
Abstract
We presented free-flying locusts (Locusta migratoria L.) with sounds that varied in temporal structure and carrier frequency as they flew toward a light source in a flight room under controlled temperature and light conditions. Previous studies have shown tethered locusts react more often to trains of 30-kHz pulses than to pulse trains below 10 kHz. Further, this acoustic startle response has been suggested to function in bat-avoidance. We expected free-flying locusts to respond similarly; however, we found locusts responded to all sounds we presented, not just high-frequency, “bat-like” sounds. Response rates of turns, loops, and dives varied from 6% to 26% but were statistically independent of carrier frequency and/or pulse structure. Free-flying moths and tethered locusts were tested using a subset of our acoustic stimuli under the same temperature and light conditions as the free-flying locusts. Moth responses were carrier frequency dependent as were responses of tethered locusts positioned along the flight path observed in our free-flight trials. All responses were unaffected by a 90% reduction in room light. We conclude that locusts possess an acoustic startle response evocable in free flight, however, free-flying locusts do not show the same discrimination observed in tethered locusts under similar conditions.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Insect ears mediate both intraspecific communication, such as finding and evaluating a mate or signalling a rival to move away, and interspecific communication, such as predator detection and avoidance. In locusts, there is evidence the abdominal, tympanate ear may function in both roles. Males in copula frequently stridulate when disturbed by another male and high-frequency, bat-like sounds have been shown to evoke avoidance responses in tethered, flying locusts. If a locust is on the ground, sounds may evoke different behaviours than if a locust is in flight (or indeed if in tethered flight), therefore, the effect of a particular sound on the ongoing behaviour of the locust presumably depends on the context with which the locust is engaged at the time of receiving the stimulus. Further, other factors such as ambient light levels (day versus night), or the presence of other locusts, may also influence the behavioural repertoire available to the animal in response to acoustic stimulation. In this paper, we show that locusts can react to sounds while in free flight and that the response rates are not entirely consistent with a predator-avoidance function.
Tympanate ears protect many nocturnal, flying insects from predation by insectivorous bats (for reviews see Hoy 1992; Fullard and Yack 1993; Hoy and Robert 1996). Typically, an insect detecting an approaching bat will react with an acoustic startle/escape response (ASR) which results in movement away from the predator (Hoy 1989, 1992; Hoy et al. 1989) or in rendering the animal less conspicuous to the predator (e.g. cessation of singing; Faure and Hoy 2000). The most extensively studied insect ASR is the bat-avoidance behaviour of tympanate moths. Moths detecting a distant bat will turn away from the sound, whereas moths detecting a close bat will respond with loops, dives and increases in flight path erraticism (Roeder 1962, 1964, 1967, 1974). Similar last-ditch responses are also seen in free-flying praying mantises (Dictyoptera) (Yager et al. 1990) and lacewings (Neuroptera) (Miller and Olesen 1979). Synthesized bat-like sounds are also able to evoke similar behaviours in a group of nocturnal butterflies, the Hedyloidea, which respond to intense ultrasound when in flight with steep dives, climbs, loops, and turns (Yack and Fullard 2000). Thus, for several diverse orders of free-flying tympanate insects, the reactions to the echolocation calls of bats seem to show convergence on behaviours consisting of turns, loops, and active and passive dives during flight.
Observing freely moving, unrestrained animals is experimentally ideal (Kutsch 1999); however, collecting data from nocturnally active insects interacting with flying bats in their natural environment poses certain technical difficulties that are not easily overcome. This fact, coupled with the obvious benefit of restraining an animal for recording behavioural elements with transducers and presenting controlled acoustic stimuli, has resulted in many biologists opting to work with tethered flying insects stimulated with synthesized bat-like sounds. How tethering affects an insect is not well understood but it is agreed that the effects are either direct, by influencing the degrees of freedom of movement by mechanically interfering with the body at the tethering location, or indirect by removing sensory input normally present when the animal is engaged in the same behaviour but is unrestrained.
When tethered, field crickets (Gryllidae) and katydids (Tettigoniidae) respond to bat-like sounds with an ASR that consists of abdomen and hindleg deflection, head rolling, flexion of the antennae, wing twisting (inferred from basalar muscle activity), and changes in wing beat frequency and flight cessation (Moiseff et al. 1978; Pollack and Hoy 1981; Moiseff and Hoy 1983; Pollack et al. 1984; Nolan and Hoy 1986; Libersat and Hoy 1991; and see reviews by Doherty and Hoy 1985; Pollack and Hoy 1989; Hoy 1989, 1992; Hoy et al. 1989). Other tethered flying insects react to bat-like sounds in a similar way. Examples include the praying mantis, Parasphendale agrionina (Yager and May 1990) and the tiger beetle, Cicindela marutha, which responds to bat-like sounds with head rolling, leg extension, and changes in wing kinematics (Yager and Spangler 1997).
The African migratory locust (Locusta migratoria L.), an acridid grasshopper, also possesses an ASR with similar characteristics to those described above and by association with crickets and katydids, the ASR has been hypothesized to function in bat avoidance (Robert 1989). Tethered flying locusts react with latencies of 50–100 ms to high-frequency, bat-like sounds consisting of trains of short-duration sound pulses with carrier frequencies greater than 10 kHz presented at intensities greater than 45 dB sound pressure level (SPL). These reactions consist of abdomen deflection and dorsiflexion, hindleg extension and deflection, changes in wing beat frequency, including flight cessation, changes in wing stroke kinematics and other postural adjustments (Hoy et al. 1989; Robert 1989; Robert and Rowell 1992; Dawson et al. 1997). Robert (1989) showed that abdomen and hindleg deflection in response to stimulation with bat-like sounds was accompanied by the production of steering torques in directions opposite to the side of stimulation. Further, Dawson et al. (1997) using a 10-ms, 30-kHz stimulus pulsed at 50 Hz showed that bat-like sounds produce asymmetric wing depression coincident with abdominal deflection and dorsiflection and suggested that free-flying locusts would react with banked, downward turns.
The objective of this paper was to observe how locusts in free-flight react to sounds and to determine if their reactions were specifically bat-avoidance reactions. We presented all combinations of stimuli that varied among three different carrier frequencies (5 kHz, 12 kHz, and 30 kHz) and three different pulse structures (stridulation-like, intermediate, and bat-like). Since flight tendencies in L. migratoria are affected by light intensity (Uvarov 1977) we also tested animals under light levels comparable to daylight (full light, >100 lx) and dusk (reduced light, <100 lx). We chose the carrier frequencies and temporal patterns of the pulses based on sounds that might have significance in the natural behaviour of locusts. For example, recordings of wing-beat noise show spectral peaks between 3.5 and 5 kHz (Haskell 1957) and stridulation sounds show peaks at 12 kHz (Kalmring 1975).
Materials and methods
Animals
Adult male and female L. migratoria L. aged 15–20 days post-imaginal moult were used in these experiments. All animals were reared in a crowded colony maintained at the Universität Konstanz at 36°C (day) and 26°C (night) with 75% relative humidity under a 12 h:12 h light:dark cycle. Only locusts in apparent excellent condition were used in this study. Moths were collected using hand nets from mercury-vapour lamps illuminating paths bordering a deciduous woodland area and unused agricultural fields near U. Konstanz (called Hockgraben: located 47° 41’ 18” N, 9° 11’ 27” E ) between 2230 and 0000 hours during July and August 1999. Moths were kept in a screen cage (approx. 30 cm×20 cm×30 cm) at 4°C to prevent self inflicted damage to their wings until tested (at most two days) in the flight room. To ensure correct moth identification, we prepared a reference collection from the area and only tested those that could be positively identified. Species were identified using Forster and Wohlfahrt (1960, 1971, 1981) and collection records cited in Bauer (1993).
Flight room
The flight room measured 8 m long, 5.7 m wide and 2.2 m tall. The floor (grey cement) and walls (cement painted white) were marked with a grid, in 0.5-m increments in red and black tape, which we used to define a co-ordinate system for referring to the location of locusts and equipment in the room (Fig. 1). Temperature was maintained at 32±3°C (65–75% relative humidity) with four 2,000-W forced-air heaters; two located at the front of the room and two at the back of the room. The heaters were directed away from the centre of the room to prevent thermal eddies in the flight path of the locusts.

