
Abstract
Sugarcane varieties (Saccharum spp. hybrids) that accumulate high levels of sucrose at the start of the harvest season are of considerable commercial interest. Our understanding of the factors that contribute to early sucrose accumulation in these varieties is limited. In this study we used the plant hormone ethylene to investigate the relationship between growth and early sucrose accumulation in sugarcane. The sugarcane variety KQ228 was exposed to a low concentration of the ethylene-forming compound 2-chloroethylphosphonic acid (CEPA) for a prolonged duration commencing from shoot emergence. The changes in sucrose accumulation and plant growth were investigated. Results from two glasshouse experiments revealed that the CEPA-treated plants accumulated a significantly higher amount of sucrose in their primary culm 2 and 3½ months post-germination. The treated plants had taller primary culms with many smaller internodes, smaller leaves, and a higher photosynthetic rate. Despite producing smaller internodes, treated culms were comparable in fresh weight and volume to the controls due to the compensating effect of faster internode formation. We identified three factors that may have contributed to the early accumulation of more sucrose in the treated culm: (1) the specific leaf area of young leaves was greater indicating efficient diversion of photoassimilate to sink tissue, (2) internode formation was initiated earlier, and (3) internodes continued to form at a faster rate. Consequently, a greater proportion of the internodes in the treated sugarcane matured earlier and began filling with sucrose sooner. The higher reducing sugar level in the apical region of the culm probably contributed to faster internode development. This coincided with elevated vacuolar and cell wall acid invertase gene expression that increased sucrose turnover in the vacuole and increased apoplastic uptake of reducing sugars. These findings extend our understanding of how some sugarcane varieties can naturally accumulate a high level of sucrose early in the season.
was exposed to a low concentration of the ethylene-forming compound 2-chloroethylphosphonic acid (CEPA) for a prolonged duration commencing from shoot emergence. The changes in sucrose accumulation and plant growth were investigated. Results from two glasshouse experiments revealed that the CEPA-treated plants accumulated a significantly higher amount of sucrose in their primary culm 2 and 3½ months post-germination. The treated plants had taller primary culms with many smaller internodes, smaller leaves, and a higher photosynthetic rate. Despite producing smaller internodes, treated culms were comparable in fresh weight and volume to the controls due to the compensating effect of faster internode formation. We identified three factors that may have contributed to the early accumulation of more sucrose in the treated culm: (1) the specific leaf area of young leaves was greater indicating efficient diversion of photoassimilate to sink tissue, (2) internode formation was initiated earlier, and (3) internodes continued to form at a faster rate. Consequently, a greater proportion of the internodes in the treated sugarcane matured earlier and began filling with sucrose sooner. The higher reducing sugar level in the apical region of the culm probably contributed to faster internode development. This coincided with elevated vacuolar and cell wall acid invertase gene expression that increased sucrose turnover in the vacuole and increased apoplastic uptake of reducing sugars. These findings extend our understanding of how some sugarcane varieties can naturally accumulate a high level of sucrose early in the season.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Commercial sugarcane varieties (interspecific hybrids of Saccharum spp.) can yield up to 35 t sucrose ha−1 year−1 under optimal conditions (Moore and others 1997). A mature sugarcane culm has the capacity to store up to 62% sucrose on a dry weight basis or 25% sucrose on a fresh weight basis (Bull and Glasziou 1963). The sugarcane culm consists of consecutive internodes and nodes, the latter being the attachment point of a leaf. A new internode is initiated shortly after the formation of a new leaf primordium at the apical meristem. The development of the internode consists broadly of two phases, maturation and ripening (Alexander 1973). Maturity is an indicator of realized growth potential, while ripeness is an indicator of realized sucrose storage potential. As the new internode matures it grows larger until a final size is attained. During this period, relatively more photoassimilate is used to build the internode than is stored as sucrose. Sucrose accumulation becomes the main activity once internode growth is completed. This phase is characterized by faster sucrose accumulation in the storage parenchyma. Under normal growth conditions the sugarcane culm ripens acropetally as consecutively formed internodes ripen.
Controlling ripening is an important aspect of sugarcane crop management. To maximize economic return from a sugarcane crop, it must be harvested at the point in its life cycle when its sucrose level is highest. If the crop is harvested too early or too late the sucrose yield will be low. Suboptimal harvest scheduling also leads to poor juice purity (that is, high reducing sugar and low sucrose) resulting in a lower sucrose recovery at the sugar mill. Often harvesting does not coincide with the period when the crop sucrose content is highest because of capacity constraints imposed by harvest machinery and the sugar mill. In such situations, commencing the harvest early is preferable to finishing the harvest late, as the crop may lodge and the sucrose concentration deteriorate toward the latter part of the season (McDonald 2006). Growing varieties that accumulate higher levels of sucrose early (referred to as early-ripening varieties) is advantageous as it enables the mill to process a larger crop by bringing forward the start of the harvest season or to increase sucrose production early in the season if the start date remains unchanged (Cox and others 1994). Use of early-ripening varieties has ensured it is commercially viable to grow sugarcane in subtropical regions such as Louisiana where the climate limits the length of the growing season (Breaux 1984).
Other practices that are used to accelerate sugarcane ripening include withholding irrigation before harvest (Gosnell and Lonsdale 1974) and applying various chemical ripeners (Morgan and others 2007). Most chemical ripeners act by temporarily arresting vigorous growth in the immature upper internodes of the culm thereby causing accelerated deposition of sucrose in those internodes. Of the numerous chemical ripeners used, the plant growth regulator ethylene is one that has proven to be effective on many varieties of sugarcane (Donaldson and Van Staden 1989). It is applied to the crop as a foliar spray containing the active ingredient 2-chloroethylphosphonic acid (CEPA), which readily decomposes into ethylene. There is a mounting body of evidence indicating that ethylene is a concentration-dependent biphasic growth modulator in higher plants (Pierik and others 2006). At high concentrations it may inhibit growth, whereas at low concentrations it may stimulate growth. The response to CEPA also depends upon the stage of plant development (Van Andel and Verkerke 1978). Thus, CEPA is also used to enhance the sprouting of seed cane, to promote vigorous tillering (Eastwood 1979; Solomon and others 1998), and to prevent flowering (Moore and Osgood 1989).
