
Abstract
Leaf senescence represents a key developmental process through which resources trapped in the photosynthetic organ are degraded in an organized manner and transported away to sustain the growth of other organs including newly forming leaves, roots, seeds, and fruits. The optimal timing of the initiation and progression of senescence are thus prerequisites for controlled plant growth, biomass accumulation, and evolutionary success through seed dispersal. Recent research has uncovered a multitude of regulatory factors including transcription factors, micro-RNAs, protein kinases, and others that constitute the molecular networks that regulate senescence in plants. The timing of senescence is affected by environmental conditions and abiotic or biotic stresses typically trigger a faster senescence. Various phytohormones, including for example ethylene, abscisic acid, and salicylic acid, promote senescence, whereas cytokinins delay it. Recently, several reports have indicated an involvement of auxin in the control of senescence, however, its mode of action and point of interference with senescence control mechanisms remain vaguely defined at present and contrasting observations regarding the effect of auxin on senescence have so far hindered the establishment of a coherent model. Here, we summarize recent studies on auxin-related genes that affect senescence in plants and highlight how these findings might be integrated into current molecular-regulatory models of senescence.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Leaf development is a complex process that starts with primordium initiation at the shoot apical meristem (SAM) and includes cell proliferation, cell elongation, leaf maturation, and induction and execution of senescence once leaves have reached a certain age. As leaves are successively formed during stem growth, they are by nature of different ages at each time point of the stem’s development. Thus, typically young leaves coexist with mature and senescent leaves on the same stem. Leaf senescence is a highly regulated process that must be appropriately integrated into whole-stem and whole-plant developmental processes.
Senescence involves the degradation of major cellular components and their transport out of the deteriorating tissue to feed other developing organs including newly forming leaves during vegetative growth or seeds when the plant has entered the reproductive phase. Although senescence is clearly a developmentally controlled process, environmental factors can have a large impact on the timing and speed of progression of senescence. A number of abiotic stresses can interfere with the typical developmental program of a given leaf (or plant) and thus affect its life span. Abiotic stresses that hasten senescence are water limitation (drought), high temperature, soil salinity, nutrient (for example, nitrogen) limitation, besides others. As an ordered onset and progression of senescence is vital to the ecological success of a plant, major variations with respect to the timing and progression of senescence exist between accessions, cultivars, and species.
Research over the last years has uncovered many molecular elements of the cellular regulatory networks that control senescence, among them being transcription factors (TFs) that bind to cis-regulatory elements in target gene promoters to affect their expression level (thus, leading to their up- or down-regulation). Numerous TFs have been reported to undergo senescence-dependent expression changes in leaves and other organs, including many members of the NAC and WRKY families, besides others (Buchanan-Wollaston and others 2005; van der Graaff and others 2006; Balazadeh and others 2008; Breeze and others 2011). Both, positive and negative regulators of senescence have been identified among senescence-associated TFs (Miao and others 2004; Balazadeh and others 2010a; Besseau and others 2012; Wu and others 2012; Zhang and Gan 2012; Hickman and others 2013; Kim and others 2013). Among the family of NAC TFs, ORE1 (also called ANAC092) has been shown to be a central positive regulator of senescence in Arabidopsis thaliana (Kim and others 2009; Balazadeh and others 2010a; Rauf and others 2013). Downstream genes directly or indirectly controlled by ORE1 have been discovered (Balazadeh and others 2010a; Breeze and others 2011; Matallana-Ramirez and others 2013). Other positive NAC regulators of senescence include, for example, AtNAP/ANAC029 (Guo and Gan 2006) and ORS1 (Balazadeh and others 2011). Recent evidence shows that AtNAP regulates leaf senescence in part by controlling the expression of SAG113, to which promoter it binds (Zhang and Gan 2012). SAG113 encodes an ABA- and senescence up-regulated protein phosphatase (PP) 2C localized in the cis-Golgi apparatus. It functions as a negative regulator of ABA signaling: during senescence, when SAG113 expression increases, it suppresses stomatal closing thereby leading to rapid water loss from leaves and desiccation (Zhang and Gan 2012; Zhang and others 2012). An example for a negative NAC senescence regulator is JUB1 (ANAC042), which delays senescence by lowering cellular H2O2 levels to balance the effect of positive senescence regulators (Wu and others 2012).
Many phytohormones are known for their capacity to modulate the onset or progression of leaf senescence when applied externally or altered in vivo through genetic means. Typically, ethylene, abscisic acid, salicylic acid, and jasmonic acid promote senescence, whereas auxin, gibberellic acids (GAs), and cytokinins retard it (Gan 2005; Lim and others 2007; Schippers and others 2007; Jibran and others 2013). Hormones can affect senescence through various signaling pathways (Guo and Gan 2005; Jibran and others 2013; Zhang and Zhou 2013) and by modulating aging and senescence at different levels of leaf development. For example, in Arabidopsis senescence can normally not be induced in young leaves, likely due to the absence of age-related factors that only accumulate during leaf aging. Such age-related changes (ARCs) appear to be a critically important prerequisite for senescence to occur. Dijkwel and colleagues define an ARC as “any irreversible change that is strictly dependent on age and thus development”, which in leaves is represented by the end of cell division or the cessation of cell expansion, but in principle could be any molecular or cellular change that results from developmental progression (see Jibran and others 2013, for a detailed discussion). The effect of hormones on senescence is then expected to not only depend on environmental impact, but also on the developmental age of the leaf.
Although auxin has been identified as a phytohormone that functions as a regulator of senescence, its mode of action with respect to senescence is only vaguely defined at present, and contrasting observations regarding the effect of auxin on senescence have so far hindered the establishment of a coherent model. Here, we summarize recent studies on auxin-related genes that were reported to affect senescence in plants and discuss how these findings might be integrated into our current molecular-regulatory models of senescence.
Overexpression of YUCCA Genes Delays Senescence
YUCCA genes encode flavin-containing monooxygenases (FMOs) that catalyze the rate-limiting step of auxin biosynthesis via the tryptamine pathway (representing one of several pathways through which auxin is produced); 11 YUCCA (YUC) genes are known in Arabidopsis (Zhao and others 2001; www.arabidopsis.org). Mutational studies have led to the discovery of several biological functions of YUC genes in processes like embryogenesis, leaf formation, floral patterning, vascular tissue formation, high temperature-induced hypocotyl elongation, and others (Cheng and others 2006; Cheng and others 2007; Sun and others 2012). Recently, Kim and others (2011) reported about the effect of elevated YUC6 expression in transgenic Arabidopsis plants (Fig. 1). The dominant yuc6-1D mutant has elevated levels of free indole-3-acetic acid (IAA) in several organs tested, including inflorescences, cauline leaves, and shoots. The increases in auxin levels were most prominent in the tissues that showed the most severe phenotypic changes, namely inflorescences and cauline leaves (Kim and others 2007).

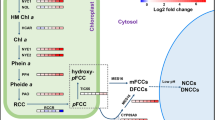
Model of the integration of auxin-related genes with senescence. The transcription factor ARF2 represses the expression of the GATA genes GNC and GNL by directly binding to their promoters, concomitant with a reduction of chlorophyll accumulation, thus contributing to shifting the regulatory balance toward senescence. In addition to ARF2, ARF7 also has the capacity to bind to the two GATA gene promoters. As suggested by Richter and others (2013), ARF2 might act in an auxin-independent manner, whereas ARF7 might function in an auxin-dependent manner to regulate GNC/GNL expression. Enhanced expression of YUC6 causes an increased free auxin level (Kim and others 2011) leading to auxin overproducer phenotypes and extended plant longevity, apparent through a continued formation of new shoots. Dark-induced expression of ORE1 is reduced in the yuc6-1D mutant (a YUC6 overexpressor) compared to wild type, in accordance with the delayed senescence observed in the mutant. As ORE1 is a key positive regulator of leaf senescence, its reduced expression in the yuc6-1D mutant is expected to limit the expression of its senescence-associated target genes, thus contributing to the observed delayed senescence. SAGs senescence-associated genes (that is, genes up-regulated during developmental senescence). Transcription factors are shown in bold. Positive regulations are indicated by arrows, negative regulations by T-ending lines
Another prominent phenotype realized in the yuc6-1D mutant, as well as in 35S:YUC6 overexpressors, was a delay of senescence (Kim and others 2011). Of note, 5-month-old yuc6-1D mutants even produced new shoots and flowers, reminiscent of a prolonged plant life span. The authors also tested dark-induced senescence in detached leaves and observed a less rapid decline of chlorophyll levels and photosystem II efficiency in yuc6-1D and 35S:YUC6 plants. In accordance with the delayed senescence in darkness, the expression of SAG12 increased less rapidly in plants with high YUC6 expression.