The flight room. A Photograph and B diagram of the room showing location of equipment and the flight path (dashed line) of locusts past the speakers. The operational area of the flight room measures 8 m long by 5.7 m wide by 2.2 m tall and was separated from equipment in the rest of the room by two large heavy cloth curtain partitions (heavy lines). To facilitate observations, the room is marked in 1-m intervals along the floor and 0.5-m intervals along the walls which define the co-ordinate system used in this study (X=width, Y=length, Z=height). The room was illuminated by a photographers studio lamp located at (2, 7, 1.1), fluorescent tube fixtures on the wall, one above the photographers lamp located at (3, 7, 2.1), and one on the opposite wall at (-2.7, 4.5, 2.1), and a single 60-W incandescent light bulb at (0, 1.5, 2.1). Note that the fluorescent tube fixture on the ceiling in the centre of the room in the photograph (A) was not illuminated during experiments. The room was heated by four 2,000-W forced air heaters (rectangles), two at the front of the room and two at the rear (arrows indicate the direction of air flow away from the centre of the room). Locusts were launched by dropping them into flight from a height of approx. 2 m at co-ordinates (0, 1, 2). As locusts fell they opened their wings and then flew forward (dashed line), under their own power, toward the photographers lamp in the corner of the flight room past two speakers located at (2.5, 5.6, 1.15 and 2.5, 6, 1.15); direction of speakers indicated by arrows. Sounds were presented when the locust flew past the 4-m mark. The reactions of the locusts were observed by two people. W.K. sat at the left side of the room while J.W.D. launched the locusts into flight and made observations from the rear of the flight room
A d.c.-powered photographic studio lamp (Profilux, model 200, with a metal halide bulb, Osram, model HMI) was positioned in one corner of the flight room (co-ordinates 2,7 at a height of 1.1 m). Additional lighting was provided by fluorescent tube fixtures and a single 60-W incandescent light bulb located 0.5 m in front of the locust launch point. For observations under reduced light intensity, we placed 90% attenuation neutral-density filters (Lee Filters, no. 211) on all the lights except the incandescent bulb. The distribution of light in the room was measured at 1-m intervals along the length and width of the entire flight room using a Panlux electronic light meter fixed to a tripod at a height of 1 m. The light meter was rotated 360° at each measurement location and the direction, read from a protractor fixed to the tripod, and magnitude of maximum intensity, was recorded. For all locations in the room, at both illumination levels, maximum light intensity was in the direction of the photographers studio lamp in the corner of the flight room (Fig. 2).

Calibration of the sound field and light intensity in the flight room. A, B, C Isointensity plots of broadcast sounds [in dB sound pressure level (SPL)], at speaker height, in relation to the location of the speakers in the flight room for the three carrier frequencies used in experiments. For all frequencies, sound intensity in the room near the point in the flight path where the locusts are stimulated (1, 4 to 1,5; see Fig. 1), was approx. 75–78 dB SPL. The complex distribution of intensities was a result of using two speakers 40 cm apart broadcasting in offset directions (see Fig. 1) and the acoustics of the flight room (cement walls and floor with a particle board ceiling). Background noise in the flight room was typically between 61 and 64 dB SPL. See methods for other measurements of the acoustics of the flight room. D, E Isointensity plots of light intensity measurements (lx) for the flight room in reduced and full light. Light intensity was uniformly decreased by placing neutral density film (90% attenuation) over all light fixtures with the exception of the incandescent bulb near the launch position. The direction of maximum light intensity at all measurement points (1-m intervals along the length and width of the room) was in the direction of the photographers lamp in the corner of the flight room
Acoustic stimuli
An arbitrary waveform generator (PC Instruments, model PCI-311) controlled by a microcomputer with software supplied by the manufacturer (BenchTop Lite, v. 3.3 J) generated all waveforms used as acoustic stimuli. Three envelopes were used in experiments. The first envelope was based on the stridulation sounds produced by L. migratoria as described by Kalmring (1975). This pattern consisted of pairs of shaped pulses each with a 6-ms rise time, 9-ms sustain time, and 2-ms fall time. The pairs of pulses (measuring a total of 34 ms) were presented with an inter-pulse-interval of 42 ms producing a stimulus period of 76 ms (13.2 pulses s−1). We presented ten pairs of pulses as a train for a total stimulus time of 718 ms. The second pattern was taken from Robert (1989). This pattern consists of single pulses with a 1-ms rise/fall time and 18 ms sustain time. These pulses are presented with an 80-ms inter-pulse-interval yielding a stimulus period of 100 ms (10 pulses s−1). We presented trains of ten pulses for a total stimulus time of 920 ms. Our last envelope, designed to resemble the temporal pattern of an approaching/tracking echolocating bat (Simmons et al. 1979; Surlykke and Moss 2000), consists of pulses with a 1-ms rise/fall time and 8 ms sustain time presented with a 20-ms inter-pulse-interval producing a period of 30 ms (33.3 pulses s−1). We presented trains of ten pulses for a total stimulus time of 280 ms. Each of the three envelopes was synthesised using sine waves with carrier frequencies of 5, 12, and 30 kHz. For each locust in free flight, we presented all combinations of carrier frequency and envelope (i.e. nine stimuli). Some of these combinations are arguably nonsensical (e.g. a bat-like sound envelope presented at 5 kHz), but were included to determine if locusts would show reactions.
A pair of two-inch cone tweeters (Motorola, model KSN1078A) were positioned on the right side of the flight room (Fig. 1). The speakers were 1.15 m above the floor, 0.5 m from the right wall and 0.4 m apart from each other. Two speakers were necessary to create a distributed sound field given the size of the room and the directional properties of the speakers. The sound field in the flight room for the carrier frequencies of 5, 12, and 30 kHz was measured at three different heights (0.5 m, 1.15 m (speaker height), and 1.5 m) (Fig. 2). Sounds were measured at 0.5-m intervals along a rectangular region of the flight room in front of the speakers (the flight zone). SPLs of pure (sinusoidal) tones with carrier frequencies and amplitudes equivalent to the stimulus pulses (Stapells et al. 1982) were measured with a Brüel and Kjaer 1/2-inch microphone (type 4133) coupled to a Brüel and Kjaer measuring amplifier (type 2606). The 1/2-inch microphone has a flat response to 40 kHz.
To verify the quality of the acoustic stimuli in the flight room, we examined recordings of the stimuli broadcast from the speakers. The broadcast sounds were recorded at speaker height (1.15 m) from the centre of the “flight zone” at co-ordinates (1,5) with a Brüel and Kjaer 1/2-inch microphone (configuration as above) and separately with a Brüel and Kjaer 1/4-inch microphone (type 4135) with a Brüel and Kjaer preamplifier (type 2618). The 1/4-inch microphone has a flat response to 100 kHz. Both microphones were coupled to the measuring amplifier described above with the 22.4-Hz high-pass filter engaged. Signals were recorded onto 7-inch reels of magnetic tape (BASF HiFi DP26) using an instrumentation tape recorder (RACAL, model Store-4) using a direct board with the tape travelling at 30 inches s−1. Signals on tape were replayed without slowing and digitised (Axon Instruments Digidata, model 1200B using Axoscope v. 7.0) at 333.3-kHz sampling rate. Spectral analysis of the sounds was performed with ScopeDSP v. 3.6a (Iowegian International).
The size of the room, the composition of the walls and the fact that we broadcast from two speakers simultaneously, resulted in a complex sound field in the locust flight zone (Fig. 2A–C). The SPL of the 30-kHz stimuli attenuated over shorter distances than the 12- and 5-kHz stimuli as expected and we adjusted the amplitude of the waveforms input to the speakers so that the SPL of the stimulus pulses at the 4.5-m mark (the point along the locusts flight path at the moment of stimulation) was between 75 dB and 78 dB for all stimulus frequencies. Sound pressures at speaker height (1.15 m) relative to measurements 0.5 m above the floor or 0.7 m below the ceiling did not differ by more than 3 dB. Recordings of the stimuli showed that some echo was present in 5-kHz stimuli but it was not sufficient to significantly distort the envelope of the stimuli. Spectral analysis of the recorded stimulus pulses showed that they were narrowly tuned at their broadcast frequency and that the stimulus pulses were the dominant sound in the room. We also confirmed that the heaters, lights, and other (electrical) equipment in the room did not produce ultrasound that would affect our experiments.