CEPA-induced ripening is usually achieved by applying a single high dose of the chemical to a sugarcane crop toward the end of the growing season. The magnitude of the ripening response depends upon the maturity of the tissue (Rostron 1973). The changes occurring after treatment are mainly restricted to the upper internodes of the culm. Therefore, the treatment principally affects immature tissue where the juice purity is low. It has been demonstrated that three applications of CEPA at 2-month intervals produce a larger increase in sucrose concentration than a single treatment (Rostron 1977). This is not surprising as ethylene induces rapid but short-term responses. A sustained response can be achieved by continuous exposure to the hormone (Warner and Leopold 1971; Fiorani and others 2002).
There is compelling evidence linking growth and sucrose accumulation in sugarcane. Assimilate is sequestered by the plant as structural carbohydrates (in leaves, culms, and roots), dissipated through respiration or stored as sucrose (in the parenchyma tissue of the culm). The allocation of this assimilate can be altered by manipulating respiration (Groenewald and Botha 2008) or imposing water stress (Inman-Bamber and others 2008). In the latter case, a greater proportion of photoassimilate is stored as sucrose because water stress has a greater impact on growth than it has on photosynthesis. Various forms of growth suppression favour the partitioning of photoassimilate to sucrose storage. Shading or partially defoliating sugarcane increases the proportion of sucrose in total dry matter (Pammenter and Allison 2002). Transferring sugarcane from a high- to a low-temperature environment causes an increase in sucrose content (Glasziou and others 1965). Therefore, it is conceivable that some varieties accumulate sucrose faster as a natural response to unfavourable growing conditions such as cooler temperatures or soil nitrogen and moisture deficits. Some sugarcane studies have linked certain underlying growth characteristics with faster sucrose accumulation. Smaller leaves have been associated with early-ripening sugarcane (Nayamuth and others 1999). The culms of some early-ripening varieties elongate faster (Mamet and Galwey 1999), start elongating earlier, and have more internodes that are characteristically shorter (Nayamuth and others 1999). This suggests that faster culm development may facilitate earlier accumulation of sucrose. However, other studies found no relationship between culm development and ripening (Lingle 1997). The inconsistency in the findings between previous studies may be due to the practice of comparing different sugarcane genotypes where the diverse genetic backgrounds tend to confound the interpretation of the results.
In this study we modified the early growth of a single sugarcane genotype by applying a low concentration of CEPA to the plant for a prolonged duration beginning from shoot emergence. The objective was to test whether this growth modification would enhance the sucrose accumulation rate in the culm. Our experimental approach involved evaluating the relationship between growth and sucrose accumulation rate in CEPA-treated sugarcane. We also attempted to relate this to the expression pattern of some relevant genes. Sucrose accumulation in sugarcane internodes is the net result of a synthesis and degradation cycle involving the sucrose-synthesizing enzyme sucrose phosphate synthase (SPS) and the sucrose-hydrolysing enzyme vacuolar acid invertase (VAI) (Zhu and others 1997). Although the enzymes sucrose synthase and cytosolic neutral invertase would be expected to contribute to the sucrose futile cycle, it was reported that these enzyme activities did not vary between high and low sucrose genotypes or with internode maturity (Zhu and others 1997). Consequently, only the SPS and VAI genes were included in our gene expression study. The SPS chosen was a member of the distinctive D family that is found only in monocots (Castleden and others 2004) and is abundant in sugarcane (Grof and others 2006). Expression of the apoplastic cell wall invertase gene CWI was also examined because the enzyme that it encodes controls the flow of sucrose from conducting tissue to growing cells (Roitsch and others 2003). The genes for cellulose synthase, CES, and β-expansin, EXP, were also examined. Cellulose synthase converts hexoses into one of the principal cell wall building blocks (namely, cellulose) while β-expansin induces slippage of cellulose microfibrils in the cell wall to enable turgor-driven extension of the wall. The gene ShCESA6 from the cellulose synthase gene family was selected because it is abundant in maturing sugarcane stems (Casu and others 2007). Finally, expression of the gene pyrophosphate-dependent d-fructose 6-phosphate phosphotransferase, PFP, was considered in the study because the enzyme that it encodes is directly correlated to total respiration in the internode and inversely correlated to sucrose content (Whittaker and Botha 1999). This study demonstrated that the growth and developmental changes arising from CEPA treatment permitted faster and higher sucrose accumulation in the culm. The findings provided valuable insight into how natural early ripening may occur.
Materials and Methods
Two glasshouse experiments were conducted in this study; the first occurred between 24 October 2007 and 15 February 2008 and the second occurred between 16 October 2008 and 10 February 2009. Both experiments were harvested at 3½ mpg (month post-germination). The second experiment was used to compare the timing of primary culm internode formation, the total primary shoot leaf area, and SLA (specific leaf area).
Planting Material
Vegetative cuttings (one-eye setts) were obtained from mature field-grown culms of the sugarcane cultivar KQ228. Each one-eye sett was propagated in a 20-cm diameter pot containing a soil substitute comprising equal parts perlite and vermiculite (Chillagoe Perlite, Mareeba, Queensland, Australia). Two groups of clonal replicates were established in a naturally ventilated glasshouse. One group was hand sprayed with a solution of 50 mg l−1 of CEPA (Alfa Aesar, Lancashire, England) in 0.5% v/v surfactant (Teepol 610S, Sigma-Aldrich, St Louis, MO, USA) and the other group was hand sprayed with water containing 0.5% v/v surfactant only (control). As the half-life of CEPA at room temperature is only 24 h (Domir and Foy 1978) and the experiment aimed to achieve a sustained response, the plants were sprayed on Tuesday and Friday each week. All vegetative cuttings germinated approximately 12 days after planting and spraying commenced 7 days post-germination. The pots were watered at 0500, 1200, and 1700 h daily for 10 min at a rate of 2 l h−1 by drippers connected to a Raindial RD-600 Irrigation Controller (Irritrol Systems, Riverside, CA, USA). Sixty grams of Osmocote (Scotts Australia Pty Ltd, Baulkam Hills, NSW, Australia), a slow-release fertilizer, was added to each pot.