The effect of elevated YUC6 expression on delaying senescence was further tested using transgenic plants overexpressing a mutated YUCCA6 (mYUC6) protein (G206V, located within the enzyme’s NADPH-binding site). Although yuc6-1D plants overexpressed many auxin-responsive genes including IAA2 (Kim and others 2007, 2011), IAA2 expression was not enhanced in the mYUC6 transgenic plants (Kim and others 2011), indicating that free auxin levels were not increased in the latter, consistent with the expectation that mYUC6 is catalytically inactive. This conclusion is supported by the absence of curly leaves which are typically observed in auxin overproducers. Dark-induced senescence parameters were similar in 35S:mYUC and wild-type plants.
Is Elevated Free Auxin Level Causative of Delayed Senescence?
Although the above experiment indicated that elevated auxin levels caused the delay of senescence, direct proof for this hypothesis was missing. Therefore, in a follow-up experiment, the authors crossed a plant overexpressing IAA-lysine synthetase (iaaL) from Pseudomonas savastanoi with the yuc6-1D mutant, to eliminate the potential effect of elevated IAA. In accordance with a lowered free auxin level in F1 plants (35S:iaaL × yuc6-1D) compared to yuc6-1D, hypocotyl elongation and IAA2 expression were reduced. With respect to dark-induced senescence, chlorophyll loss in leaves was not delayed as in yuc6-1D, whereas it was more rapid in the 35S:iaaL plant compared to wild type, consistent with the model that elevated auxin level causes the delay in senescence (Kim and others 2011). In accordance with this model, free IAA levels as well as YUC6 expression decreased in detached wild-type leaves subjected to dark-induced senescence. In contrast, expression of several GH3 family genes, including GH3.1, GH3.3, GH3.5, and GH3.6, increased in darkness. IAA-amido synthetases encoded by GH3 genes conjugate excess IAA to amino acids thereby contributing to auxin homeostasis (Staswick and others 2005). Additional GH3 genes were found to be up-regulated during developmental and dark-induced senescence in global transcriptome studies (Buchanan-Wollaston and others 2005; van der Graaff and others 2006), suggesting a wider role of auxin conjugation during senescence.
The conclusion that elevated auxin delays senescence is supported by the observation that similar to YUC6 the overexpression of YUC1 as well as the application of α-naphthalene acetic acid (NAA, a synthetic auxin) to wild-type leaves delays dark-induced senescence (Kim and others 2011).
Recently, Lee and others (2012) reported that enhanced expression of YUC7 in an activation-tagged Arabidopsis mutant (yuc7-1D) and a constitutive overexpression line (35S:YUC7) coincides with phenotypic changes typically observed in auxin-overproducing plants, including slender stems and curled and narrow leaves. Although total auxin level was elevated in yuc7-1D, an increase in free auxin level was not found. However, expression of the auxin-responsive genes IAA5, IAA19, GH3.3, and GH3.5 was increased in the yuc7-1D mutant, indicating an elevation of free IAA levels. A senescence-related phenotype was not reported, but drought tolerance was found to be enhanced in the yuc7-1D mutant (Lee and others 2012). Furthermore, no overt phenotype was seen in a YUC7 knockout mutant (yuc7-1).
The Potential Role of NAC Transcription Factor ORE1 in Auxin-Mediated Senescence Retardation
Transcription factors of the NAC family play important regulatory roles in senescence (for example, Guo and Gan 2006; Yoon and others 2008; Kim and others 2009; Balazadeh and others 2010a, 2011; Yang and others 2011; Wu and others 2012). One of the best characterized positive regulatory NAC factors of senescence is ORE1 (also called ANAC092; Kim and others 2009; Balazadeh and others 2010a; Trivellini and others 2012; Rauf and others 2013). Expression of ORE1 increases in leaves during developmental senescence (Balazadeh and others 2008). Similarly, ORE1 expression increased in detached wild-type leaves during dark-induced senescence, but its transcript accumulated much less prominently in the yuc6-1D mutant under the same condition (Kim and others 2011). ORE1 has been shown to regulate the expression of an appreciable number of the known senescence-associated genes (SAGs) (Balazadeh and others 2010a; Breeze and others 2011; Matallana-Ramirez and others 2013); thus, the delayed senescence observed in the high-free-auxin yuc6-1D mutant may in part be controlled through the lowered induction of ORE1 during dark incubation (Fig. 1; Kim and others 2011). Currently, however, it is unknown how exactly high auxin levels affect the expression of ORE1, in particular as auxin treatment only slightly alters its transcript abundance (www.genevestigator.com).
The Role of ARF2 as a Senescence Regulator
Lim and others (2010) performed a screen on ethyl methane sulphonate (EMS)-mutagenized populations of Arabidopsis to discover mutants showing delayed leaf senescence. The initial screen was performed by incubating detached leaves in the dark, a simple method often used to promote senescence. The screen discovered two allelic oresara14 mutants, ore14-1 and ore14-2 (“oresara” stands for “long-living” in Korean), and MAP-based cloning revealed ORE14 to encode AUXIN RESPONSE FACTOR2 (ARF2), a transcription factor known for its repressor role in auxin-related signaling. ARFs regulate the expression of target genes by binding to TGTCTC auxin response elements (AuxRE) in their promoters. ARFs typically interact with Aux/IAA repressors in an auxin-dependent manner; however, no Aux/IAA protein interacting with ARF2 has so far been reported. The Arabidopsis genome encodes 22 full-length ARF TFs, and a similar number (25 ARFs) is known in rice (Guilfoyle and Hagen 2007).
Sequencing revealed the presence of premature stop codons in both ore14 mutants, indicating these mutations caused a complete loss of ARF2 function (Lim and others 2010). The ore14 mutation triggered a pleiotropic phenotype and besides a delay in leaf and stem senescence it caused late flowering, failure of floral bud opening, and an increase of aerial organ size. Similarly, previous reports discovered multiple phenotypic changes in arf2 T-DNA insertion mutants including large, dark green rosette leaves, delayed senescence, abnormal flower morphology, reduced fertility due to impaired self-pollination (caused by expanded gynoecia failing to properly receive pollen from the shorter stamens), elongated hypocotyls with enlarged cotyledons, and enlarged seed size (Okushima and others 2005; Schruff and others 2006).
Senescence-related phenotypes of ore14/arf2 mutants were carefully analyzed by Lim and others (2010). First, the authors tested leaf longevity during age-dependent senescence by focusing on rosette leaves 3 and 4 of the ore14-1/arf2-10 mutant. In the wild type, leaf 4 started to turn yellow (due to chlorophyll loss) at day 20 after its visible emergence (DAE, days after emergence), whereas in the case of ore14-1/arf2-10, leaf yellowing was observed only 8 days later, that is, at a leaf age of 28 days. Wild-type leaves were completely senescent at 36 DAE; in contrast, ore14-1/arf2-10 showed complete yellowing at 44 DAE (although at this age, the leaves still appeared more intact than the fully senesced wild-type leaves). Thus, lack of ARF2 function appears to mainly cause a delay of the onset of senescence whereas progression of senescence, once initiated, seems to be less affected; the execution of full senescence in both, the wild type and the ore14-1/arf2-10 mutant required approximately 16 days in the Lim and others (2010) study.
Another way of measuring age-dependent senescence is determining the time point at which 50 % of the entire leaf population of a plant is still alive. In the report of Lim and others (2010), this point was reached at 26 DAE in the wild type, but at 35 DAE in the ore14-1/arf2-10 mutant. Thus, the later beginning of senescence in the ore14-1/arf2-10 mutant determined at the individual leaf level was in accordance with the fact that it kept its entire rosette alive longer than the wild type. However, as in the individual leaf measurements, the rate of progression of entire rosette senescence appeared to be similar in the wild-type and mutant plants.