Observations of locusts in free flight
Locusts were assigned to individually numbered clear plastic containers (15 cm deep with a 10-cm2 mouth) kept under a heat lamp at the rear of the flight room. The rear area of the flight room is out of the sound field of the speakers and was separated from the observation area by heavy black cotton curtain partitions.
All locusts were launched from the same position at the centre of one end of the flight room (co-ordinates 0,1). Locust flight was initiated by holding the locust horizontally at a height of 2 m (±5 cm) and releasing it (Kutsch and Fuchs 2000). The locust was held between the thumb and first and second fingers, dorsum up, by pinching the metathoracic tibia-femur joint and folded wings. The descent of the locust induced wing opening and locusts normally began forward flight at a distance of between 1 m and 0.3 m from the floor. No forward velocity was intentionally added to the locust during launch. Each locust was launched into flight and allowed to fly within the flight room at least three times before formal observations began. During formal observations, the nine stimulus patterns were presented in random order and we cycled through the locusts at each stimulus pattern, light intensity, and sound intensity regime. We consider each flight of each locust to be independent as an individual locust was never tested under the same conditions more than once (except for controls, see below).
Locusts typically flew from the launch position toward the photographic studio lamp in the corner of the flight room (dashed line in Fig. 1B). All flights were observed by two people (W.K. and J.W.D.) in the flight room at different vantage points. J.W.D. launched and observed the locusts as they flew the length of the room while W.K. made observations while sitting along the left wall of the room (at approximate co-ordinates −2, 2). Locusts were stimulated with sound as they approached the speakers. Stimuli were triggered manually (by J.W.D.) when locusts were within 2.5 m (i.e. on the right side of the room) of the speaker and at least 20 cm above the floor and 50 cm below the ceiling. Locusts that did not fly through this “flight zone” were immediately flown again. If after five flights the locust did not enter the flight zone, they were omitted from the observations for that experiment. From the calibrations of the sound field, sound intensity was typically between 75 and 78 dB (Fig. 2A–C) at the location of stimulus onset.
We made detailed notes about the in-flight reaction of each locust as it flew past the speakers toward the lamp. Details such as whether or not a reaction was observed, changes in flight speed, direction, altitude, and the location of the reaction relative to the speakers (for SPL estimates) were noted. We also scored the landing positions of locusts on the walls, floor and ceiling of the flight room on a three dimensional diagram of the flight room. For the purpose of scoring the landing position, some locusts flew on a straight flight path then, within 0.5 m of the wall (or lamp) would veer into a new direction which we presume was to avoid collision. Where locusts veered away from the walls and lamps, we scored the location before veering (i.e. the landing/collision site extrapolated from the flight path immediately before veering). Locusts did not exhibit this veering activity in the centre of the flight room in the absence of acoustic stimuli (i.e. we did not mistake auditory evoked responses for collision avoidance responses).
To be sure we were not mistaking coincident erratic manoeuvres with reactions to sounds, we observed control flights in which locusts were launched into flight but were not presented with sound. Animals that flew in such a way that we could not distinguish reactions to sounds from their flight tendencies were discarded from the analysis. Similarly, after all data was collected, we discarded all data from animals that could not be tested for three or more trials of the nine sound pattern-frequency combinations.
Observations of moths in free flight
Moths were tested under identical room conditions as the locusts. After moths were identified, they were separated into individual, labelled containers (the same used for holding the locusts, see above). All moth flights were observed from two different vantage points in the flight room by two individuals. It was not possible to launch a moth into flight the same way we launched a locust, therefore, we held the container in a way that the moth could climb out and enter flight without touching it. For some individuals, a gentle flick of the container was necessary to coax it from the rim. When moths were within the “flight zone” (see above), they were presented with sound. We tested moths with the bat-like sound pattern with carrier frequencies of 5, 12, and 30 kHz in both full light and reduced light. Notes describing the flight path before and after stimulation indicating any reactions to sound, including the direction of turns relative to the speaker positions, were recorded in a manner identical to the locust flight trials. Control flights were observed in which no sounds were presented to the moths.
Observations of locusts tethered in the flight room
All physical aspects of the flight room were identical to those during free-flight tests. A 1.15-m-tall retort stand was positioned in the flight room at co-ordinates (1,5), the “far” location or (2,6) the “near” location relative to the speakers. The “far” tether was 1.7 m from the speakers and the “near” tether was 0.5 m from the speakers. Sound intensity, depending on frequency, was between 78 dB and 80 dB at the far location and between 82 dB and 90 dB at the near location. When tethered, the locusts faced the photographers lamp in an alignment consistent with the observed flight path of free-flying locusts.
At least 1 h before testing, a copper saddle was fixed to the pronotum of each locust with melted bees-wax. A desk fan (Braun, model HL1) was placed 25 cm in front of the tether and produced a wind stream of 2.5 m s−1 (measured with a Lambrecht Type 641 bN anemometer). We recorded the sounds produced by the fan (recording and analysis method as above for stimulus pulses) at the same distance as the tethered locusts. Spectral analysis showed that most of the sound energy was below 10 kHz and the SPL of the noise produced by the fan was 60 dB (at 25 cm).
During testing, locusts were retrieved from their individual containers, affixed to the tether and were stimulated with sound. When necessary, locusts were prodded into flight by touching their abdomen or head or by suddenly removing tarsal contact with a finger momentarily presented to the locust. Sounds were presented only after locusts had assumed a tucked flight posture (forelegs flexed and tucked behind the head-capsule, hindleg tibia flexed and held next to femur, abdomen held straight or with slight dorsiflexion of the tip) which occurred after 10–30 s of flight. Tethered locusts were tested with the bat-like sound pattern with carrier frequencies of 5, 12 and 30 kHz. Locusts were tested at both locations in full light and reduced light.
Tethered locusts were videotaped at 25 frames s−1 (fps) from behind using a Panasonic MS1 SVHS video camera with a 1:1.4 TV zoom lens shuttered at 1/250th of a second. For analysis, videotapes were converted from European PAL format (25 fps) to North American NTSC format (30 fps) and were reviewed on a Panasonic editing VCR (model AG7300) with single-frame shuttle job capability and a Sony Trinitron video monitor (model PVM1341). During review of the videotape, detailed notes were made of any reactions to the stimulus. These notes included abdomen deflections, abdomen dorsiflections, hindleg extensions, changes in wing-beat frequency, wing kinematics and flight cessation.
Three-dimensional re-construction of locust flight paths
We used two cameras [a Panasonic MS1 SVHS video camera, model NV-MS1 HQ and a Bauer (Bosch) SVHS video camera, model VCC550 AF] positioned at equal heights such that their video fields overlapped at 90°. One camera was positioned at co-ordinates (1,1.8), while the other camera was placed at co-ordinates (-2.2,5). Both cameras had a 1:1.4 TV zoom lens shuttered at 1/250th of a second. As locusts flew through the overlapping video fields, the relative position of the images of the locust in the two cameras was used to reconstruct the location of the locust relative to the co-ordinate system of the flight room (see below). Additional cameras were used to monitor stimulus presentation which provided a method for synchronizing the two videotapes. The cameras and stimulus monitoring equipment used for 3-D reconstruction of flight path data were not present during the free-flight tests (described above).
For reconstruction of the flight paths the videotapes were converted from PAL (25 fps) to NTSC (30 fps) format and reviewed on a Panasonic AG7300 editing VCR. The horizontal and vertical location of the locust relative to the flight room walls and floor (i.e. its projected location on the walls) was recorded for each video frame. For each camera, the distance from the camera lens to the wall in view was known and we could calculate the angle, relative to the optical axis of each camera, of a line from the camera lens to the projected locust location on the flight room wall. The actual location of the locust is the point where these two lines intersect. Because the cameras were at the same height, we could find this point relative to one of the cameras by solving a system of three equations in two unknowns. Finally, because the location of that camera in the flight room was known, we could find the locust location relative to the room co-ordinates.
Statistical treatment of the data
All statistical tests assumed significance with P≤0.05. Unless otherwise indicated, N represents the number of individual locusts observed. Contingency table G-tests and P values were calculated using procedures in Zar (1984). Two-by-two contingency table G-tests and associated William’s corrected G values were calculated with Windows 2×2 Tables (v.2.0 by Kelvin F. Conrad).