Timing of Leaf Emergence and Internode Expansion
An internode begins expanding inside the leaf bundle when the corresponding leaf attached to it becomes fully expanded. Thus, the commencement of internode expansion can be determined nondestructively by noting the appearance date of each new fully expanded leaf. A leaf was deemed fully expanded when its ligule was completely visible. Data collection for each primary culm commenced after the appearance of a leaf that was greater than 2 cm in length and continued until the trial was harvested. The average number of internodes produced by the primary culms in experiments 1 and 2 were equivalent. As the experiments were of similar duration, we inferred that the internode formation rates in the two experiments were also equivalent. The data from experiment 2 was therefore used to estimate the age of the internodes produced in the first experiment.
Sampling and Phenotyping
Clonal replicates were randomly chosen for harvest from the CEPA-treated and control groups. The primary culm, leaf components (top, lamina, and sheath), and tillers were weighed separately. The diameter (midway along the internode) and length (distance between consecutive leaf scars) of each internode was measured using a digital vernier calliper. The internode volume was estimated from the length and diameter by assuming the internode was cylindrical. The total culm volume was estimated by summing the volume of all internodes in the culm. A 4-mm diameter cylindrical tissue sample was excised transversely from a position that was midway along each internode using a cork borer. These samples were used to determine the sugar concentration in the internode. Leaves and internodes were sequentially numbered from the top to the bottom of the culm according to the system suggested by Kuijper (Van Dillewijn 1952). Internode four (the internode above the point of attachment of the fourth-last fully expanded leaf) was collected for RNA extraction. The remaining tissue from the primary culm was collected for average sugar, average insoluble solids, and cellulose analysis. For each green leaf, lamina mass, length and maximum width, and sheath mass and length were measured. A leaf was regarded as green if more than 50% of the lamina was green. The second-last, fourth-last, and sixth-last fully expanded leaves were collected to measure lamina area, dry weight, and sugar concentration. All collected samples were stored at −80°C until processed.
Leaf Area Measurement
The surface area of a leaf lamina (A) was determined by the leaf shape factor method (A = ϕ LW m, where ϕ is the shape factor, L is the lamina length, and W m is the maximum width). Due to the considerable difference in leaf size between the treated and control plants, it was considered prudent to determine the shape factor independently for each group. Leaves one and seven from each plant were used to calibrate the shape factor.
Leaf shape can be depicted by a polynomial equation describing the leaf width as a function of distance from the ligule (Stewart and Dwyer 1999). The shape factor is calculated from the coefficients of that polynomial equation. Width measurements were taken at 10-cm intervals along the leaf length for the control leaves and at 5-cm intervals for the CEPA-treated leaves. A fourth-order polynomial equation was found to be the best fit for the shape of sugarcane leaves. Comparison of the shape factors by analysis of variance indicated that there was no significant difference between leaves one and seven or between the control and CEPA groups (results not shown). Therefore, an average shape factor of 0.6 was used in the area equation.
Sugar, Insoluble Solids, and Cellulose Measurement
Whole culms were shredded in a blender and then dried to a constant weight. The dry material was subsequently ground into powder in a ring mill (Rocklabs, Auckland, New Zealand) with a steel mill head comprising a single 20-cm discus in a 25-cm bowl. Internode samples and whole leaves were dried to a constant weight. Internode samples were processed as described previously (Fong Chong and others 2007) and leaf samples were ground to powder in a Retsch mixer mill (MM300, Retsch GmbH & Co., Hann, Germany). The total sugars in subsamples of the leaf and whole culm powder were extracted twice into 0.02% w/v sodium azide (10 ml g−1 tissue) by incubating each extraction at 80°C for 3 h. Sucrose, glucose, and fructose in the filtered tissue extracts were quantified by HPLC as described previously (Fong Chong and others 2007) except that the HPLC column used was a Shodex 8 × 300 mm SUGAR KS-801 analytical column (Showa Denko, Tokyo, Japan). The HPLC column was operated at 65°C and eluted with water as the mobile phase at a flow rate of 0.9 ml min−1. The sugar yield per culm was estimated by calculating the product of the average sugar concentration (mg g−1 FW) and the culm fresh weight.
The insoluble solids remaining after the extraction of leaves or whole culm were dried to a constant weight. The final dry weight was used to calculate the insoluble fraction in the original sample. The cellulose content was then measured from 40 mg of the dried insoluble fraction using the acetic-nitric method (Updegraff 1969). The cellulose standards used in the assay were prepared from Avicel PH-101 (Sigma-Aldrich, St. Louis, MO, USA).
Photosynthesis Measurement
Photosynthesis-related measurements for the fourth-last fully expanded leaf were measured in situ (24 h after CEPA treatment) using the LI-6400 portable photosynthesis system (LI-COR Biosciences, Inc., Lincoln, NE, USA) at 3 mpg in the first experiment. Measurements were taken under a constant irradiance of 2000 μmol m−2 s−1 from a leaf area of 6 cm2 located approximately one third of the leaf length away from the distal end of the leaf. The airflow rate through the leaf chamber was preset to 400 μmol s−1. Photosynthetic rates were calculated on the basis of an adaxial-to-abaxial stomatal ratio of 0.5 (Van Dillewijn 1952). Leaf CO2 response curves were generated by assessing the photosynthetic rate at various reference CO2 levels (400, 300, 200, 100, 40, 600, 900, 1200, and 1500 μmol CO2 mol−1 in that sequence). An asymptotic function of the form J = J o + a(1 − e−bCi) was fitted to the photosynthetic rate (J) and intercellular CO2 concentration (C i) data. The CO2-saturated photosynthetic rate (J max = J o + a) and carboxylation efficiency [slope of the curve at J = 0, CE = b(J o + a)] were evaluated from this equation.
In situ Stomata Characterization
A silicon imprint of the adaxial and abaxial lamina surfaces of the fourth-last fully expanded leaf was prepared in situ from a position that was midway along the lamina (Weyers and Johansen 1985). This method supposedly prevents the stomata from moving during measurement. The material used to make the silicon mould was Xantopren VL Plus with Optosil-Xantopren universal activator (Heraeus Kulzer GmbH, Germany). The negative lamina imprint in the silicon was then transferred to a layer of transparent fingernail varnish that was applied to the surface of a microscope slide. The positive lamina imprint in the varnish was examined by bright-field microscopy (Olympus BX-50 System microscope, Olympus Optical Co. Ltd, Japan). The number of stomata observed in the 0.58-mm2 microscope field of view was counted. In addition, the length and width of the stomatal aperture were measured on the adaxial and abaxial surfaces. Image resolution under light microscopy was problematic, thus, only 72 stomata from six CEPA-treated replicates and 101 stomata from six control replicates were identified as suitable for evaluation.