As senescence is affected by various environmental parameters such as darkness, drought or nutrient limitation, the role of a given gene for senescence can also be studied by subjecting plants to senescence-inducing stress treatments. Lim and others (2010) tested the effect of darkness on fully grown leaves (no. 3 and 4) detached at 12 DAE (when leaves were fully green). Chlorophyll loss was followed and after 6 days of dark incubation wild-type leaves had lost 90 % of their initial chlorophyll, while both, ore14-1 and ore14-2 mutants required more than twice as long (approximately 14 days) for 90 % chlorophyll loss. Similarly, photochemical efficiency (Fv/Fm) declined much more rapidly in wild-type than ore14 mutants: no photosystem II (PS II) activity was detectable in the wild type after 8 days of dark incubation, in both ore14 mutants it was around 70 % of the initial value at day 8 and still above 20 % after 14 days of darkness. Thus, the slope of the decrease of PS II activity was largely different in the two sets of plants. Similarly, the increase in ion leakage as an indicator of membrane damage during senescence was much less pronounced in the ore14-1 mutant than the wild type, and expression of SEN4, a marker gene for dark-induced senescence, increased less rapidly. Note, that in these experiments leaves of the same chronological age were analyzed. However, it may well be that the biological (physiological) age of leaves 3 and 4 of the two types of plants, that is, wild type and ore14 mutants, may have been different and that the ore14 mutants may have been physiologically younger at the same chronological age than the wild-type plants. In this scenario, dark-induced senescence could be primed to progress faster in the wild type than the ore14 mutants.
The authors also tested the effect of three senescence-promoting phytohormones, namely abscisic acid (ABA), methyl jasmonate (MeJA), and ethylene, on detached leaves and for all three hormones a more rapid decline in chlorophyll content and PS II activity was observed in the wild type than the ore14-1 mutant (Lim and others 2010).
A role of ARF2 for age-dependent and induced senescence has independently been reported by Ellis and others (2005). These authors also showed that ARF1 on its own had little effect on senescence, but had the capacity to enhance the effect of ARF2 on senescence. ARF2 transcript abundance increased during dark-induced senescence, similar to SAG12 (Ellis and others 2005). Furthermore, analyzing P ARF2 :GUS-ARF2 transgenic plants which express a GUS-ARF2 fusion protein from the ARF2 promoter (Li and others 2004), the authors observed an upregulation of GUS activity in detached dark-induced leaves, suggesting an increase in GUS-ARF2 fusion protein level under these conditions (Fig. 1; Ellis and others 2005). In contrast, the expression level of ARF1 slightly decreased upon dark incubation, indicating that ARF1 and ARF2 play different roles in senescence.
Besides ARF2, ARF7 (identical to NPH4), and ARF19 are also induced in senescing Arabidopsis leaves (Lin and Wu 2004; Ellis and others 2005). Although mutating these genes individually did not affect senescence of rosette leaves or floral organs, an arf2-8 nph4-1 arf19-4 triple mutant was more retarded in detached leaf senescence than the arf2-8 single-gene mutant, indicating that ARF7/NPH4 and ARF19 enhanced the effect of ARF2.
An important and currently unresolved question is through which molecular or physiological mechanism ARF2 affects the aging of the plant tissue. An earlier observation of Ohto and others (2006) might offer a potential link between auxin signaling and senescence-related physiology. These authors discovered that a loss-of-function mutant of the HOOKLESS1 (HLS1) gene shows a defect in sugar-responsive gene expression. In particular, the expression response to exogenous sucrose was more sensitive in the hls1 mutant than the wild type. Application of auxin (IAA) partially repressed the response to exogenous sugar and at the same time led to the activation of some auxin-responsive genes. Furthermore, compared to the wild type the level of free IAA was reduced in hsl1 leaves. Interestingly, ARF2 is known to act downstream of HLS1 (Li and others 2004). Thus, HLS1 and ARF2 may together establish a regulatory frame that links auxin with sugar signaling to control tissue aging and senescence, although the regulatory interaction of these proteins during leaf senescence has not been studied yet. Notably, an increase of sugar levels during developmental leaf senescence has frequently been observed (for example, Diaz and others 2005; Pourtau and others 2006; Wingler and others 2006; Watanabe and others 2013). However, because both sugar starvation and treatment can trigger the expression of SAGs (Gan and Amasino 1997; Fujiki and others 2000, 2001; van Doorn 2004) their precise roles in senescence remain controversial (Yoshida 2003; van Doorn 2004, 2008; Wingler and others 2006, 2009).
Another link between ARF2 and leaf senescence was recently revealed by Richter and others (2013). These authors discovered that ARF2 represses the expression of the two GATA transcription factors GNC (for GATA, NITRATE-INDUCIBLE, CARBON-METABOLISM INVOLVED) and GNL (for GNC-LIKE; also called CGA1 for CYTOKININ-RESPONSIVE GATA FACTOR1) by directly binding to their promoters. Expression of GNC and GNL is enhanced in the arf2 mutant; notably, both genes are also higher expressed in an arf7 arf19 double mutant and this is further enhanced in an arf2 arf7 arf19 triple mutant. Similar to ARF2, direct binding to GNC and GNL promoters could also be demonstrated for ARF7. Notably, like the arf2 mutant, GNC and GNL overexpressors accumulate more chlorophyll than the wild type and they senesce later, a phenotype that is consistent with the model that ARFs repress GNC and GNL transcription (Richter and others 2013). As there is currently no evidence for an interaction of ARF2 with Aux/IAA repressors, it might function in an auxin-independent manner, whereas ARF7, which interacts with Aux/IAA14 (Fukaki and others 2005), might act in dependence of auxin. Thus, ARF7 might modulate the effect of ARF2 on GNC/GNL in an auxin-dependent manner (Richter and others 2013). Furthermore, it was previously shown, that the two GATA factors are repressors of GA signaling (Richter and others 2010). However, experimental evidence indicates that GA and auxin signaling act through independent pathways to control GNC and GNL expression (Richter and others 2013).
SAUR36 in Arabidopsis
SMALL AUXIN UP-REGULATED RNA (SAUR) genes represent a large family of auxin-responsive genes in Arabidopsis and other plants (Hagen and Guilfoyle 2002; Jain and others 2006). Although the biological function of most SAUR genes remains unknown, experimental evidence indicates an involvement of some of them in cell expansion (Esmon and others 2006; Spartz and others 2012). Recently, it was reported that SMALL AUXIN UP RNA (SAUR) 36, which is identical to SENESCENCE-ASSOCIATED GENE201 (SAG201), functions as a positive regulator of leaf senescence in Arabidopsis (Fig. 2). SAUR36 is a nuclear-targeted protein (shown by GFP fusion studies; Narsai and others 2011). Expression of SAUR36 is low in young and fully expanded leaves, but enhanced in senescent leaves as shown by microarray-based expression profiling, qRT-PCR and promoter-GUS studies (Guo and others 2004; Hou and others 2013). SAUR36 expression is also induced in intact plants during dark-induced senescence (observed in a transcriptome dataset by Lin and Wu 2004).

In Arabidopsis, SAUR36 links auxin signaling with senescence. Note, that SAUR36 inhibits cell elongation and leaf size, whereas it promotes the onset, but less evidently the progression of senescence. GA 3 gibberellic acid, IAA indole-3-acetic acid, NAA α-naphthalene acetic acid
Furthermore, SAUR36 is rapidly (within 30 min) induced in leaves of 3-week-old plants by auxin (NAA) treatment, and expression rises to high levels within 6 h (Hou and others 2013). Auxin (IAA)-induced SAUR36 expression was also reported by Stamm and Kumar (2013) in 3-day-old seedlings, whereas expression is repressed by treatment with the auxin transport inhibitor 1-N-naphthylphthalamic acid (NPA). SAUR36 expression is suppressed by treatment with gibberellic acid (GA3), but stimulated when seedlings are treated with the gibberellin biosynthesis inhibitor paclobutrazol (PAC). Considering its response to auxin and GA treatment, Stamm and Kumar (2013) renamed SAUR36 as RESPONSE TO AUXINS AND GIBBERELLINS1 (RAG1). In contrast to the induced expression in leaves and seedlings, previous reports and microarray studies (www.genevestigator.com) revealed either no or a minimal effect of auxin treatment on SAUR36 expression or a strong repression in roots 6 h after auxin treatment (Paponov and others 2008).