Results
Observations of locusts in free-flight
Locusts had little difficulty manoeuvring in our flight room. Under bright light, free-flying locusts were observed circling the lamps and speakers, landing on the walls, floor, and ceiling and jumping into flight without apparent difficulty. We spent 3 weeks systematically trying different lighting conditions, speaker locations and stimulus parameters in the flight room. From this period of preliminary observation we found that locusts would not fly or enter into sustained flight from our launch technique in the dark or under very low light (less than approximately 2 lx). We also found that locusts strongly orient to sources of light in the flight room. We used this to our advantage and found that by varying the amount of ambient light in the room, and the location of the lights, we could launch locusts at one end of the flight room and they would fly on a direct flight path, past our speakers, to the photographers studio lamp positioned in the corner of the room (Fig. 1, dashed line). Under the same conditions, repeated flights of the same locust yielded consistent flight paths and landing positions. After this period of trial-and-error we were satisfied that locusts responded to sounds while in free-flight, that our equipment was arranged in a manner sufficient for eliciting responses, and that we were making observations from good vantage points in the flight room.
We saw a diversity of behaviours in response to sounds including left and right turns, upward and downward turns, loops, spirals, ditches/drops to the ground and zigzagging (wobbling) in the flight path. Unlike tethered flight, locusts in free-flight are not confined to a single plane of movement and left and right turns were often combined with changes in elevation producing, for example, an upward left turn or a downward right turn. We categorized responses according to the most pronounced component observed. For example, a large left turn with slight upward motion was categorized as a left turn. Downward turns, when the locust did not contact or land on the ground, and ditches/drops to the ground were combined into the same category. Upward turns, due to the relatively low ceiling in the flight room (2.2 m), often yielded collisions with the ceiling. Loops and spirals (two or more loops in succession) were frequently observed and consisted of locusts following a cork-screw type flight path with an initial upward component and being completed without the locust contacting the ground. We observed both large and small diameter cylinders in the spirals. Zigzag and wobbling flight paths were observed when the locusts repeatedly changed either elevation or azimuth in an up-down-up-down-up-down or a left-right-left-right-left-right fashion respectively.
Three dimensional re-constructions of the flight paths of four locusts responding to sound in the flight room are shown in Fig. 3, and are typical reactions to sound such as loops (Fig. 3A), turns away from the speaker (Fig. 3B, C), and drops to the ground (Fig. 3D). Figure 3C shows a clear increase in flight speed away from the speaker as the turn is completed. We frequently observed changes in flight speed as part of the reactions to sound. Also, many locusts would remain motionless for several seconds on the ground or walls, apparently frozen, after certain reactions, e.g. ditches to the ground.

Three-dimensional reconstructions of segments of the flight paths of four different locusts responding to sounds in the flight room. A bat-like sound with 30-kHz carrier frequency elicited a loop/spiral (A) and a turn away from the speakers (B). The locust in C was stimulated when flying close to the speakers and responded with a turn and a clear increase in flight speed away from the source of the sounds. Bat-like sounds at 12 kHz elicited a quick drop to the ground in front of the lamp (D). Arrows indicate the time of stimulus onset and the asterisk indicates the beginning of the reconstructed sequence. The length of the flight path segments are defined by the overlapping field of views of the two cameras used to film the responses (see Materials and methods). Sequences were typically 16–19 frames or approx. 532–633 ms total time. Time between points is 33.3 ms
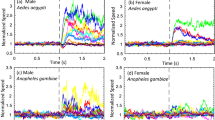
The frequency of the different manoeuvres (e.g. left turn, right turn, loop) varied between 0 and 11% in response to the different sound patterns with reactions to sounds occurring between 6% and 26% of flights (Fig. 4). The type of manoeuvre observed did not depend on the stimulus given in either full light (Fig. 4A) or reduced light (Fig. 4B; full light: contingency table G-test, G=40.404, df=40, P=0.452; reduced light: G=24.859, df=40, P=0.971). Since the speakers were positioned on the right side of the flight path, we expected more left turns (i.e. away from the speaker) than right turns. Of all the manoeuvres that were turns, all except one were left turns in flights in full light, and all were left turns in flights in reduced light (all sound patterns combined, responses tested against an expected 1:1 ratio of left:right turns, full light: Χ2 c goodness-of-fit test, Χ2=11.529, df=1, P=0.0007; reduced light: Χ2 c=5.143, df=1, P=0.023).

The distribution of locust responses to the nine combinations of sound patterns and frequencies in A full light and B reduced light. Sound patterns and frequencies were chosen to establish a gradient from presumably non-aversive (stridulatory-like, 5 kHz) to aversive (bat-like, 30 kHz) stimuli. Locusts responded to all sound patterns with steering manoeuvres and the distribution of manoeuvres observed were not contingent on the stimulus presented. Response rates were low to all stimuli. Left turns (i.e. turns away from the speaker) were more frequent than right turns. Locusts were observed by two people in the flight room and the reactions of the locusts were categorised as turns (left or right), changes in altitude (up or down), loops (sometimes spiralling flight) or other. The proportion of locusts showing any of the indicated responses (R) and no responses (NR) is summed at the right of each histogram. N represents number of individuals observed
When all manoeuvres were collapsed into response or no response, response rate was not contingent on the sound pattern presented. In other words, no one sound pattern elicited more responses than any other sound pattern (Fig. 5A; full light: contingency table G-test, G=7.895, df=8, P=0.444; reduced light: G=11.091, df=8, P=0.197). When responses to each sound pattern were considered individually, the number of locusts responding did not depend on the light intensity under which they were tested except for the intermediate sound pattern presented at 12 kHz (contingency table G-test, G c=4.270, df=1, P=0.039; all other patterns, P=0.116 to P=0.959). When data were collapsed into the three sound patterns, the number of locusts responding to each pattern was not contingent on light intensity (stridulatory-like: contingency table G-test, G c=2.889, df=1, P=0.089; intermediate: G c=2.829, df=1, P=0.093; bat-like: G c=0.332, df=1, P=0.565). Similarly, when data were collapsed into the three carrier frequencies of the sound patterns, the number of locusts responding at each frequency was not contingent on light intensity except at 5 kHz (5 kHz: contingency table G-test, G c=4.087, df=1, P=0.043; 12 kHz: G c=1.154, df=1, P=0.283; 30 kHz: G c=0.033, df=1, P=0.856; Fig. 5B).

The proportion of locusts responding in full light and reduced light was not contingent on the sound pattern presented (A) or the carrier frequency of the sound pattern (B)
In summary, locusts in free-flight reacted to sounds while they flew past the speakers toward the photographers studio lamp in the corner of the flight room. The frequency of responses did not depend on the temporal pattern of the stimulus or on carrier frequency. A variety of reactions were observed including turns, loops and spirals but the type of reaction also did not depend on the temporal pattern of the stimulus or carrier frequency. Responses were also independent of light intensity. These findings are in contrast to predictions, from previous studies using tethered locusts, that locusts in free-flight would react to high-frequency bat-like sounds more frequently than low-frequency sounds. To verify that our results were not artefacts of conditions in the flight room, we examined responses of wild-caught moths and tethered locusts tested in the same flight room under the same conditions as the free-flight tests.
Observations of moths in free-flight
We tested a total of 74 individuals derived from three superfamilies and seven families (Table 1). The majority of moths tested did not fly in a straight flight path from the point of launch to the lamp in the corner of the flight room. There was considerable variation both within and between moth species in their flight tendencies. Typically, moths flew with intermittent bouts of fast and slow flight sequences with frequent turns and changes in elevation. Notable examples of this type of flight behaviour come from Ochropleura plecta (Noctuidae) and Ochrostigma melagona (Notodontidae). However, other species such as Craniophora ligustri (Noctuidae), Diacrisia sannio (Arctiidae), and Phoesia tremula (Notodontidae) flew with slower (flight speeds estimated to be less than 1 ms−1), mostly straight, smooth flights from the point of release to the walls or lights (not necessarily the photographers lamp). Because the moths did not fly in a straight path from launch to the light past the speakers, we did not distinguish between left and right turns but note that all turns were away from the speakers.