Chlorophyll Measurement
One half of the lamina from the fifth-last fully expanded leaf from plants in the first experiment was homogenized in liquid nitrogen by mortar and pestle. Fifty milligrams of the homogenate was immediately collected for the measurement of chlorophyll. The cold methanol method was used to quantify the chlorophyll content (Porra and others 1989).
Real-Time Quantitative PCR (RTqPCR)
Between 4 and 5 g of pith tissue was excised from internode four from plants in the first experiment. The total RNA was extracted from the tissue using a modified acid guanidinium-phenol-chloroform method (Chomczynski and Sacchi 1987). The RNA was treated with DNase I (Qiagen Pty Ltd, Doncaster, Victoria, Australia) and purified through an RNeasy column (Qiagen Pty Ltd). The absence of contaminating genomic DNA was confirmed by PCR using a diagnostic primer set flanking the intron of the sugarcane sucrose synthase gene (NCBI accession No. AY118266). The ImProm-II reverse transcription system (Promega Corp., Madison, WI, USA) and random hexamer primers were used to synthesize cDNA from 1 μg of total RNA.
Primers were designed from sugarcane target and reference gene sequences obtained from either the National Centre for Biotechnology Information (NCBI) or The Institute for Genomic Research (TIGR) (Quackenbush and others 2000). The primer design software Primer3Plus (Untergasser and others 2007) was utilized. The reference gene chosen was β-actin (Iskandar and others 2004) (ACT, NCBI accession No. AY742219, forward ACACTGTGCCCATTTACGAA, reverse CAGCAAGATCCAAACGAAGA, 63-bp amplicon). The target genes were sucrose phosphate synthase (SPS, NCBI accession No. EU278617, forward CCCGAACATTGCAAGAATTA, reverse CTCCGCTCCTCTCTGTTACC, 83-bp amplicon), vacuolar acid invertase (VAI, NCBI accession No. AY302083, forward CAGAGGAACTGGATGAACGA, reverse GTCCGGGTTGTATTGGTAGAA, 81-bp amplicon), cellulose synthase (CES, TIGR accession No. TC56553, forward CAAAGGTTCTGCACCTCTCA, reverse GCAGTGGTTGCTGAAGAAAA, 94-bp amplicon), β-expansin (EXP, TIGR accession No. TC65007, forward CCTGCTACCAGATACGATGC, reverse GCCACCGGGTAGTAGTTCAT, 91-bp amplicon), pyrophosphate-dependent d-fructose 6-phosphate phosphotransferse β-subunit (PFP, NCBI accession No. EF103287, forward GCCTGAATCAAAGGAGCTGT, reverse GGGATCCCTTTCAAGCATAA, 73-bp amplicon), and cell wall invertase (CWI, NCBI accession No. AY302084, forward TCTGTACAAGCCAACCTTCG, reverse CCGCTTGAAATGTCAATGTC, 51-bp amplicon).
The SensiMix dT SYBR® Green I detection system (Quantace Pty Ltd, Alexandria, Australia) was used for RTqPCR. The reference gene and target gene were measured in every sample. Each reaction comprised 4 μl of template (equivalent to 20 ng cDNA), 6 μl of 2 × SensiMix, and 2 μl of 3 μM primer mix. Individual reactions were prepared in triplicate by a CAS-1200 liquid handling robot (Corbett Research Pty Ltd, Mortlake, NSW, Australia). RTqPCR was performed in a Rotagene 3000 PCR machine (Corbett Research Pty Ltd). The cycling conditions used were 95°C for 10 min then 40 cycles of 95°C for 15 s and 60°C for 1 min. Amplicon sizes were verified by agarose gel electrophoresis. The absence of nonspecific products was confirmed by melt curves.
Transcript abundance was calculated from the real-time PCR curves using the relative expression method (Pfaffl 2001). Actual PCR efficiencies were used in the calculation. Efficiencies were determined for each primer pair by analysing the amplification of different template amounts spanning a six-log dilution range. The efficiencies obtained were SPS, 0.91; VAI, 0.89; EXP, 0.95; CES, 0.91; PFP, 1.0; CWI, 0.96; and ACT, 1.0.
Statistical Analysis
Statistically significant differences between CEPA treatment and control were identified by the two-sample Student’s t test or analysis of variance. Both were performed in Statistix 8 for Windows (Analytical Software, Tallahassee, FL, USA). SigmaPlot 9.01 for Windows (Systat Software, Inc., Richmond, CA, USA) was used to perform regression analysis.
Results
Two experiments were performed in this study. The first experiment examined the effect of CEPA treatment on certain morphological, compositional, and photosynthetic characteristics in sugarcane after 3½ mpg. In addition, the expression of particular genes in the culm was investigated. The data that were collected from this experiment indicated that the internode appearance rate, total leaf area, and the SLA were involved in the sugarcane ethylene response. Subsequently, a second experiment was conducted the following year over the same period on a new group of plants to specifically obtain these data. The measurements were collected 3½ mpg in the second group of plants. Some measurements were also collected 2 mpg to verify that the growth behaviour was consistent with the plants in the first experiment.
CEPA-Treated Sugarcane Culms Accumulated More Sucrose
Sugarcane plants treated with 50 mg l−1 CEPA displayed significantly higher average sucrose concentrations in their primary culms by 2 mpg compared to control plants sprayed with water plus surfactant (Table 1). At 3½ mpg, the difference in sucrose concentration between control and CEPA-treated plants was still significant. The average sucrose concentration of the whole culm was approximately 45% (2 mpg) and 32% (3½ mpg) higher in the treated plants compared to the control plants. This was equivalent to approximately 423% (2 mpg) and 48% (3½ mpg) higher average sucrose yield per culm in treated plants (Table 1).
Average glucose and fructose concentrations in the primary culm of CEPA-treated plants were significantly higher at 2 mpg but not at 3½ mpg (Table 1). Insoluble solids, which largely comprise plant fibre, were not significantly different at 2 and 3½ mpg (Table 1). No difference was observed at 3½ mpg for cellulose (Table 1), which is the most abundant component of the fibre, typically comprising 40–50% of the fibre dry weight. The CEPA-treated plants contained significantly less moisture in their culms after 3½ mpg (Table 1).