Using two different T-DNA insertion lines for SAUR36, Hou and others (2013) observed no major phenotypic change compared to wild type during the early stages of growth and development; however, analysis of physiological data (including leaf mortality, leaf chlorophyll content, ion leakage, and maximal photochemical efficiency of PS II in dark-adapted state) revealed a later onset (by 3–4 days) of senescence in one of the saur36 mutants analyzed, although progression of senescence, once initiated, did not appear to be affected to a large extent. Using transgenic Arabidopsis plants expressing SAUR36 (from which a 3′-UTR-located mRNA destabilizing element had been removed) from a dexamethasone (DEX)-inducible promoter, the authors demonstrated induction of leaf senescence within 3 days of DEX treatment. Enhanced SAUR36 expression after DEX treatment was accompanied by elevated expression of the senescence marker gene SAG12. Stamm and Kumar (2013) observed retarded and twisted growth of the inflorescence axis in 35S:RAG1 (hence, 35S:SAUR36) overexpressors, indicating potential defects in auxin transport as reported by Chae and others (2012) for SAUR63 overexpressors. SAUR36/RAG1 overexpressors also showed reduced fertility, but the cause for this was not analyzed. In addition, seed germination was affected in SAUR36-modified transgenic plants (Stamm and Kumar 2013).
Of note, besides a delayed onset of senescence in the saur36 knockout mutant approximately 83 % increase of organ size of fully expanded leaves was observed (compared to the Col-0 wild type). This rather dramatic increase in leaf size was accompanied by an increase in epidermal cell size by 67 % in the saur36 mutant analyzed, suggesting that SAUR36 functions as an inhibitor of cell expansion (Fig. 2; Hou and others 2013).
SAUR39 in Rice
In rice (Oryza sativa), SAUR39 has been shown to act as a negative regulator of auxin synthesis and transport. Expression of SAUR39 rapidly increases in rice shoots within 30 min after auxin treatment, increases further within a timeframe of 8 h and decreases again thereafter. In leaves, SAUR39 expression was higher in mature than young leaves, and relatively high expression was also observed in senescent leaves (Kant and others 2009). Rice plants overexpressing SAUR39 were smaller than wild-type plants, and primary root length and the number of lateral roots was reduced. Notably, senescence started much earlier in SAUR39 overexpressors than wild-type plants, whereas flowering occurred at approximately the same age in both types of plants (Kant and others 2009). Metabolite studies revealed reduced leaf chlorophyll content in SAUR39 overexpressors compared to wild type, whereas the anthocyanin level was elevated; also ABA level was increased. Transcriptome profiling identified 1,094 genes to be significantly up-, and 411 genes to be down-regulated (by at least 2-fold) in shoots of 4-week-old SAUR39 overexpressors. Although an early onset of senescence was observed in the overexpressor lines, the expression of only five genes with a putative role for senescence was up-regulated compared to wild type, whereas 12 anthocyanin biosynthesis genes were up-regulated (consistent with the elevated anthocyanin level in the transgenic plants). Moreover, in accordance with the reduced chlorophyll level in SAUR39 overexpressors, the expression of 23 genes with putative roles in chlorophyll production and photosynthesis was reduced (Kant and others 2009). Using real-time PCR the authors detected a slight reduction in the expression of some auxin biosynthesis genes (TRP1, TRP2, TRP6, and YUC6) which may cause the reduced free auxin level in these plants.
The regulatory link between SAUR39 and delayed senescence in rice is not particular clear at the current stage. As reported by Kant and others (2009) the early progression of senescence in SAUR39 overexpression plants was accompanied by an increase in sugar and starch content; furthermore, the expression of sugar-responsive genes was elevated, which might be due to the reduced auxin content in these plants. Thus, as in Arabidopsis (see above) a regulatory link might exist between auxin and sugar signaling that might be important for the precise control of leaf senescence also in this species; however, further research is clearly needed to unravel the underlying principles through which auxin affects senescence.
Neutron Irradiation and its Effects on Auxin-Related Genes and Senescence
Previously, the effect of neutron irradiation on senescence was analyzed in Arabidopsis (Fortunati and others 2010). The authors tested the expression of genes connected to auxin signaling, senescence, and oxidative stress in seedlings and mature wild-type plants (ecotypes Col-0 and Ws) after a 3-h exposure to different doses of radiation. Most of the auxin-connected genes tested, including ARF1, ARF2, and ARF19, as well as of the auxin-activated genes Aux/IAA3 and Aux/IAA7, were down-regulated. An opposite expressional response, that is, an upregulation of these genes, was observed in the aux1 mutant, which lacks an auxin influx transporter encoded by the AUX1 gene, and generally also in the pin2 mutant that lacks the PIN2 auxin efflux transporter. In contrast to the auxin-related genes, the SAGs SAG12 and SAG13 were up-regulated upon neutron irradiation in Arabidopsis wild-type plants, whereas expression of both genes was down-regulated in the aux1 mutant (Fortunati and others 2010). Similarly, genes connected to oxidative stress responses, namely CAT1, CAT3, and FeSOD1, were up-regulated in the wild type, but repressed in the aux1 mutant.
The authors also tested the effect of neutron irradiation on the accumulation of thiobarbituric acid reactive substances (TBARS) as an indicator of the lipid state of the membranes and found an increase of TBARS compared to control plants. Additionally, photosystem II activity (Fv/Fm) decreased upon irradiation, as did the expression of the photosynthesis-related genes CAB1 and RUBS-1B (Fortunati and others 2010).
The results reported by Fortunati and others (2010) suggest a role of auxin transport and perhaps auxin signaling for setting the frame of senescence induction and progression.
Senescence-Associated Receptor-Like Kinase (SARK)
GmSARK from soybean (Glycine max) encodes a leucine-rich repeat receptor-like kinase (LRR-RLK). Relatively prominent expression was observed in young leaves, but transcript abundance decreased thereafter in maturing leaves whereas it increased again during senescence. GmSARK expression also increased in dark-incubated detached leaves when senescence was induced and treatment with the cytokinin 6-benzyl aminopurine (6-BA), known to delay senescence in soybean, slowed down the increase of GmSARK expression (Li and others 2006). Knocking down GmSARK expression by RNA interference (RNAi) led to chlorophyll overaccumulation and a retardation of senescence. On the contrary, overexpression of GmSARK reduced chlorophyll levels and strongly enhanced expression of Gmlls1 (a gene proposed to encode an enzyme involved in chlorophyll breakdown), whereas expression of Gmgtr1 (encoding an enzyme of chlorophyll biosynthesis) was reduced (Fig. 3). Overexpression of GmSARK in transgenic Arabidopsis plants triggered a similar (pale-yellow) phenotype in leaves, and strongly impaired shoot development. The authors concluded that GmSARK-mediated signaling may be involved in controlling chloroplast development and chlorophyll accumulation (Li and others 2006). The observed effect on senescence may then result from shifting the balance between chloroplast maintenance toward chloroplast breakdown upon GmSARK-controlled signaling which may be an important control element for the initiation of leaf senescence. Of note, control of senescence by balancing chloroplast maintenance against deterioration has recently been reported to be mediated by the interaction of two types of transcription factors, namely the G2-like transcription factors GLK1 and GLK2 which are important for chloroplast formation (Waters and others 2008. 2009), and ORE1, a key positive regulator of senescence (Kim and others 2009; Balazadeh and others 2010a; Rauf and others 2013).

