a quantitative footing, using over 15,000 observations of black wildebeest collected at different seasons and at all daylight hours. Circular statistics revealed that black wildebeest do use orientation behavior to avoid solar heat loads. Orientation was influenced by the time of day and time of year and appeared to be mediated by prevailing climatic conditions. Orientation parallel to incident solar radiation increased as dry-bulb temperature increased, solar radiation intensity increased, or wind speed decreased. We believe that these changes were mediated via their effect on skin temperature which acts as a signal to the brain to control orientation behavior. Orientation also differed depending on the prevailing behavior and was probably influenced by the availability of water for thermoregulation, in that the proportion of the time spent orienting parallel was reduced when water was freely available. We conclude that the use of orientation behavior by black wildebeest is well developed and forms part of the suite of adaptations that help them to maintain heat balance while living in an often hot, shadeless environment.
References
Acocks JPH (1975) Veld types of South Africa. Mem Bot Surv S Afr 40, 128 pp
Altmann J (1974) Observational study of behaviour: sampling methods. Behavior 49:227–267
Batschelet E (1981) Circular statistics in biology. Academic, London
Ben Shahar R, Fairall N (1987) Comparison of the diurnal activity patterns of blue wildebeest and red hartebeest. S Afr J Wildl Res 17:49–54
Berry HH, Siegfried WR, Crowe TM (1982) Activity patterns in a population of free-ranging wildebeest Connochaetes taurinus at Etosha National Park. Z Tierpsychol 59:229–246
Berry HH, Siegfried WR, Crowe TM (1984) Orientation of wildebeest in relation to sun angle and wind direction. Madoqua 13:297–301
Clapperton JL, Joyce JP, Blaxter KL (1965) Estimates of the contribution of solar radiation to the thermal exchanges of sheep at a latitude of 55° north. J Agric Sci 64:37–49
Clark RG, Ohmart RD (1985) Spread-winged posture of Turkey Vultures: single or multiple function? Condor 87:350–355
David JHM (1973) The behaviour of the bontebok, Damaliscus dorcas dorcas, (Palls 1766), with special reference to territorial behaviour. Z Tierpsychol 33:38–107
Estes RD (1991) The behaviour guide to african mammals. University of California Press Ltd, Oxford, England
Finch VA (1972) Energy exchanges with the environment of two East african antelopes, the eland and the hartebeest. Symp Zool Soc Lond 31:315–326
Frank SM, Raja SN, Bulcao CF, Goldstein DS (1999) Relative contribution of core and cutaneous temperatures to thermal comfort and autonomic responses in humans. J Appl Physiol 86:1588–1593
Hofmeyr MD, Louw GN (1987) Thermoregulation, pelage conductance and renal function in the desert-adapted springbok, Antidorcas marsupialis. J Arid Environ 13:137–151
Hogan AG, Skouby CI (1923) Determination of the surface area of cattle and swine. J Agric Res 25:419–430
Jarman MV, Jarman PJ (1973) Daily activity of impala. East Afr Wildl J 11:75–92
Jessen C, Laburn HP, Knight MH, Kuhnen G, Goelst K, Mitchell D (1994) Blood and brain temperatures of free-ranging black wildebeest in their natural environment. Am J Physiol Regul Integr Comp Physiol 36:R1528–R1536
King JM (1979) Game domestication for animal production in Kenya: field studies of the body-water turnover of game and livestock. J Agric Sci 93:71–79
Lewis JG (1978) Game domestication for animal production in Kenya: shade behaviour and factors affecting the herding of eland, oryx, buffalo and zebu cattle. J Agric Sci 90:587–595
Maloney SK, Dawson TJ (1995) The heat load from solar radiation on a large, diurnally active bird, the emu (Dromaius novaehollandiae). J Therm Biol 20:381–387
Maloney SK, Moss G, Cartmell T, Mitchell D (2005) Alteration in diel activity patterns as a thermoregulatory strategy in black wildebeest (Connochaetes gnou). J Comp Physiol A (in press)
Mitchell A (1977) Preliminary observations on the daytime activity patterns of lesser kudu in Tsavo National Park, Kenya. East Afr Wildl J 15:199–206
Norusis MJ (1992) SPSS for Windows Based System User’s Guide Release 5. SPSS Inc, USA
Rautenberg W, May B, Arabin G (1980) Behavioral and autonomic temperature regulation in competition with food intake and water balance of pigeons. Pflugers Arch Eur J Physiol 384:253–260
Refinetti R, Carlisle HJ (1986) Effects of anterior and posterior hypothalamic temperature changes on thermoregulation in the rat. Physiol Behav 36:1099–1104
Riemerschmid G (1943) Some aspects of solar radiation in its relation to cattle in South Africa and Europe. Onderstepoort J vet Res 18:327–353
Sellers RM (1995) Wing-spreading behaviour of the Cormorant Phalacrocorax carbo. Ardea 83:27–36
Skinner JD, Smithers RHN (1990) The mammals of the southern african subregion. University of Pretoria Press, Pretoria
Torres-Contreras H, Bozinovic F (1997) Food selection in an herbivorous rodent: Balancing nutrition with thermoregulation. Ecology 78:2230–2237
Zar JH (1996) Biostatistical analysis, 3rd edn. Prentice Hall Inc, New Jersey
Acknowledgements
We thank Dr. Mark Berry and DeBeers Consolidated Mines for permission to carry out the study on Benfontein, and Peter, Jennifer, Gregory, and Nicky Gibbs for their friendship and help on site. Tammy Cartmell, Steven Cartmell and Simone Glassom helped with data collection. Petro Vorster from the Kimberley office of the South African Weather Bureau kindly supplied the rainfall data. Professor Phil Withers and an anonymous reviewer helped to improve the manuscript. The study was funded in part by the South African Foundation for Research Development. SKM was in receipt of a University of the Witwatersrand Post Doctoral Fellowship. These experiments comply with the “Principles of animal care” publication number 86–23, revised 1985 of the National Institute of Health, and also the laws of South Africa.