CEPA-Treated Sugarcane Culms Accumulated Sucrose Faster
The sucrose concentration profiles along the primary culm at 3½ mpg indicated that sucrose accumulated faster in the CEPA-treated group (Fig. 1a). Unlike conventional ripening-inducing treatments (which are applied to older plants and predominantly increase sucrose concentration in the immature upper internodes), the ripening effect was not restricted to the upper regions of the culm. The sucrose titre seemed to plateau at a maximum value in older internodes beginning at the tenth youngest internode for CEPA-treated plants and at the seventh youngest internode for control plants (Fig. 1a). This implied that possibly eight older internodes (including internode ten) had reached their maximum sucrose storage potential in CEPA-treated plants. As the maximum sucrose level was lower in the controls compared to the treated plants, the older control internodes may be still filling with sucrose.

The sucrose (a) and reducing sugar (b) concentration in each internode along the primary culm of CEPA-treated sugarcane at 3½ mpg. Each internode is numbered and is plotted against its chronological age. Control (filled circle), CEPA (open circle). The data plotted are the mean ± standard error of measurements collected from six independent plants per treatment
Glucose and fructose concentration profiles along the culm were equivalent and hence they are reported as a reducing sugar pool (Fig. 1b). At the top of the culm, the reducing sugar concentration was higher in the treated cane than in the control. Toward the middle and bottom of the culm the concentrations were lower in the treated cane.
CEPA Treatment Induced Early Internode Development
The CEPA-treated plants produced 4 ± 0.3 leaves and the controls 5.3 ± 0.3 leaves before forming their first internode. Although this difference was small it was still statistically significant (P < 0.05, n = 6 per treatment), indicating that on the basis of leaf development, the treated plants were marginally younger when they produced their first internode. As a result, the first internode on the CEPA-treated primary culms formed approximately 15 days prior to the first internode forming on the controls (Fig. 2a). This equated to the formation of three internodes before the first internode appeared in the control group (Fig. 2a). The average phyllochron (defined as the time interval between the formation of consecutive fully expanded leaves) of the CEPA-treated group (4.2 ± 0.1 days) was significantly less than that of the control (6.1 ± 0.2 days, P < 0.01, n = 6 per treatment; Fig. 2a). This led to the production of internodes at a faster rate.

The timing of (a) internode appearance on the primary culm and (b) tiller formation evaluated for an extended duration after shoot germination. Control (filled circle), CEPA (open circle). The data plotted are the mean ± standard error of measurements collected from 6 independent plants per treatment for (a) and 12 independent plants per treatment for (b). The lines of best fit are (a) control, y = 0.167x − 6.888, r 2 = 0.998, P < 0.0001; CEPA, y = 0.243x − 7.288, r 2 = 0.996, P < 0.0001 (where x is days post-germination and y is number of internode formed) and (b) control, y = 0.036x − 0.385, r 2 = 0.89, P < 0.0001; CEPA, y = 0.091x − 1.836, r 2 = 0.98, P < 0.0001 (where x is days post-germination and y is average number of tillers per pot)
CEPA-Treated Sugarcane Culms Were Taller and Contained More Internodes
The primary culms of CEPA-treated sugarcane were taller and contained approximately twice as many internodes as the controls (Table 2). Many of the later-formed internodes were shorter (Fig. 3a) and all were thinner (Fig. 3b). As a result, the volume and fresh weight of each internode was generally less (Fig. 3c, d). At 2 mpg the fresh weight of the treated primary culms was approximately fourfold larger than that of the control (Table 2). The extent of internode elongation in treated and control culms at 2 mpg is illustrated in Fig. 4a. However, by 3½ mpg the fresh weight and volume of the primary culms in the control group had drawn even with those of the treated group. Over 3½ months of growth, the treated primary culms accumulated mass (on both fresh and dry weight basis) and volume at a slower rate than the controls. This was indicated by the fact that their final mass and volume were equivalent to the controls but their growth duration was longer (since their first internode formed earlier).

The length (a), diameter (b), volume (c), and fresh weight (d) of each internode along the primary culm of CEPA-treated sugarcane at 3½ mpg. Each internode is numbered and is plotted against its chronological age. Control (filled circle), CEPA (open circle). The data plotted are the mean ± standard error of measurements collected from six independent plants per treatment

The physical appearance of the primary culm in CEPA-treated sugarcane. (a) The extent of internode elongation was greater in the CEPA-treated culms (left of the cm scale) than in the control culms (right of the cm scale) at 2 mpg. (b) Advanced lateral bud and adventitious root development in a CEPA-treated culm at 3½ mpg
The growth behaviour of the first few internodes dictated the final size of the internodes. Control internodes three to seven were longer than the corresponding internodes on the treated culms. This was mainly due to the elongation that occurs as internode two develops into internode three in control plants (Fig. 3a). Control internode diameters were intrinsically larger as exemplified by internode one (Fig. 3b). The maximum internode volume and mass were reached by internode four in both the CEPA-treated and control primary culms (Fig. 3c, d). This indicated that the older internodes had completed their growth. The internodes in each group reached their maximum volume differently. In the treated plants, internode four was significantly longer than internode three (65.5 ± 6.2 mm for internode three compared to 79 ± 2.7 mm for internode four, P < 0.05, n = 6; Fig. 3a). In the control plants, the diameter of internode four was significantly greater than internode three (17.1 ± 0.6 mm for internode three compared to 19.1 ± 0.3 mm for internode four, P < 0.01, n = 6; Fig. 3b). Hence, the treated internode three completed growth by elongation in contrast to the control internode three which completed growth by diameter expansion. As the CEPA-treated culms possessed almost twice as many internodes (Table 2), they had more mature internodes on their primary culm than the control plants.
The plant extension rate for each primary shoot was obtained by measuring the distance between the soil line to the dewlap of the last fully expanded leaf on a fortnightly basis over a period of 6 weeks prior to harvesting the first trial. The average plant extension rate of the CEPA-treated group was significantly greater than that of the control group (13 ± 0.6 mm day−1 for the control compared to 15.6 ± 0.6 mm day−1 for CEPA, P < 0.01, n = 12 per treatment). This result was corroborated by the average culm elongation rate, which was calculated from the final culm length and the growth time from the 3½ mpg data (11.6 ± 0.2 mm day−1 for the control compared to 14.8 ± 0.4 mm day−1 for CEPA, P < 0.001, n = 6 per treatment).