Model for the role of leucine-rich repeat receptor-like kinase SARK in regulating senescence. In soybean, GmSARK represses expression of Gmgtr1, but enhances expression of Gmlls1, thereby inhibiting chloroplast maintenance leading to leaf yellowing. Besides a promotion of senescence, elevated GmSARK expression leads to multiple morphological changes (for example, dwarfism, formation of curved roots, altered flower morphology). CYP79B2 and TSA1 encode enzymes involved in auxin biosynthesis, whereas GH3.5 encodes an enzyme of the GH3 family involved in conjugating excess free auxin to amino acids thereby contributing to auxin homeostasis. SAG12, WRKY6, and AtNAP are senescence-associated genes
To further evaluate the signaling integration of SARK, the Wang group extended their research toward Arabidopsis (Xu and others 2011). Transgenic Arabidopsis plants expressing GmSARK under the control of a DEX-inducible transcription system (GVG:GmSARK lines) showed a rapid (within 2 h) increase of SAG12 expression after DEX treatment in seedlings. DEX treatment strongly delayed seedling growth and triggered the formation of short and curved roots; the severe growth depression was accompanied by leaf yellowing similar to Arabidopsis seedlings expressing GmSARK from the CaMV 35S promoter (Li and others 2006). Leaf yellowing upon DEX treatment was also observed in adult plants. Expression analysis revealed increased transcript accumulation of the senescence-related transcription factors WRKY6 and AtNAP. Similar to GmSARK, the Arabidopsis homolog AtSARK caused an altered root structure and precocious leaf yellowing (and potentially senescence) upon overexpression, which was accompanied by enhanced expression of senescence-related transcription factors. In contrast, an AtSARK knockout mutant (sark-1) showed a delay in chlorophyll loss and senescence compared to wild type (Xu and others 2011). The sark-1 mutant also produced fever leaves than the wild type, the rosette diameter remained smaller, and bolting and flowering were delayed.
Enhanced Response to Auxin in GmSARK Overexpressors
An interesting aspect is that GmSARK affects auxin responses and distribution when overexpressed in Arabidopsis. Notably, treatment of seedlings transformed with a GmSARK promoter:GUS construct showed significant induction of GUS activity upon treatment with IAA. In addition, extended IAA treatment (96 h) more strongly stimulated the formation of lateral roots in DEX-treated GVG:GmSARK seedlings than in controls, suggesting that GmSARK enhances the sensitivity toward auxin (Xu and others 2011). By employing the DR5:GUS reporter gene the authors further observed that GmSARK altered the cellular distribution of auxin, indicating that signaling through this receptor kinase affects auxin distribution, at least in roots. Furthermore, the expression of two genes related to auxin biosynthesis, namely CYP79B2 and TSA1, as well as the expression of the auxin-responsive gene GH3.5 was significantly elevated upon DEX-induced GmSARK expression, whereas transcript abundance of ARF2 decreased.
DEX-induced GmSARK expression increased the transcript abundance of four ACC synthase genes involved in ethylene biosynthesis, namely ACS4, ACS6, ACS7, and ACS9, but lowered the expression of ACS2, ACS5, and ACS11. The expression changes observed for the different ACS genes are in accordance with the interpretation that GmSARK triggers the formation of more active ACS7 homo- and ACS7-containing heterodimers whereas expression of ACS genes contributing to the formation of inactive heterodimers (that is, ACS2, ACS5, and ACS11; Tsuchisaka and others 2009) was down-regulated. Also the expression of typical ethylene-responsive genes (ERF1, ERF2, ERF4, and AtEBP) was elevated in GmSARK-induced plants, indicating that not only ethylene formation was promoted, but also ethylene signaling (Xu and others 2011). Thus, auxin and ethylene may both involve SARKs to regulate senescence.
Transcriptome Profiling Reveals Limited Overlap Between Senescence- and Auxin-regulated Genes
The reports summarized above indicate a potential role of auxin in the control of senescence-related processes and highlight the fact that several auxin-related genes are indeed induced during developmental or dark-induced senescence. To further investigate the possible involvement of auxin-related processes in senescence, we took advantage of public transcriptome data to analyze the expressional changes of auxin homeostasis and auxin-responsive genes during developmental and dark-induced senescence in Arabidopsis thaliana. First, we compiled a list of 3,716 SAGs (that is, up-regulated) from the reports of Buchanan-Wollaston and others (2005), Balazadeh and others (2008), and Breeze and others (2011). We used the data from Buchanan-Wollaston and others (2005) and Breeze and others (2011) to compile a list of 2,633 senescence-down-regulated genes (SDGs). Genes affected during dark-induced senescence were taken from Lin and Wu (2004); the list comprises 3,230 up- and 2,063 down-regulated genes. As the expression of many senescence-regulated genes is induced by reactive oxygen species (ROS) like hydrogen peroxide (H2O2) (Navabpour and others 2003; Balazadeh and others 2010b, 2011), we also compiled a list of 1,299 genes whose expression is promoted by H2O2 treatment, using transcriptome data generated in-house (unpublished). Finally, we assembled a cohort of auxin-responsive genes from the reports of Zhao and others (2003), Goda and others (2004, 2008), and Nemhauser and others (2004, 2006), yielding 683 auxin up- and 476 down-regulated genes. All genes assembled in this way are listed in Supplemental Table S1.
Next, we searched for genes commonly affected in the various experiments. First, we focused on genes up-regulated during developmental senescence (SAGs) and dark-induced senescence. As shown in Supplemental Table S2, 734 of the 3,716 SAGs (that is, 20 %) are also induced by H2O2-treatment, whereas more than half (57 %) of the H2O2-up-regulated genes are SAGs, which is in accordance with previous reports that presented strong evidence for a role of ROS in senescence (Navabpour and others 2003; Balazadeh et al. 2010a, b; Cui and others 2013; Zhou and others 2013). One-third of all SAGs (that is, 1,232 genes) were also induced during dark-induced senescence, 418 of the dark-induced genes were also up-regulated by H2O2-treatment, and 331 genes overlapped in all three datasets (Supplemental Table S2). Thus, as reported earlier (Buchanan-Wollaston and others 2005; van der Graaff and others 2006), a considerable overlap of gene expression changes is observed during age-dependent and dark-induced senescence, and many genes are also affected when plants are challenged by H2O2 treatment.
When comparing senescence-relevant genes with those affected by auxin, we observed that 207 (30 %) and 127 (19 %) of the 683 auxin-up-regulated genes are SAGs or stimulated by dark-induced senescence, respectively (Fig. 4). On the contrary, auxin-up-regulated genes constitute only approximately 5 % of either SAGs or genes induced by darkness. Although 1,232 genes are both up-regulated during developmental and dark-induced senescence, only 87 genes are induced in all three scenarios, indicating that auxin does not play a major role in triggering the expression of the vast majority of the senescence up-regulated genes, in accordance with the general senescence retarding effect of auxin (Lim and others 2007; Jibran and others 2013). We also looked at the 476 genes down-regulated by auxin treatment and observed that 61 (that is, 13 %) and 40 (8 %) of those are induced during developmental or dark-induced senescence, respectively. However, the auxin-down-regulated genes constitute only less than 2 % of all SAGs or dark-induced genes (that is., 61 or 40 out of 3,716 or 3,230 genes, respectively). Of the 2,633 SDGs, only 99 and 90 genes (that is, 4 %) were down- or up-regulated, respectively, by auxin treatment; similarly, only 88 and 98 of the 2,063 genes down-regulated in darkness (that is, below 5 %) were, respectively, down- or up-regulated by auxin (Fig. 4; Supplemental Table S2). Thus, overall only a relatively small percentage of the SAGs, SDGs or genes affected during dark-induced senescence are auxin-responsive, indicating that a large fraction of the SAG regulatory network is only indirectly affected by auxin.