The proportion of moths responding to bat-like sounds at 5, 12, and 30 kHz with the different manoeuvres above are presented in Fig. 6A. The manoeuvres with which the moths responded (e.g. turn, loop) were not contingent on the sound presented in either full light or reduced light (full light: contingency table G-test, G=9.6751, df=8, P=0.289; reduced light: G=5.4258, df=8, P=0.711). More moths responded to 30-kHz bat-like sounds than to 5- or 12-kHz sounds in both full light and reduced light (full light: contingency table G-test, G=62.3654, df=2, P<<0.0001; reduced light: G=108.4610, df=2, P<<0.0001) (Fig. 6B). The number of moths reacting to bat-like sounds at each of the three frequencies was not contingent on light intensity (5 kHz, contingency table G-test, G c=0.037, df=1, P=0.848; 12 kHz, G c=0.112, df=1, P=0.738; 30 kHz, G c=3.190, df=1, P=0.074).

The reaction of moths to bat-like sounds at 5, 12, and 30 kHz in the flight room. The type of reaction (turns, changes in elevation, loops) observed in response to the stimulus did not depend on the frequency of the sound (A), but moths responded more often to 30-kHz sounds than to 5- or 12-kHz sounds (B). The number of moths responding in full light was not different than the number responding in reduced light
Observations of locusts tethered in the flight room
Locusts were tethered at speaker height at two locations in the flight room; a location near to the speaker (co-ordinates 2,6) and one farther from the speaker (co-ordinates 1,5). At both locations, locusts responded to sound with: flight cessation, deflection of the abdomen and hindlegs either toward or away from the speaker, extending a hindleg by opening the femoral-tibial leg joint, dorsiflection of the abdomen, skipping a stroke in the normal wing beat cycle, and increasing wing beat frequency. From our videotapes, we carefully noted the presence of these and other clear responses to the onset of the sound stimulus. Typically responses occurred within three wingbeats (approximately 150–200 ms) of the stimulus and behaviours observed outside of this window were arbitrarily deemed not to be a reaction to the sound. Similarly, locusts that were exhibiting aspects of the above behaviours before the stimulus was applied were discarded from analysis. Our detailed notes revealed no observable differences in responses to sound between tethered locusts in this study and other studies with which we have participated. Since the above responses to bat-like sounds of tethered locusts have been amply documented in the literature (Robert 1989; Robert and Rowell 1992; Dawson et al. 1997), we simply counted the number of locusts responding and not responding from our detailed notes.
The proportion of locusts responding to bat-like sounds with carrier frequencies of 5, 12 and 30 kHz, in full light and reduced light, at the two tether locations, are presented in Fig. 7. Sound intensity was greater at the near location than at the far location for all frequencies tested (12 dB louder at 30 kHz, 2 dB louder at 12 kHz, and 7 dB louder at 5 kHz). Overall, more locusts responded to high-frequency than to low frequency stimuli and responses were more frequent to 12 and 30 kHz stimuli at the near tether than at the far tether. In full light, the proportion of locusts responding was contingent upon the carrier frequency of the bat-like sounds with more locusts responding to 30 kHz (near: contingency table G-test, G=18.096, df=2, P=0.0001; far: G=12.359, df=2, P=0.002); the same trend is present in reduced light, but is not statistically significant (near: contingency table G-test, G=4.985, df=2, P=0.082; far: G=5.255, df=2, P=0.072) most likely because fewer locusts were tested in reduced light. When each frequency tested is considered independently, the number of locusts responding in full light and reduced light are not different (contingency table G-test, P values range 0.294–0.961).

Tethered locusts responded to bat-like sounds with flight cessation, abdomen deflection and dorsiflexion, deflections or extensions of the legs and changes in the wing beat including increases in wing beat frequency. The number of locusts responding was contingent on the frequency of the bat-like sounds with more locusts responding to the high frequencies at both near and far (relative to the speaker) tether locations. More locusts responded at the near tether than to the far tether. Response rates were not different in full light and reduced light. Sample sizes for locusts at the near tether are N=10, 23 and 8 for full light at 5, 12, and 30 kHz, respectively; N=12, 13 and 11 for reduced light at 5, 12, and 30 kHz. Sample sizes for locusts at the far tether are N=25, 24 and 24 for full light at 5, 12 and 30 kHz, respectively; N=12, 13 and 10 for reduced light at 5, 12, and 30 kHz, respectively
Discussion
The objective of this study was to determine how locusts react to sounds in free-flight. From previously published observations of tethered locusts responding to bat-like sounds (Robert 1989; Dawson et al. 1997), including behavioural audiograms (Robert 1989), free-flying locusts were predicted to produce carrier frequency and pulse-structure dependent responses. Further, if tethering does not adversely affect locust reactions to sound, response rates of free-flying and tethered locusts should not differ to the same stimuli.
Observations of locusts in free-flight
We found free-flying locusts responded to sounds with a diversity of manoeuvres consisting of turns, loops and dives. All turns we observed, except one in full light, were away from our speaker position; an observation consistent with the idea locusts were producing a negative phonotactic response. In one sense, the hypothesis that the ASR functions in predator (bat) avoidance (Robert 1989; Robert and Rowell 1992) is supported by the types of manoeuvres we observed and the fact that locusts were reluctant to fly under reduced light conditions. However, we found responses were independent of stimulus carrier frequency and pulse structure and that response rates were low (between 13% and 26%) relative to tethered locusts (between 50 and 80%) and free-flying moths (between 75% and 85%). An ASR functioning in bat-avoidance does not require it to be elicited exclusively with high-frequency, bat-like sounds, a fact particularly important given that locusts, unlike moths, are not tone-deaf. What is important, however, is that the low response rates relative to moths, an insect that unquestionably uses it ear for bat defence, cause us to question the selective advantage of the locust ASR for bat avoidance. Our response rates suggest that less than one in four locusts will attempt evasion compared with more than eight of ten moths attempting evasion. This is true despite selective pressures on moths and locusts may be different. We feel the most parsimonious explanation of our data, excluding experimenter error, is locusts either possess alternate defences from bats and/or that locusts possess an ASR that is a general startle response evocable by a diversity of sounds, not just biologically relevant sounds.
We have considered a variety of reasons that could account for the discrepancy in response rates between free-flying and tethered locusts. We do not believe that low free-flight response rates are due to observer error, poor stimuli, artefact or other technical problems in the flight room for several reasons. All flight tests, including controls, were observed by two people from different vantage points and both observers had to agree on what was seen before it was tallied. Disagreements resulted in locusts being re-flown or flights being discarded from the data set. This reduced our sample size while retaining locusts that showed definite reactions; a factor that increased the percentage of responding locusts tallied. Our estimates of response rate are, therefore, arguably generous. Further, we are confident that the quality of the stimuli presented, which were spectrally verified, were sufficient to elicit responses from free-flying locusts because we saw steering responses from free-flying moths and locusts tethered at two different locations in the sound field.
We are also confident the intensity of stimuli were sufficient for eliciting responses. The maximum intensity our speakers could broadcast was 98 dB at 5 kHz and 84 dB at 12 kHz (measured at the location locusts were stimulated). Trials at these maximum intensities (data not shown) produced higher response rates (between 12% and 48%) for all the frequencies and sound patterns tested but response rates were still statistically independent of carrier frequency and sound pattern tested.
Another factor contributing to higher response rates, in general, of tethered locusts is that it is easier to observe responses from a tethered animal than from one in free-flight. We videotaped the tethered locusts and scored responses after watching the tapes, sometimes using a frame-by-frame shuttle jog. Using this technology it is possible to see very subtle movements of the locust in response to the stimuli. This may partially account for why response rates for tethered locusts at 30 kHz are higher than for free-flying locusts but does not account for why they are the same at 5 kHz and 12 kHz. While observing the tapes we made detailed notes of all behaviours (postural adjustments) that occurred in response to sound. If we were able to see reactions in tethered locusts but not in free-flying locusts, it suggests that some of the postural adjustments are either not elicited in free-flight or do not have behavioural consequences (i.e. aerodynamic significance) during free-flight. We are not suggesting that abdomen or hindleg extension is not important aerodynamically to a free-flying locust, but simply that we scored all movements in response to sound, including those of small magnitude. Further, for a postural or wing kinematic change to be adaptive in the context of predator avoidance in free-flight, it should result in an observable change in flight path.
We feel the two most plausible explanations for our data are that either a stationary speaker emitting a pattern of calls mimicking an approaching or searching bat, at constant intensity, may not be an adequate simulation of an insect-bat encounter, or, that the ASR was affected differently in free-flying and tethered locusts.