Both the internode elongation rate and the internode formation rate (which is related to the phyllochron) contribute to the culm elongation rate. Although the internode elongation rate was not directly measured in this experiment, it can be estimated from the increase in length between expanding internodes such as internodes one and three. The average internode elongation rate calculated from internodes one and three was not significantly different between the CEPA-treated and control groups (6.8 ± 0.7 mm day−1 for CEPA and 6.5 ± 0.7 mm day−1 for the control, P > 0.05, n = 6 per treatment). Therefore, faster culm elongation was probably solely due to faster internode generation. This was represented by the presence of more actively elongating internodes on the culm. The elongating internodes in the CEPA-treated culms were internodes one, two, and three but only internodes one and two in the control culms.
CEPA Treatment Increased Tiller Formation but Reduced Leaf Fresh Weight and Leaf Area
Tiller formation was prolific in the treated pots, with tillers forming approximately twice as fast as the controls (0.64 ± 0.06 tillers per week versus 0.27 ± 0.02 tillers per week, P < 0.01, n = 12 per treatment, Fig. 2b). Accordingly, the final tiller count was twofold higher in the treated pots (Table 2). The total tiller fresh weight per pot was comparable between the two groups because the tillers in the CEPA-treated group were numerous but small (with respect to culm and leaf size) and less abundant but larger in the control group. Some buds on the lower internodes of the primary culm germinated early with adventitious roots also forming on those internodes (Fig. 4b). These structures did not develop on the control plants prior to 3½ mpg.
In control and CEPA-treated plants at 2 mpg, the total primary leaf fresh weight represented the largest proportion of the aerial biomass followed by the primary culm and tillers (Table 2). This trend was also evident in the control plants at 3½ mpg. However, in CEPA-treated plants at 3½ mpg, the primary culm fresh weight was the largest proportion of the aerial biomass, followed by total primary leaves and tillers (Table 2).
Although the CEPA-treated plants produced more green leaves than the control plants, on a fresh weight basis they represented a smaller fraction of the aerial biomass at 2 and 3½ mpg when compared to the controls (Table 2). Also, the total mass of each leaf component [top (the unexpanded leaves), lamina, and sheath] on the treated plants was smaller than that of the control plants (Table 2). The treated leaf sheaths were shorter and their associated laminas had shorter lengths and narrower widths (results not shown). Consequently, there was approximately 56% less total green leaf area per plant at 3½ mpg (1433 ± 97 cm2 for CEPA and 3273 ± 164 cm2 for the control, P < 0.0001, n = 6 per treatment). The percentage of leaves that were still green was similar in the two groups but was lower in both groups at 3½ mpg.
The SLA decreased between the second-last fully expanded leaf and the sixth-last fully expanded leaf in the CEPA-treated group but did not change significantly in the control group (Fig. 5). To determine whether the difference in SLA between treated and control leaves was due to the accumulation of soluble carbohydrates in the leaves, the SLA was also examined on a structural dry weight basis, that is, total dry weight minus soluble carbohydrates. It was found that the SLA profile expressed on a structural dry weight basis was similar to the profile expressed on a total dry weight basis (results not shown).

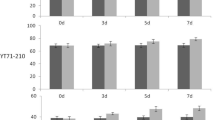
The specific leaf area of the second-, fourth-, and sixth-last fully expanded leaves on the primary culm of CEPA-treated sugarcane at 3½ mpg. Control (filled bar), CEPA (open bar). The data plotted are the mean ± standard error of measurements collected from six independent plants per treatment. Significant differences were identified by analysis of variance followed by post hoc comparisons using Tukey’s HSD (honesty significance difference) test. Means with the same letter were not significantly different at P ≤ 0.05
CEPA Treatment Increased Leaf Photosynthetic Rate
The photosynthetic rate of the fourth-last fully expanded leaf was 38% higher at 400 μmol CO2 mol−1 (approximately ambient CO2 level) in the CEPA-treated plants compared to that of the control plants (Table 3). The CO2 assimilation kinetics of the fourth-last fully expanded leaf was characterized through CO2 response curves (Fig. 6). At low CO2 levels, where gas supply limits photosynthesis, the CEPA-treated leaves assimilated CO2 more efficiently as confirmed by the higher carboxylation efficiency (CE) (Table 3). Furthermore, the CO2-saturated photosynthetic rate (J max) was higher in the CEPA-treated leaves (Table 3).

The photosynthetic rate (J) plotted as a function of intercellular CO2 concentration (C i) for CEPA-treated leaves. The measurements were taken from the fourth-last fully expanded leaf on the primary culm at 3 mpg. Control (filled circle), CEPA (open circle). The data plotted are the mean ± standard error of measurements collected from four independent plants per treatment. The line of best fit for control is J = 0.345 + 33.968(1 − e−0.01Ci), r 2 = 0.99, P < 0.0001 and for CEPA, J = -0.819 + 45.182(1 − e−0.013Ci), r 2 = 0.99, P < 0.0001
Chlorophyll levels were measured in the fifth-last fully expanded leaf to gauge the light-harvesting potential of the leaves. The concentration of chlorophylls a and b were significantly higher in the treated plants (Table 3), but ratios of chlorophyll a/b were not significantly different between the two groups (ratios not shown).
Because stomata govern the diffusive conductance of water vapour and CO2, stomata-related measurements were collected from the fourth-last fully expanded leaf at the same time as the photosynthesis measurement. The stomatal conductance and the transpiration rate were higher in the CEPA-treated group (Table 3). Because the photosynthetic rate is normalized on an area basis, stomatal density could affect the photosynthetic rate. Enumeration of stomata on the abaxial and adaxial surfaces of treated leaves revealed fewer stomata per unit area on the abaxial surface but no significant difference on the adaxial surface (data not shown). This resulted in fewer total stomata per unit of leaf area (Table 3). It was also noted that the average length of the stomatal aperture was larger in the treated leaves (Table 3).
CEPA Treatment Increased Vacuolar Acid Invertase and Cell Wall Invertase Expression in Internode Four
A simple gene expression model comprising six genes was used to evaluate the relationship between sucrose storage, growth, and respiration in internode four. Real-time quantitative PCR analysis indicated that there was no significant difference in expression (relative to ACT) of SPS, CES, EXP, and PFP between the two groups (Table 4). However, VAI and CWI expression was 3.3 and 1.4 times higher, respectively, in the CEPA-treated sugarcane (Table 4).