Venn diagrams of Arabidopsis genes affected at the transcript level during developmental senescence, dark-induced senescence, and by auxin treatment. Numbers of genes commonly changing their expression status are given in the overlapping circle areas. The complete gene lists and experimental conditions are given in Supplemental Tables S1 and S2. SAGs senescence-associated genes (that is, genes induced during developmental senescence), SDGs senescence-down-regulated genes (that is, genes down-regulated during developmental senescence). Genes up- or down-regulated during dark-induced senescence are labeled ‘Dark-UP’ and ‘Dark-DOWN’, respectively, and genes up- or down-regulated by auxin treatment are labeled ‘Aux-UP’ and ‘Aux-DOWN’
Several Auxin-Related Genes are Modulated During Senescence
We next focused our analysis on auxin-related genes, that are responsive to the phytohormone and/or play central roles in auxin metabolism or signaling: SAURs (78 genes; www.arabidopsis.org), Aux/IAAs (29 genes; Liscum and Reed 2002), GH3s (20 genes; Hagen and Guilfoyle 2002), YUCCAs (11 genes; Zhao and others 2001; www.arabidopsis.org), and ARFs (22 genes; Hagen and Guilfoyle 2002), and checked their expression during developmental and dark-induced senescence. Of the 160 genes analyzed in this way, 17 are induced during developmental senescence (8 SAURs, 2 Aux/IAAs, 5 GH3s, 2 ARFs), whereas 22 genes are down-regulated (14 SAURs, 6 Aux/IAAs, 2 ARFs). Similarly, 14 of the 160 genes show enhanced expression during dark-induced senescence (5 SAURs, 3 Aux/IAAs, 3 GH3s, 3 ARFs), and 19 genes show reduced expression (9 SAURS, 8 Aux/IAAs, 1 GH3, 1 ARF) (Supplemental Table S3). Thus, expression of similar numbers of auxin-related genes are induced or repressed during developmental and dark-induced senescence.
Conclusions
The initiation and subsequent progression of senescence towards cell death and final tissue decomposition represents a key developmental process in plants. Given its great importance for whole-plant physiology, growth, and seed yield it is not surprising that senescence is controlled at different levels by multiply interlocked regulatory loops. Senescence, that is, the process that typically involves the decline of chlorophyll level, an increase of ion leakage due to membrane damage, and an elevation of the expression of SAGs, is under considerable developmental control: young leaves typically do not enter senescence as exemplified by the inability of ethylene to induce senescence in immature leaves, which otherwise—in mature leaves—readily stimulates senescence, pointing to the existence of currently vaguely defined age-related factors (see Jibran and others 2013, for a detailed discussion). Several recent reports, as presented above, suggest an involvement of auxin in the control of senescence. However, global expression profiling revealed that auxin-regulated genes are underrepresented in the large set of SAGs, compared to H2O2-regulated genes which show an appreciable overlap with SAGs. Furthermore, similar numbers of auxin-related genes (SAUR, GH3, Aux/IAA, ARF, and YUCCA) are induced and repressed, respectively, during developmental and dark-induced senescence. A possible explanation for these somewhat contrasting observations might be that auxin level and signaling, by affecting leaf (and plant) development, contribute to controlling the establishment of age-related changes (ARCs). In leaves, ARCs are represented by a cessation of cell proliferation (during early development) or the end of cell expansion (when leaves mature), however, in principle any molecular, biochemical, or cellular change that manifests as a result of developmental progression could represent an ARC (Jibran and others 2013). Considering auxin’s prominent role in the regulation of cell division and cell expansion (Mockaitis and Estelle 2004; Perrot-Rechenmann 2010), auxin may exert its control over senescence by affecting the build-up of age-related cellular factors and thus timing of ARCs. It will be interesting in the future to determine what exactly sets the stage to allow bona fide senescence executing mechanisms (for example, involving the ORE1 regulon; Balazadeh and others 2010a, b; Breeze and others 2011) to start their program. The reports discussed here indicate that auxin could be one of the relevant players in this process.
AGI Codes of Genes Discussed in This Article
SAG12 (At5g45890); YUC1 (At1g21430); YUC6 (At5g25620); YUC7 (At2g33230); GH3.1 (At2g14960); GH3.3 (At2g23170); GH3.5 (At4g27260); GH3.6 (At5g54510); SAG201/SAUR36/RAG1 (At2g45210); ORE1/ANAC092 (At5g39610); AtNAP/ANAC029 (At1g69490); WRKY6 (At1g62300).
References
Balazadeh S, Riaño-Pachón DM, Mueller-Roeber B (2008) Transcription factors regulating leaf senescence in Arabidopsis thaliana. Plant Biol (Stuttg) 10(Suppl 1):63–75
Balazadeh S, Siddiqui H, Allu AD, Matallana-Ramirez LP, Caldana C, Mehrnia M, Zanor MI, Köhler B, Mueller-Roeber B (2010a) A gene regulatory network controlled by the NAC transcription factor ANAC092/AtNAC2/ORE1 during salt-promoted senescence. Plant J 62:250–264
Balazadeh S, Wu A, Mueller-Roeber B (2010b) Salt-triggered expression of the ANAC092-dependent senescence regulon in Arabidopsis thaliana. Plant Signal Behav 5:733–735
Balazadeh S, Kwasniewski M, Caldana C, Mehrnia M, Zanor MI, Xue GP, Mueller-Roeber B (2011) ORS1, an H2O2-responsive NAC transcription factor, controls senescence in Arabidopsis thaliana. Mol Plant 4:346–360
Besseau S, Li J, Palva ET (2012) WRKY54 and WRKY70 co-operate as negative regulators of leaf senescence in Arabidopsis thaliana. J Exp Bot 63:2667–2679
Breeze E, Harrison E, McHattie S, Hughes L, Hickman R, Hill C, Kiddle S, Kim YS, Penfold CA, Jenkins D, Zhang C, Morris K, Jenner C, Jackson S, Thomas B, Tabrett A, Legaie R, Moore JD, Wild DL, Ott S, Rand D, Beynon J, Denby K, Mead A, Buchanan-Wollaston V (2011) High-resolution temporal profiling of transcripts during Arabidopsis leaf senescence reveals a distinct chronology of processes and regulation. Plant Cell 23:873–894
Buchanan-Wollaston V, Page T, Harrison E, Breeze E, Lim PO, Nam HG, Lin JF, Wu SH, Swidzinski J, Ishizaki K, Leaver CJ (2005) Comparative transcriptome analysis reveals significant differences in gene expression and signalling pathways between developmental and dark/starvation-induced senescence in Arabidopsis. Plant J 42:567–585
Chae K, Isaacs CG, Reeves PH, Maloney GS, Muday GK, Nagpal P, Reed JW (2012) Arabidopsis SMALL AUXIN UP RNA63 promotes hypocotyl and stamen filament elongation. Plant J 71:684–697
Cheng Y, Dai X, Zhao Y (2006) Auxin biosynthesis by the YUCCA flavin monooxygenases controls the formation of floral organs and vascular tissues in Arabidopsis. Genes Dev 20:1790–1799
Cheng Y, Dai X, Zhao Y (2007) Auxin synthesized by the YUCCA flavin monooxygenases is essential for embryogenesis and leaf formation in Arabidopsis. Plant Cell 19:2430–2439
Cui MH, Ok SH, Yoo KS, Jung KW, Yoo SD, Shin JS (2013) An Arabidopsis cell growth defect factor-related protein, CRS, promotes plant senescence by increasing the production of hydrogen peroxide. Plant Cell Physiol 54:155–167