Inspection of strobe photographs of moths (Roeder 1962) and lacewings (Miller and Oleson 1979) interacting with bats show that the movements of the predator and prey through three dimensional space are complex. The intensity of the echolocation calls received by an insect pursued by a bat are more variable than those received by locusts (and moths) in our flight room. A similar situation exists when tethering an insect at a fixed distance to a speaker. Tethering ensures the intensity of the sounds received by the insect are constant during a stimulus and thus response rates of tethered locusts (and other insects) may be generous.
Robert (1989) showed that pure tone stimuli (mimicking a hunting bat) and two-tone stimuli, consisting of a 30-kHz tone presented concurrently with a 5-kHz tone 10–15 dB more intense than the first, have different behavioural effects with the former eliciting negative phonotaxis and the latter suppressing it in L. migratoria. Robert suggested that the biological significance of the difference in behavioural effects would be to suppress an ASR that would otherwise be elicited by high-frequency components in the sounds produced by beating wings. We agree with this interpretation, however, we do not believe it is the reason for our low response rates to 30-kHz stimuli. Measurements from Schistocerca gregaria show that wingbeat noise is composed primarily of frequencies between 3.2 kHz and 5 kHz and has an intensity between 62 dB and 71 dB (Haskell 1957). Locusts flying in the sound field (Fig. 2A, B, C) in the flight room would always be receiving stimuli that are, at a minimum, 5 dB greater than the intensities of wing beat noise and therefore we believe suppression of the ASR via this mechanism is not a likely explanation.
We found locusts orient toward sources of bright light (relative to background light intensity) and we used this to establish a consistent flight path past our stationary speakers. This was necessary because it was not possible to evoke consistent flight paths in locusts without the use of the photographers lamp. Our light intensity calibrations confirm the brightest point in the room was the photographers lamp and the direction of maximum light intensity was always in the direction of the lamp. Svensson and Rydell (1998) showed the ASR of geometrid winter moths can be suppressed by mercury vapour street lamps. They showed winter moth response rates to ultrasound (26 kHz, 110 dB SPL) within 4 m of the lamp were suppressed by 43%. Moths in this study, flying within 5 m of the photographers lamp, responded to our sounds in 75–85% of trials; a figure different than 100% suggesting a decrease in responsiveness consistent with Svensson and Rydell’s (1998) observation. It is not possible to directly compare the responsiveness of locusts and moths to ultrasound, but moths and tethered locusts in the flight room under the same light conditions, stimulated with the same acoustic patterns, responded more frequently than free flying locusts. Tethered locusts tested facing the photographers lamp produced higher response rates to 30-kHz bat-like sounds than free-flying locusts. Therefore, although we can not conclude the photographers lamp did not influence the free-flying locusts, we are confident it does not entirely explain our results. We feel our results are better explained as a combination of the effects of tethering and possible influences of the bright photographers lamp.
Observations of moths in free-flight
Our motivation for testing moths in the flight room was to use them as a “biological control” to show that it was possible to elicit avoidance reactions from a free-flying insect in the conditions of our flight room using our stimuli. There is now ample evidence in the literature that bats are the predominant selection pressure shaping the ASR of moths. Moths tested in the flight room, unlike free-flying locusts, showed high response rates (75–85%) to high-frequency (30 kHz) bat-like sounds and few reactions to low-frequency (5 kHz) bat-like sounds. Moths did not have difficulty flying in our flight room under either light intensity and our observations are consistent with both field and flight room observations by others. Roeder (1962) observed 87.5% of wild moths responding to broadcast bat-like sounds. The responses were 49% dives, 30% turns and 8.5% loops. Similarly, Rydell et al. (1997) observed behavioural reactions of geometrid winter moths (Agriopis spp., Erannis spp. and Alsophilia aescularia) to 26-kHz pure tones presented in pulses controlled by hand (5–10 pulses over 1–2 s) in the field and in a flight room. In the field, 96% of moths responded to bat-like sounds with zigzags (at distances greater than 5 m), spiralling and diving flight (at distances less than 5 m), and in a flight room, 100% of trials showed responses.
Unlike locusts, the frequency dependant nature of moth responses can be explained by the tuning characteristics of the moth auditory organ. For the families tested (Table 1), the sensitivity of the ears, as revealed by audiograms of similar species from extracellular tympanic nerve recordings, is greatest for frequencies from 25 kHz to 70 kHz (Fullard 1988; Surlykke and Filskov 1997; Fullard and Dawson 1999).
We observed moths responding with turns, loops, dives, and a number of other manoeuvres that were clear responses to the stimulus (grouped as ‘other’ in Fig. 4A). These manoeuvres are similar to those observed by Roeder (1962) who noted a variety of responses to bat-like sounds (70-kHz pulse trains) broadcast from a speaker atop a tall pole in the field. He described some responses as sharp dives, where the moth kept its wings motionless during descent, as power dives, where the moth flew toward the ground at a rapid rate, and passive dives, where the descent was interrupted by wing movements. Responses also consisted of deviations from a horizontal flight path (turns), as well as series of loops and tight turns. Roeder admitted having difficulty categorizing the behaviours (as we did for locusts and moths) because in many instances one type of response would be followed by another (e.g. a dive preceded by a tight climb, loop or turn). Roeder also noted responses involved 10–20% increases in wing beat frequency and latencies between 0.2 and 1 s. When stimulation resulted in moths landing, Roeder noted they often remained motionless for several seconds; an observation we also made of many free-flying locusts immediately after a response was observed.
Effects of light intensity
Ambient light intensity may be a cue signalling potential threat from bats, therefore, we tested locusts and moths under full and reduced light to determine if animals flying in light conditions comparable to dusk (less than 100 lx) might be more sensitive to high-frequency sounds. We used gregarious phase locusts in this study and reports of locusts flying at night are typically in the solitary phase (Kennedy 1956; numerous citations in Uvarov 1977; see also Farrow 1990; Gatehouse and Zhang 1995). To our knowledge differences in auditory sensitivity and processing between gregarious and solitary L. migratoria are yet to be determined. Flight in gregarious phase L. migratoria can be stimulated or inhibited by abrupt changes in light intensity (Uvarov 1977). We did not find a systematic effect of light intensity on the reactions of the locust to sounds except that locusts were more reluctant to fly under our reduced light conditions (but note smaller sample sizes in Fig. 4B). When ambient light levels were less than approximately 2 lx, we had difficulty launching locusts into flight wherein they would immediately land on the ground and remain still. This is consistent with the notion that migratory locusts may avoid predation by bats by not flying at night.
Tethering effects and auditory sensitivity
Tethering affecting the auditory system of locusts is one plausible explanation for why we observed tethered locusts reacting more frequently than free-flying locusts to high frequency sounds. During tethered flight, movement of the wings and mechanical deformations of the thorax produce phasic and tonic activity in the auditory nerve (N6) that arises from stimulation of the receptors at the tympanum (Hedwig 1988). This activity results in the auditory system being approximately 20 dB less sensitive to certain (low frequency) sounds during flight relative to when an animal is stationary. It is therefore possible that tethering artifactually accentuates high-frequency responses. Tethering may also affect the processing of auditory information within the locust CNS by removing sensory input normally present during flight, such as an optical flow field (Baader 1991; Baader et al. 1992; Spork and Preiss 1991), input to wind-sensitive hairs, and proprioceptive feedback from cuticular receptors. The absence of changing visual input may have resulted in locusts disregarding visual cues and shifting attention to (high-frequency) sounds in the environment.
This shift could mean that tethered locusts behave acoustically as locusts on a substrate. When a locust is on the ground sound plays an important role in its biology, and like other orthopterans, the ability to distinguish different types of sounds in the environment is adaptive. For crickets, pitch is one cue for categorical discrimination (Wyttenbach et al. 1996). For locusts, low-frequency sounds such as stridulation, mandible clicking, wing-beat noise, and even wind are not aversive stimuli, whereas higher-frequency sounds, such as might be produced by gleaning bats and rodents, or coincident sounds such as rustling vegetation produced by terrestrial vertebrate predators, signal potential dangers (Sales and Pye 1974; Fullard 1988). The eastern sword-bearer conehead, Neoconocephalus ensiger, possesses an ASR in which it ceases or pauses calling when stimulated with high-frequency, but not low frequency, sounds (Faure and Hoy 2000). Tethered locusts may categorically discriminate sounds and produce avoidance reactions preferentially to high frequency stimuli. Free-flying locusts, lacking the mechanical interference of a tether, might not gate their responses according to frequency. In this case, a free-flying locust may openly gate all auditory responses to an avoidance reaction. This would be a conservative response to all sounds, both alluring and aversive, but would be adaptive when the source of sounds is a predator such as an insectivorous bat. Further, if bright illumination suppresses the ASR in free-flying locusts, as discussed above, it had either less effect or no effect on tethered locusts. This could be because tethering prevents motor output from the CNS from effecting changes in visual input (i.e. opens the sensory-motor loop) and thus vision would become less effective in suppressing the ASR.