Discussion
Rapid Internode Development Led to Faster Sucrose Accumulation
Our results indicated that the CEPA treatment induced early internode development and increased the sucrose accumulation rate in the sugarcane culm. The rapid internode development in the CEPA-treated sugarcane was reflected by the increase in number of internodes with smaller volumes (Fig. 3c). Smaller internodes may fill with sucrose quicker and can lead to earlier ripening of the internode. However, this does not entirely explain the rapid sucrose accumulation observed as there were more internodes to fill but there was no difference in the primary culm volume.
The internodes from the CEPA-treated and control sugarcane may be compared on the basis of their chronological age or physiological age. However, their chronological age will not reflect their physiological age because of the difference in internode formation rates between the two groups. For example, internode four was 14 days old in the CEPA-treated group whereas in the control group it was 20 days old. However, based on the internode volume profile (Fig. 3c), internode four was the last fully expanded internode in both the treated and control groups and hence can be considered physiologically equivalent. Therefore, although the internodes were chronologically different, the CEPA-treated internode four finished expanding earlier. The implication is that the CEPA-treated sugarcane produced fully expanded internodes faster, which was presumably linked to the shorter phyllochron. Consequently, there were 15 fully expanded internodes (that is, internode four and all the older internodes) representing 95% of the total culm volume in the CEPA-treated plants at 3½ mpg. In contrast, the control group had only seven fully expanded internodes representing 85% of the total culm volume. Hence, the early formation as well as faster development of new internodes in CEPA-treated sugarcane led to faster formation of fully expanded culm tissue. This enabled internodes to ripen sooner and is succinctly demonstrated by comparing the sucrose concentration in internode four (Fig. 1a) where the 14-day-old CEPA-treated internode accumulated a similar concentration of sucrose as the 20-day-old control internode (204 ± 19 mg g−1 DW for CEPA and 178 ± 21 mg g−1 DW for the control, P > 0.05, n = 6 per treatment). Faster generation of fully expanded culm tissue increases sink strength thereby promoting faster sucrose storage.
Higher Photosynthesis and Reduced Leaf Expansion May Favour Faster Sucrose Accumulation
Stomatal and biochemical constraints dictate leaf CO2 assimilation characteristics. In C4 plants, the leaf photosynthetic rate reaches a maximum value (J max) at high intercellular CO2 concentrations due to various biochemical limitations, including the CO2-fixing capacity of the C4 cycle in the mesophyll cells or the capacity of the photosynthetic carbon reduction (PsCR) cycle in the bundle sheath cells. The CE and J max were larger in the treated leaves than in the control leaves (Table 3), which implied that the stomatal and/or biochemical limitation was less in the treated leaves. The higher leaf chlorophyll levels observed in the treated plants (Table 3) could improve the ATP and NADPH supply which in turn may improve the energy and reducing power supply to the C4 and/or PsCR reactions leading to enhanced photosynthesis capability. Stomatal regulation of the CO2 supply also contributed to the observed difference in photosynthesis because the stomatal conductance and transpiration rate were higher in CEPA-treated leaves. As ethylene can inhibit abscisic acid-induced stomatal closure (Tanaka and others 2005), the increased transpiration rate in treated plants could have been due to the pores opening further. To confirm if this was the case, the stomatal apertures were compared by microscopic examination of silicon lamina surface impressions. This revealed that the width of the stomatal pore was unaffected by CEPA treatment (Table 3).
In some Gramineae, exposure to CEPA causes the formation of smaller leaves (by reducing the number of epidermal cells) and also increases the rate of leaf emergence (Van Andel 1973; Van Andel and Verkerke 1978). The leaves of CEPA-treated sugarcane were also smaller and emerged faster. The younger CEPA-treated leaves exhibited a higher SLA, which declined for progressively older leaves (Fig. 5). This relationship between SLA and leaf position is interesting because the last fully expanded leaf on a sugarcane plant does not undergo any further area expansion (Van Dillewijn 1952). The majority of the leaf structural matter is deposited early in the leaf’s development between leaf tip appearance and full expansion. Therefore, the SLA profile observed in the treated plants may simply be an intrinsic property of the internode/leaf position.
The physiological basis of dry matter partitioning can be explored through the SLA, which is a practical measure of the material cost of producing leaf area. The significantly higher SLA for leaf two in the treated plants indicates that a smaller amount of dry weight was used to produce each unit of leaf area. Hence, less photoassimilate was incorporated into leaf dry matter and more was potentially partitioned to sink tissue. As sugarcane leaves respire faster than the culm on a dry weight basis (Glover 1973) and the respiration rate is generally dictated by leaf growth rate, it is possible that less foliage in the CEPA-treated sugarcane reduced respiratory carbon losses from the system. In addition, there was less leaf area and it was constructed more efficiently using less photoassimilate. This implies less carbon was invested in foliage infrastructure. Coupled with the higher photosynthesis to offset the lower leaf area available for CO2 capture, this resulted in more carbon partitioned to the culm and sucrose. Some benefit may have also accrued from the change in light interception pattern. In CEPA-treated plants, the lower leaves would capture more sunlight because the upper leaves were smaller.
Higher sink strength has been simulated in sugarcane by reducing the leaf area through shading (McCormick and others 2006) or defoliation (McCormick and others 2008b). This had the effect of increasing leaf photosynthesis and lowering sucrose levels in the culm. Those studies concluded that decreased assimilate availability at the sink tissue may invoke the upregulation of source tissue photosynthesis. In addition, they suggested that sink demand and source photosynthetic activity was linked through a kinase-mediated sugar signalling mechanism that controlled the expression of metabolite transporters and photosynthesis-related genes (McCormick and others 2008a). The reduced leaf area and increased photosynthetic rate observed in our work are somewhat analogous to those of the previous work. However, this did not correspond to lower sucrose levels in the culm. In our study, the culm sucrose level probably did not instigate the increase in source activity. However, the higher demand for carbon due to the accelerated rate of culm maturation may have been perceived as “low sucrose” and subsequently caused the increase in photosynthetic rate. Clearly, further investigations are needed to fully unravel this aspect of the source–sink relationship in sugarcane.