Author information
Authors and Affiliations
Corresponding author
Appendix
Appendix
The heat absorbed from direct solar radiation was calculated for black wildebeest oriented parallel and perpendicular to incident solar radiation for each hour and then summed for the diurnal period for each survey. Heat absorbed was calculated as the product of surface area presented × measured incident solar radiation intensity × absorption coefficient.
-
1.
Solar radiation intensity incident on a horizontal surface was measured at the field site (Fig. 1).
Fig. 8 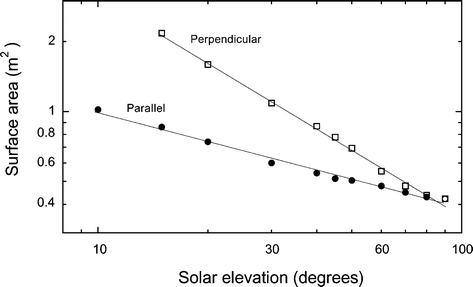
The calculated surface area projected on to a horizontal surface, presented to incident solar radiation by a 130 kg black wildebeest at solar elevation angles between 10° and 90° when oriented perpendicular (squares) or parallel (circles) to incident solar radiation. See Appendix for details of calculations. The best fit for the relationship proved to be a power relationship, as indicated by the linear fit on a log–log plot
-
2.
The absorption coefficient was assumed be 80%, the same as that measured for dark brown cattle (Riemerschmid 1943). The heat load from radiation in dark coats is influenced by wind speed, coat conductance, and the penetrance of radiation into the pelage (Maloney and Dawson 1995). Ignoring these factors would result in an overestimate of the heat load, and so the figures we arrived at would represent the maximal heat load of the orientations. However, the proportional difference between orientations would be independent of these errors.
-
3.
The fraction of the total body surface area presented to incident solar radiation varies with body orientation and solar elevation. Solar elevation was calculated for each hour during each survey for the latitude and longitude of our study site using software available at http://www.susdesign.com/sunangle. The surface area presented at different solar elevations was calculated based on data from Riemerschmid (1943) obtained for an 800 kg bull (total surface area 6.5 m2) oriented either parallel or perpendicular to the solar beam. The mass of an average adult female black wildebeest is 130 kg (Skinner and Smithers 1990). The total surface area of a cow weighing 130 kg is 1.86 m2 (Hogan and Skouby 1923). We assumed a similar geometry for wildebeest and cattle, and so the surface areas presented for cattle was scaled by 1.86/6.5. The best fit was a power equation (Fig. 8). For perpendicular orientation SA (m2)=26.9×solar elevation−0.98 and for parallel orientation SA (m2)=2.54×solar elevation−0.41, with solar elevation in degrees. For each hour (0700 to 1800) during each survey, the surface area presented to incident solar radiation was calculated from these equations for the appropriate solar elevation.
-
4.
Solar radiation absorbed was then calculated as the surface area (m2) × the measured incident solar radiation intensity (W/m2)×0.8. For each day the total absorbed solar energy was calculated by summing the hourly values for over 12 h. These values represent the extremes of the range of orientations, because they assume a fixed orientation for the full day.

Rights and permissions
About this article
Cite this article
Maloney, S.K., Moss, G. & Mitchell, D. Orientation to solar radiation in black wildebeest (Connochaetes gnou). J Comp Physiol A 191, 1065–1077 (2005). https://doi.org/10.1007/s00359-005-0031-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00359-005-0031-3