Diaz C, Purdy S, Christ A, Morot-Gaudry JF, Wingler A, Masclaux-Daubresse CL (2005) Characterization of markers to determine the extent and variability of leaf senescence in Arabidopsis. A metabolic profiling approach. Plant Physiol 138:898–908
Ellis CM, Nagpal P, Young JC, Hagen G, Guilfoyle TJ, Reed JW (2005) AUXIN RESPONSE FACTOR1 and AUXIN RESPONSE FACTOR2 regulate senescence and floral organ abscission in Arabidopsis thaliana. Development 132:4563–4574
Esmon CA, Tinsley AG, Ljung K, Sandberg G, Hearne LB, Liscum E (2006) A gradient of auxin and auxin-dependent transcription precedes tropic growth responses. Proc Natl Acad Sci USA 103:236–241
Fortunati A, Tassone P, Damasso M, Migliaccio F (2010) Neutron irradiation affects the expression of genes involved in the response to auxin, senescence and oxidative stress in Arabidopsis. Plant Signal Behav 5:959–967
Fujiki Y, Ito M, Nishida I, Watanabe A (2000) Multiple signaling pathways in gene expression during sugar starvation. Pharmacological analysis of din gene expression in suspension-cultured cells of Arabidopsis. Plant Physiol 124:1139–1147
Fujiki Y, Yoshikawa Y, Sato T, Inada N, Ito M, Nishida I, Watanabe A (2001) Dark-inducible genes from Arabidopsis thaliana are associated with leaf senescence and repressed by sugars. Physiol Plant 111:345–352
Fukaki H, Nakao Y, Okushima Y, Theologis A, Tasaka M (2005) Tissue-specific expression of stabilized SOLITARY-ROOT/IAA14 alters lateral root development in Arabidopsis. Plant J 44:382–395
Gan S (2005) Hormonal regulation of senescence. In: Davies PJ (ed) Plant hormones: biosynthesis, signal transduction, action. Kluwer, Dordrecht, pp 561–581
Gan SS, Amasino RM (1997) Making sense of senescence—molecular genetic regulation and manipulation of leaf senescence. Plant Physiol 113:313–319
Goda H, Sawa S, Asami T, Fujioka S, Shimada Y, Yoshida S (2004) Comprehensive comparison of auxin-regulated and brassinosteroid-regulated genes in Arabidopsis. Plant Physiol 134:1555–1573
Goda H, Sasaki E, Akiyama K, Maruyama-Nakashita A, Nakabayashi K, Li W, Ogawa M, Yamauchi Y, Preston J, Aoki K, Kiba T, Takatsuto S, Fujioka S, Asami T, Nakano T, Kato H, Mizuno T, Sakakibara H, Yamaguchi S, Nambara E, Kamiya Y, Takahashi H, Hirai MY, Sakurai T, Shinozaki K, Saito K, Yoshida S, Shimada Y (2008) The AtGenExpress hormone and chemical treatment data set: experimental design, data evaluation, model data analysis and data access. Plant J 55:526–542
Guo Y, Cai Z, Gan SS (2004) Transcriptome of Arabidopsis leaf senescence. Plant Cell Environ 27:521–549
Guilfoyle TJ, Hagen G (2007) The authors declare that they have no conflict of interest. Auxin response factors. Curr Opin Plant Biol 10:453–460
Guo Y, Gan S (2005) Leaf senescence: signals, execution, and regulation. Curr Top Dev Biol 71:83–112
Guo Y, Gan S (2006) AtNAP, a NAC family transcription factor, has an important role in leaf senescence. Plant J 46:601–612
Hagen G, Guilfoyle T (2002) Auxin-responsive gene expression: genes, promoters and regulatory factors. Plant Mol Biol 49:373–385
Hickman R, Hill C, Penfold CA, Breeze E, Bowden L, Moore JD, Zhang P, Jackson A, Cooke E, Bewicke-Copley F, Mead A, Beynon J, Wild DL, Denby KJ, Ott S, Buchanan-Wollaston V (2013) A local regulatory network around three NAC transcription factors in stress responses and senescence in Arabidopsis leaves. Plant J 75:26–39
Hou K, Wu W, Gan SS (2013) SAUR36, a small auxin up RNA gene, is involved in the promotion of leaf senescence in Arabidopsis. Plant Physiol 161:1002–1009
Jain M, Tyagi AK, Khurana JP (2006) Genome-wide analysis, evolutionary expansion, and expression of early auxin-responsive SAUR gene family in rice (Oryza sativa). Genomics 88:360–371
Jibran R, Hunter DA, Dijkwel PP (2013) Hormonal regulation of leaf senescence through integration of developmental and stress signals. Plant Mol Biol 82:547–561
Kant S, Bi YM, Zhu T, Rothstein SJ (2009) SAUR39, a small auxin-up RNA gene, acts as a negative regulator of auxin synthesis and transport in rice. Plant Physiol 151:691–701
Kim JI, Sharkhuu A, Jin JB, Li P, Jeong JC, Baek D, Lee SY, Blakeslee JJ, Murphy AS, Bohnert HJ, Hasegawa PM, Yun DJ, Bressan RA (2007) yucca6, a dominant mutation in Arabidopsis, affects auxin accumulation and auxin-related phenotypes. Plant Physiol 145:722–735
Kim JH, Woo HR, Kim J, Lim PO, Lee IC, Choi SH, Hwang D, Nam HG (2009) Trifurcate feed-forward regulation of age-dependent cell death involving miR164 in Arabidopsis. Science 323:1053–1057
Kim JI, Murphy AS, Baek D, Lee SW, Yun DJ, Bressan RA, Narasimhan ML (2011) YUCCA6 over-expression demonstrates auxin function in delaying leaf senescence in Arabidopsis thaliana. J Exp Bot 62:3981–3992
Kim YS, Sakuraba Y, Han SH, Yoo SC, Paek NC (2013) Mutation of the Arabidopsis NAC016 transcription factor delays leaf senescence. Plant Cell Physiol 54(10):1660–1672
Lee M, Jung JH, Han DY, Seo PJ, Park WJ, Park CM (2012) Activation of a flavin monooxygenase gene YUCCA7 enhances drought resistance in Arabidopsis. Planta 235:923–938
Li H, Johnson P, Stepanova A, Alonso JM, Ecker JR (2004) Convergence of signaling pathways in the control of differential cell growth in Arabidopsis. Dev Cell 7:193–204
Li XP, Gan R, Li PL, Ma YY, Zhang LW, Zhang R, Wang Y, Wang NN (2006) Identification and functional characterization of a leucine-rich repeat receptor-like kinase gene that is involved in regulation of soybean leaf senescence. Plant Mol Biol 61:829–844
Lim PO, Kim HJ, Nam HG (2007) Leaf senescence. Annu Rev Plant Biol 58:115–136
Lim PO, Lee IC, Kim J, Kim HJ, Ryu JS, Woo HR, Nam HG (2010) Auxin response factor 2 (ARF2) plays a major role in regulating auxin-mediated leaf longevity. J Exp Bot 61:1419–1430
Lin JF, Wu SH (2004) Molecular events in senescing Arabidopsis leaves. Plant J 39:612–628
Liscum E, Reed JW (2002) Genetics of Aux/IAA and ARF action in plant growth and development. Plant Mol Biol 49:387–400
Matallana-Ramirez LP, Rauf M, Farage-Barhom S, Dortay H, Xue GP, Dröge-Laser W, Lers A, Balazadeh S, Mueller-Roeber B (2013) NAC Transcription factor ORE1 and senescence-induced BIFUNCTIONAL NUCLEASE1 (BFN1) constitute a regulatory cascade in Arabidopsis. Mol Plant 6(5):1432–1452
Miao Y, Laun T, Zimmermann P, Zentgraf U (2004) Targets of the WRKY53 transcription factor and its role during leaf senescence in Arabidopsis. Plant Mol Biol 55:853–867
Mockaitis K, Estelle M (2004) Integrating transcriptional controls for plant cell expansion. Genome Biol 5:245
Narsai R, Law SR, Carrie C, Xu L, Whelan J (2011) In-depth temporal transcriptome profiling reveals a crucial developmental switch with roles for RNA processing and organelle metabolism that are essential for germination in Arabidopsis. Plant Physiol 157:1342–1362
Navabpour S, Morris K, Allen R, Harrison E, A-H-Mackerness S, Buchanan-Wollaston V (2003) Expression of senescence-enhanced genes in response to oxidative stress. J Exp Bot 54:2285–2292
Nemhauser JL, Mockler TC, Chory J (2004) Interdependency of brassinosteroid and auxin signaling in Arabidopsis. PLoS Biol 2:E258
Nemhauser JL, Hong F, Chory J (2006) Different plant hormones regulate similar processes through largely non overlapping transcriptional responses. Cell 126:467–475
Ohto MA, Hayashi S, Sawa S, Hashimoto-Ohta A, Nakamura K (2006) Involvement of HLS1 in sugar and auxin signaling in Arabidopsis leaves. Plant Cell Physiol 47:1603–1611