Do aerially hawking bats eat locusts?
To our knowledge, the remains of L. migratoria, or other locust species, have not been positively identified in stomach or faecal pellet contents of bats. This result is surprising given that locusts would be a meal of nutritional value to a bat given their high protein and carbohydrate content (Uvarov 1966). Locusts are palatable to birds (Uvarov 1966) and orthopterans in general are palatable to gleaning bats (La Val and La Val 1980; Belwood and Morris 1987). Locusts are also known to fly in open environments at altitudes as high as 100–1000 m during migratory flights and during dispersal flights of solitary-phase individuals (Chapman 1976; Riley and Reynolds 1997). Many large, high-flying, aerial hawking, bats (i.e. the predicted characteristics of bats that might feed on locusts—see Freeman 1981; Barclay and Brigham 1991, 1994; Waters et al. 1995), e.g. Tadarida spp. (Kingdon 1974) are found in the same geographic areas as locusts and use echolocation calls with peak frequencies around 30 kHz, e.g. Taphozous mauritianus, 25 kHz (Fenton et al. 1980); larger Tarida spp., ~25 kHz and Scotophilus nigrita, 30 kHz (Fenton and Bell 1981). These frequencies are in agreement with the best frequency of the behavioural audiogram for the ASR recorded for tethered flight; however, it is not possible at present to know if this syntony is a result of predation pressure.
Free-flying locusts may have low response rates (relative to moths) because they possess additional defences against bat predation. One defence may be in the tendencies of locusts to aggregate in large numbers and fly in large swarms (Uvarov 1943). Swamping is a strategy used by many animals, such as mayflies (Sweeney and Vannote 1982), monarch butterflies (Calvert et al. 1979; Brower and Calvert 1985), and minnows (Hager and Helfman 1991) for protection from predators. Detecting the echolocation calls of bats is not the only way of avoiding predation by bats. Many moth species (both eared and earless) avoid bats by flying at times that bats are not active, flying at altitudes different than bats, or flying erratically (Roeder 1974; Morrill and Fullard 1992; Lewis et al. 1993). Others (e.g. Saturniidae, Sphingidae) enjoy protection by their size and agility in flight (Roeder 1974). It is not unreasonable to think that the large size of locusts, and their agile flight and flight heights offers them additional mechanisms of protection from bats. It must be noted, however, that possessing alternative defensives to auditory detection of bats is not in itself sufficient to cause a decrease in sensitivity of the ear to high-frequency sounds (Fullard and Dawson 1999).
Conclusions
This paper has shown that free-flying locusts produce a negative phonotactic response to sound. The response is an acoustic startle response that is a general startle response to sound and is not specifically a bat-avoidance reaction. The life history of L. migratoria raises the question of whether the absence of high-frequency sensitivity in the ASR in free flight is because locusts possess additional non-acoustic defences from insectivorous bats, or because tethering, and possibly other factors, influence auditory processing during flight. These questions can only be resolved by studies that directly observe locusts and bats interacting in the wild and studies aimed at understanding multimodal sensorimotor integration mechanisms (i.e. a neuroethological approach). Finally, we caution against concluding that tethering adversely affects the behaviour of locusts (or other insects) in all ways. The consistency of responses of tethered locusts to different stimuli (auditory, visual, thermal) suggests the wing kinematic and postural adjustments evoked by these stimuli are important for steering and are not artefacts of tethering.
Abbreviations
- ASR:
-
acoustic startle response
- dB SPL:
-
decibel sound pressure level (RMS re: 20 μPa)
References
Baader A (1991) Auditory interneurons in locusts produce directional head and abdomen movements. J Comp Physiol A 169:87–100
Baader A, Schäfer M, Rowell CHF (1992) The perception of the visual flow field by flying locusts: A behavioural and neuronal analysis. J Exp Biol 165:137–160
Barclay RMR, Brigham RM (1991) Prey detection, dietary niche breadth, and body size in bats: why are aerial insectivorous bats so small? Am Nat 137:693–703
Barclay RMR, Brigham RM (1994) Constraints on optimal foraging: a field test of prey discrimination by echolocating insectivorous bats. Anim Behav 48:1013–1021
Bauer R (1993) Untersuchung zur Anlockung von nachtaktiven Insekten durch Beleuchtungseinrichtungen. Diploma thesis, University of Konstanz
Belwood JJ, Morris GK (1987) Bat predation and its influence on calling behaviour in neotropical katydids. Science 238:64–67
Brower LP, Calvert WH (1985) Foraging dynamics of bird predators on overwintering monarch butterflies in Mexico. Evolution 39:852–868
Calvert WH, Hedrick LE, Brower LP (1979) Mortality of the monarch butterfly (Danaus plexippus L.): avian predation at five overwintering sites in Mexico. Science 204:847–851
Chapman RF (1976) A biology of locusts. Edward Arnold, London
Dawson JW, Dawson-Scully K, Robert D, Robertson RM (1997) Forewing asymmetries during auditory avoidance in flying locusts. J Exp Biol 200:2323–2335
Doherty J, Hoy R (1985) Communication in insects. III. The auditory behaviour of crickets: some views of genetic coupling, song recognition, and predator detection. Q Rev Biol 60:457–472
Farrow RA (1990) Flight and migration in acridoids. In: Chapman RF, Joern A (eds) Biology of grasshoppers. Wiley, New York, pp 227–314
Faure PA, Hoy RR (2000) The sounds of silence: cessation of singing and song pausing are ultrasound-induced acoustic startle behaviors in the katydid Neoconocephalus ensiger (Orthoptera; Tettigoniidae). J Comp Physiol A 186:129–142
Fenton MB, Bell GP (1981) Recognition of species of insectivorous bats by their echolocation calls. J Mammal 62:233–243
Fenton MB, Bell GP, Thomas DW (1980) Echolocation and feeding behaviour of Taphozous mauritianus (Chiroptera: Emballonuridae). Can J Zool 58:1774–1777
Forster W, Wohlfahrt TA (1960) Die Schmetterlinge Mitteleuropas. Band III. Spinner und Schwärmer (Bombyces und Sphinges). Franckh’sche Verlagshandlung. Keller, Stuttgart
Forster W, Wohlfahrt TA (1971) Die Schmetterlinge Mitteleuropas. Band IV. Eulen (Noctuidae). Franckh’sche Verlagshandlung. Keller, Stuttgart
Forster W, Wohlfahrt TA (1981) Die Schmetterlinge Mitteleuropas. Band V. Spanner (Geometridae). Franckh’sche Verlagshandlung. Keller, Stuttgart
Freeman PW (1981) Correspondence of food habits and morphology in insectivorous bats. J Mammal 62:166–173
Fullard JH (1988) The tuning of moth ears. Experientia 44:423–428
Fullard JH, Dawson JW (1999) Why do diurnal moths have ears? Naturwissenschaften 86:276–279
Fullard JH, Yack JE (1993) The evolutionary biology of insect hearing. Trends Ecol Evol 8:248–252
Gatehouse AG, Zhang XX (1995) Migratory potential in insects: variation in an uncertain environment. In: Drake VA, Gatehouse AG (eds) Insect migration: tracking resources through space and time. Cambridge University Press, London, pp 193–242
Hager MC, Helfman GS (1991) Safety in numbers: shoal size choice by minnows under predatory threat. Behav Ecol Sociobiol 29:271–276
Haskell PT (1957) The influence of flight noise on the behaviour in the desert locust Schistocerca gregaria (Forsk.). J Insect Physiol 1:52–75
Hedwig B (1988) Modulation of auditory information processing in tethered flying locusts. J Comp Physiol A 164:409–422
Hoy RR (1989) Startle, categorical response, and attention in acoustic behavior of insects. Annu Rev Neurosci 12:355–377
Hoy RR (1992) The evolution of hearing in insects as an adaptation to predation from bats. In: Webster DB, Fay RR, Popper AN (eds) The evolutionary biology of hearing. Springer, Berlin Heidelberg New York, pp 115–129
Hoy RR, Robert D (1996) Tympanal hearing in insects. Annu Rev Entomol 41:433–450
Hoy RR, Nolan T, Brodfuehrer P (1989) The neuroethology of acoustic startle and escape in flying insects. J Exp Biol 146:287–306
Kalmring K (1975) The afferent auditory pathway in the ventral cord of Locusta migratoria (Acrididae). II. Responses of the auditory ventral cord neurons to natural sounds. J Comp Physiol 104:143–159
Kennedy JS (1956) Phase transformation in locust biology. Biol Rev 31:349–370
Kingdon J (1974) East African mammals: an atlas of evolution in Africa, vol II, part A (insectivores and bats). University of Chicago Press, Chicago
Kutsch W (1999) Telemetry in insects: the “intact animal approach”. Theor Biosci 118:29–53
Kutsch W, Fuchs U (2000) Locust flight initiation: a comparisons of normal and artificial release. Physiol Entomol 25:370–382
La Val RK, La Val ML (1980) Prey selection by a neotropical foliage-gleaning bat, Micronycteris megalotis. J Mammal 61:327–330
Lewis FP, Fullard JH, Morrill SB (1993) Auditory influences on the flight behaviour of moths in a Nearctic site. II. Flight times, heights, and erraticism. Can J Zool 71:1562–1568
Libersat F, Hoy RR (1991) Ultrasonic startle behavior in bushcrickets (Orthoptera; Tettigoniidae). J Comp Physiol A 169:507–514
Miller LA, Olesen J (1979) Avoidance behavior in green lacewings. I. Behavior of free flying green lacewings to hunting bats and ultrasound. J Comp Physiol 131:112–120
Moiseff A, Hoy RR (1983) Sensitivity to ultrasound in an identified auditory interneuron in the cricket: a possible neural link to phonotactic behavior. J Comp Physiol 152:155–167
Moiseff A, Pollack G S, Hoy RR (1978) Steering responses of flying crickets to sound and ultrasound: mate attraction and predator avoidance. Proc Natl Acad Sci USA 75:4052–4056
Morrill SB, Fullard JH (1992) Auditory influences on the flight behaviour of moths in a Nearctic site. I. Flight tendency. Can J Zool 70:1097–1101
Nolen TG, Hoy RR (1986) Phonotaxis in flying crickets. I. Attraction to the calling song and avoidance of bat-like ultrasound are discrete behaviours. J Comp Physiol A 159:423–439
Pollack GS, Hoy RR (1981) Phonotaxis in flying crickets: neural correlates. J Insect Physiol 27:41–45
Pollack GS, Hoy RR (1989) Evasive acoustic behavior and its neurobiological basis. In: Huber F, Moore T, Loher W (eds) Cricket behaviour and neurobiology. Cornell University Press, Ithaca, New York, pp 340–363
Pollack GS, Huber F, Weber T (1984) Frequency and temporal patter-dependent phonotaxis of crickets (Teleogryllus oceanicus) during tethered flight and compensated walking. J Comp Physiol A 154:13–26
Riley JR, Reynolds DR (1997) Vertical-looking radar as a means to improve forcasting and control of desert locusts. In: Krall S, Peveling R, Ba Diallo D (eds) New strategies in locust control. Birkhäuser, Basel, Switzerland, pp 47–54
Robert D (1989) The auditory behaviour of flying locusts. J Exp Biol 147:279–301
Robert D, Rowell CHF (1992) Locust flight steering. II. Acoustic avoidance manoeuvres and associated head movements, compared with correctional steering. J Comp Physiol A 171:53–62
Roeder KD (1962) The behaviour of free flying moths in the presence of artificial ultrasonic pulses. Anim Behav 10:300–304
Roeder KD (1964) Aspects of the noctuid tympanic nerve response having significance in the avoidance of bats. J Insect Physiol 10:529–546
Roeder KD (1967) Nerve cells and insect behaviour. Harvard University Press, Cambridge, MA
Roeder KD (1974) Acoustic sensory responses and possible bat-evasion tactics of certain moths. Proceedings of the Canadian Society of Zoologists Annual Meeting, pp 71–78
Rydell J, Skals N, Surlykke A, Svensson M (1997) Hearing and bat defence in geometrid winter moths. Proc R Soc Lond Ser B 264:83–88
Sales G, Pye D (1974) Ultrasonic communication by animals. Chapman & Hall, London
Simmons JA, Fenton MB, O’Farrell MJ (1979) Echolocation and pursuit of prey by bats. Science 203:16–21
Spork P, Preiss R (1991) Control of flight by means of lateral visual stimuli in the desert locust, Schistocerca gregaria. In: Elsner N, Penzlin H (eds) Synapse-transmission-modulation. Proc 19th Göttingen Neurobiol Conf. Thieme, Stuttgart
Stapells DR, Picton TW, Smith AD (1982) Normal hearing thresholds for clicks. J Acoust Soc Am 72:74–79
Surlykke A, Filskov M (1997) Hearing in geometrid moths. Naturwissenschaften 84:356–359
Surlykke A, Moss CF (2000) Echolocation behaviour of big brown bats, Eptesicus fuscus, in the field and laboratory. J Acoust Soc Am 108:2419–2429
Svensson AM, Rydell J (1998) Mercury vapour lamps interfere with the bat defense of tympanate moths (Operophtera spp.; Geometridae). Anim Behav 55:223–226
Sweeney BW, Vannote RL (1982) Population synchrony in mayflies: a predator satiation hypothesis. Evolution 36:810–821
Uvarov BP (1943) The locust plague. J R Soc Arts 91:109–118
Uvarov BP (1966) Grasshoppers and locusts: a handbook of general acridology, vol 1. Anatomy, physiology, development, phase polymorphism, introduction to taxonomy. Cambridge University Press, Cambridge and Centre for Overseas Pest Research, London
Uvarov BP (1977) Grasshoppers and locusts: a handbook of general acridology, vol 2. Behaviour, ecology, biogeography, population dynamics. Cambridge University Press, Cambridge and Centre for Overseas Pest Research, London
Waters DA, Rydell J, and Jones G (1995) Echolocation call design and limits on prey size: a case study using the aerial-hawking bat Nyctalus leisleri. Behav Ecol Sociobiol 37:321–328
Wyttenbach RA, May ML, Hoy RR (1996) Categorical perception of sound frequency by crickets. Science 273:1542–1544
Yack JE, Fullard JH (2000) Ultrasonic hearing in nocturnal butterflies. Nature 403:265–266
Yager DD, May ML (1990) Ultrasound-triggered, flight-gated evasive maneuvers in the praying mantis Parasphendale agrionina. II. Tethered flight. J Exp Biol 152:41–58
Yager DD, Spangler HG (1997) Behavioral response to ultrasound by the tiger beetle Cicindela marutha Dow combines aerodynamic changes and sound production. J Exp Biol 200:649–659
Yager DD, May ML, Fenton MB (1990) Ultrasound-triggered, flight-gated evasive maneuvers in the praying mantis Parasphendale agrionina. I. Free flight. J Exp Biol 152:17–39
Zar JH (1984) Biostatistical analysis, 2nd edn. Prentice-Hall, Englewood Cliffs, New Jersey
Acknowledgements
From the Universität Konstanz, we thank Holger Martz, Sebastian Berger, Urs Fuchs and Tobias Mueller for technical assistance, Dr. Hanno Fischer for valuable advice, and Prof. Dr. Wolfgang Kirchner for the loan of certain acoustic equipment and also for providing valuable advice. Dr. James Fullard (University of Toronto) kindly read an early draft of the manuscript and offered many valuable suggestions. We also thank the anonymous reviewers of this paper for their input and constructive criticism. This project was funded by grants from the Natural Science and Engineering Research Council of Canada (NSERC) to R.M.R. and by a grant from the Deutsche Forschungsgemeinschaft (DFG Ku 240/17-2) to W.K. The experiments described in this manuscript comply with the Principles of animal care, publication No. 86-23, revised 1985 of the National Institutes of Health and also with the current laws of Germany and Canada.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Dawson, J.W., Kutsch, W. & Robertson, R.M. Auditory-evoked evasive manoeuvres in free-flying locusts and moths. J Comp Physiol A 190, 69–84 (2004). https://doi.org/10.1007/s00359-003-0474-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00359-003-0474-3