Okushima Y, Mitina I, Quach HL, Theologis A (2005) AUXIN RESPONSE FACTOR 2 (ARF2): a pleiotropic developmental regulator. Plant J 43:29–46
Paponov IA, Paponov M, Teale W, Menges M, Chakrabortee S, Murray JA, Palme K (2008) Comprehensive transcriptome analysis of auxin responses in Arabidopsis. Mol Plant 1:321–337
Perrot-Rechenmann C (2010) Cellular responses to auxin: division versus expansion. Cold Spring Harb Perspect Biol 2:a001446
Pourtau N, Jennings R, Pelzer E, Pallas J, Wingler A (2006) Effect of sugar-induced senescence on gene expression and implications for the regulation of senescence in Arabidopsis. Planta 224:556–568
Rauf M, Arif M, Dortay H, Matallana-Ramírez LP, Waters MT, Gil Nam H, Lim PO, Mueller-Roeber B, Balazadeh S (2013) ORE1 balances leaf senescence against maintenance by antagonizing G2-like-mediated transcription. EMBO Rep 14:382–388
Richter R, Behringer C, Müller IK, Schwechheimer C (2010) The GATA-type transcription factors GNC and GNL/CGA1 repress gibberellin signaling downstream from DELLA proteins and PHYTOCHROME-INTERACTING FACTORS. Genes Dev 24:2093–2104
Richter R, Behringer C, Zourelidou M, Schwechheimer C (2013) (2013) Convergence of auxin and gibberellin signaling on the regulation of the GATA transcription factors GNC and GNL in Arabidopsis thaliana. Proc Natl Acad Sci USA 110(32):13192–13197
Schippers JHM, Jing HC, Hille J, Dijkwel PP (2007) Developmental and hormonal control of leaf senescence. In: Gan S (ed) Senescence processes in plants. Blackwell, Oxford, pp 145–170
Schruff MC, Spielman M, Tiwari S, Adams S, Fenby N, Scott RJ (2006) The AUXIN RESPONSE FACTOR 2 gene of Arabidopsis links auxin signalling, cell division, and the size of seeds and other organs. Development 133:251–261
Spartz AK, Lee SH, Wenger JP, Gonzalez N, Itoh H, Inzé D, Peer WA, Murphy AS, Overvoorde PJ, Gray WM (2012) The SAUR19 subfamily of SMALL AUXIN UP RNA genes promote cell expansion. Plant J 70:978–990
Stamm P, Kumar PP (2013) Auxin and gibberellin responsive Arabidopsis SMALL AUXIN UP RNA36 regulates hypocotyl elongation in the light. Plant Cell Rep 32(6):759–769
Staswick PE, Serban B, Rowe M, Tiryaki I, Maldonado MT, Maldonado MC, Suza W (2005) Characterization of an Arabidopsis enzyme family that conjugates amino acids to indole-3-acetic acid. Plant Cell 17:616–627
Sun J, Qi L, Li Y, Chu J, Li C (2012) PIF4-mediated activation of YUCCA8 expression integrates temperature into the auxin pathway in regulating Arabidopsis hypocotyl growth. PLoS Genet 8:e1002594
Trivellini A, Jibran R, Watson LM, O’Donoghue EM, Ferrante A, Sullivan KL, Dijkwel PP, Hunter DA (2012) Carbon deprivation-driven transcriptome reprogramming in detached developmentally arresting Arabidopsis inflorescences. Plant Physiol 160:1357–1372
Tsuchisaka A, Yu G, Jin H, Alonso JM, Ecker JR, Zhang X, Gao S, Theologis A (2009) A combinatorial interplay among the 1-aminocyclopropane-1-carboxylate isoforms regulates ethylene biosynthesis in Arabidopsis thaliana. Genetics 183:979–1003
van der Graaff E, Schwacke R, Schneider A, Desimone M, Flügge UI, Kunze R (2006) Transcription analysis of Arabidopsis membrane transporters and hormone pathways during developmental and induced leaf senescence. Plant Physiol 141:776–792
van Doorn WG (2004) Is petal senescence due to sugar starvation? Plant Physiol 134:35–42
van Doorn WG (2008) Is the onset of senescence in leaf cells of intact plants due to low or high sugar levels? J Exp Bot 59:1963–1972
Watanabe M, Balazadeh S, Tohge T, Erban A, Giavalisco P, Kopka J, Mueller-Roeber B, Fernie AR, Hoefgen R (2013) Comprehensive dissection of spatiotemporal metabolic shifts in primary, secondary, and lipid metabolism during developmental senescence in Arabidopsis. Plant Physiol 162:1290–1310
Waters MT, Moylan EC, Langdale JA (2008) GLK transcription factors regulate chloroplast development in a cell-autonomous manner. Plant J56:432–444
Waters MT, Wang P, Korkaric M, Capper RG, Saunders NJ, Langdale JA (2009) GLK transcription factors coordinate expression of the photosynthetic apparatus in Arabidopsis. Plant Cell 21:1109–1128
Wingler A, Purdy S, MacLean JA, Pourtau N (2006) The role of sugars in integrating environmental signals during the regulation of leaf senescence. J Exp Bot 57:391–399
Wingler A, Masclaux-Daubresse C, Fischer AM (2009) Sugars, senescence, and ageing in plants and heterotrophic organisms. J Exp Bot 60:1063–1066
Wu A, Allu AD, Garapati P, Siddiqui H, Dortay H, Zanor MI, Asensi-Fabado MA, Munné-Bosch S, Antonio C, Tohge T, Fernie AR, Kaufmann K, Xue GP, Mueller-Roeber B, Balazadeh S (2012) JUNGBRUNNEN1, a reactive oxygen species-responsive NAC transcription factor, regulates longevity in Arabidopsis. Plant Cell 24:482–506
Xu F, Meng T, Li P, Yu Y, Cui Y, Wang Y, Gong Q, Wang NN (2011) A soybean dual-specificity kinase, GmSARK, and its Arabidopsis homolog, AtSARK, regulate leaf senescence through synergistic actions of auxin and ethylene. Plant Physiol 157:2131–2153
Yang SD, Seo PJ, Yoon HK, Park CM (2011) The Arabidopsis NAC transcription factor VNI2 integrates abscisic acid signals into leaf senescence via the COR/RD genes. Plant Cell 23:2155–2168
Yoon HK, Kim SG, Kim SY, Park CM (2008) Regulation of leaf senescence by NTL9-mediated osmotic stress signaling in Arabidopsis. Mol Cells 25:438–445
Yoshida S (2003) Molecular regulation of leaf senescence. Curr Opin Plant Biol 6:79–84
Zhang K, Gan SS (2012) An abscisic acid-AtNAP transcription factor-SAG113 protein phosphatase 2C regulatory chain for controlling dehydration in senescing Arabidopsis leaves. Plant Physiol 158:961–969
Zhang H, Zhou C (2013) Signal transduction in leaf senescence. Plant Mol Biol 82:539–545
Zhang K, Xia X, Zhang Y, Gan SS (2012) An ABA-regulated and Golgi-localized protein phosphatase controls water loss during leaf senescence in Arabidopsis. Plant J 69:667–678
Zhao Y, Christensen SK, Fankhauser C, Cashman JR, Cohen JD, Weigel D, Chory J (2001) A role for flavin monooxygenase-like enzymes in auxin biosynthesis. Science 291:306–309
Zhao Y, Dai X, Blackwell HE, Schreiber SL, Chory J (2003) SIR1, an upstream component in auxin signaling identified by chemical genetics. Science 301:1107–1110
Zhou Q, Yu Q, Wang Z, Pan Y, Lv W, Zhu L, Chen R, He G (2013) Knockdown of GDCH gene reveals reactive oxygen species-induced leaf senescence in rice. Plant Cell Environ 36(8):1476–1489. doi:10.1111/pce.12078
Acknowledgments
We thank the Deutsche Forschungsgemeinschaft (DFG) for funding (FOR 948; MU 1199/14-1 and 14-2, BA4769/1-2). We also thank the University of Potsdam and the Max-Planck Institute of Molecular Plant Physiology, Potsdam-Golm, for supporting our research.
Conflict of interest
The authors do not declare any conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
344_2013_9398_MOESM1_ESM.xlsx
Table S1. Lists of senescence-associated genes (SAGs), senescence-down-regulated genes (SDGs), dark-regulated genes, H2O2-responsive genes, and auxin-responsive genes. (XLSX 467 kb)
Rights and permissions
About this article
Cite this article
Mueller-Roeber, B., Balazadeh, S. Auxin and Its Role in Plant Senescence. J Plant Growth Regul 33, 21–33 (2014). https://doi.org/10.1007/s00344-013-9398-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00344-013-9398-5