CEPA Treatment did not Affect Culm Structural Biomass Accumulation
The difference in moisture content between the treated and control plants did not account for the difference in the leaf to culm fresh weight ratio observed between the two groups. Thus, the lower leaf to culm fresh weight ratio of the CEPA-treated primary shoots indicated that they tended to partition more carbon to the culm. To gain further insight into this aspect, the allocation of carbon between sucrose storage and culm structural biomass synthesis was investigated by examining the expression of specific genes in internode four.
CEPA treatment did not alter the structural biomass composition, as the average amount of insoluble solids and the amount of cellulose in the insoluble solids were unaffected (Table 1). This is consistent with the fact that CES expression in internode four was unchanged. Nonetheless, the elevated reducing sugars present in the internode may increase the metabolic flux through the pathway involving CES, thus enabling faster internode growth. Similarly, despite no changes in PFP expression, the metabolic flux through the respiratory pathway involving PFP may increase due to the higher reducing sugars. Indeed, faster respiration would be expected to accompany faster internode growth.
In sugarcane, there is a positive correlation between total acid invertase enzyme activity and internode elongation (Glasziou and Bull 1965). Acid invertase hydrolyses sucrose into reducing sugar to fuel respiration and provide structural carbohydrates for tissue elongation. The elevated osmotic pressure within the cell arising from the high hexose levels can also contribute to cell elongation (Morris and Arthur 1984). The dramatic difference in primary culm length at 2 mpg indicated that internode elongation was vigorous in the CEPA-treated culms. Furthermore, a significant amount of tissue elongation occurred during the transformation of internode three into internode four in the CEPA-treated culms compared to the corresponding internodes in the control culms. This growth behaviour is consistent with the higher VAI and CWI expression (Table 4) and the elevated reducing sugar concentration observed in the CEPA-treated internode four (Fig. 1b).
Potential Causes of the Sugarcane CEPA Response
The genes responsible for the phenotype observed in CEPA-treated sugarcane are presently unknown. Ethylene interacts with other phytohormones to control plant growth and development. For example, ethylene can regulate auxin biosynthesis in a tissue-specific manner (Swarup and others 2007; Stepanova and others 2008). Thus, the ethylene effect may be manifested through complex interactions with other phytohormones. Auxin is believed to control apical dominance in plants (Cline 1997). It is synthesized at the shoot tip and transported basipetally through the culm where it inhibits lateral bud germination. In some plants ethylene inhibits polar auxin transport in the shoot (Morgan and Gausman 1966; Burg and Burg 1967) leading to depletion of auxin in the lower regions of the shoot and a localized auxin surplus at the shoot apex. Moreover, the influence of ethylene was also proven by the reduction of auxin movement in the culm of L. esculentum that was engineered to exhibit a constitutive ethylene response (Whitelaw and others 2002). Auxin is an important factor regulating the development of leaf primordia (Shani and others 2006). However, elevated levels inhibit leaf expansion (Keller and others 2004). Interestingly, auxin also increases acid invertase enzyme activity in sugarcane and other plants (Sacher and others 1963; Morris and Arthur 1984). Perhaps similar ethylene-auxin interactions in sugarcane were responsible for suppressing apical dominance (lateral shoots and tillers), reduced leaf size, and increased internode VAI and CWI expression.
In summary, we have shown that protracted exposure to low ethylene levels beginning from shoot emergence can induce higher sucrose accumulation in sugarcane from an early stage of culm development. Based on these findings, we postulate that early internode ripening may have been facilitated by (1) favourable partitioning of dry matter between source and sink tissue, (2) earlier initiation of internodes, and (3) faster internode formation. Ethylene inhibited leaf growth but not photosynthesis; thus, more photoassimilate was transferred to sink tissue during leaf growth. Internodes were initiated early and formed quickly enabling them to reach maturity sooner. This promoted a stronger storage sink that was more conducive to early sucrose storage. It is likely that the higher sucrose turnover in the vacuole and increased apoplastic uptake of reducing sugars influenced the rate of culm development and sucrose storage. These findings lay the foundation for a hypothesis that suggests that there is direct causality between these growth traits and early internode ripening in sugarcane. Although these traits were induced artificially, they may explain why some sugarcane varieties ripen earlier than others. Furthermore, the influence of endogenous ethylene production and ethylene sensitivity on natural sugarcane ripening would be interesting future work.
References
Alexander AG (1973) Sugarcane physiology. A comprehensive study of the Saccharum source-to-sink system. Elsevier Scientific, New York, 752 pp
Breaux RD (1984) Breeding to enhance sucrose content of sugarcane varieties in Louisiana. Field Crop Res 9:59–67
Bull TA, Glasziou KT (1963) The evolutionary significance of sugar accumulation in Saccharum. Aust J Biol Sci 16:737–742
Burg SP, Burg EA (1967) Inhibition of polar auxin transport by ethylene. Plant Physiol 42:1224–1228
Castleden CK, Aoki N, Gillespie VJ, MacRae EA, Quick WP, Buchner P, Foyer CH, Furbank RT, Lunn JE (2004) Evolution and function of the sucrose-phosphate synthase gene families in wheat and other grasses. Plant Physiol 135:1753–1764
Casu RE, Jarmey JM, Bonnett GD, Manners JM (2007) Identification of transcripts associated with cell wall metabolism and development in the stem of sugarcane by Affymetrix GeneChip Sugarcane Genome Array expression profiling. Funct Integr Genomic 7:153–167
Chomczynski P, Sacchi N (1987) Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem 162:156–159
Cline MG (1997) Concepts and terminology of apical dominance. Am J Bot 84:1064–1069
Cox MC, Hogarth DM, Hansen PB (1994) Breeding and selection for high early season sugar content in a sugarcane (Saccharum spp. hybrids) improvement program. Aust J Agr Res 45:1569–1575
Domir SC, Foy CL (1978) A study of ethylene and CO2 evolution from ethephon in tobacco. Pestic Biochem Phys 9:1–8
Donaldson RA, Van Staden J (1989) A review of chemicals used as ripeners of sugarcane in South Africa. Proc Int Soc Sugar Cane Technol 20:647–655
Eastwood D (1979) Tillering and early growth of sugar-cane setts in response to pre-plant treatment with (2-chloroethyl) phosphonic acid. Trop Agr (Trinidad) 56:11–17
Fiorani F, Bogemann GM, Visser EJW, Lambers H, Voesenek LACJ (2002) Ethylene emission and responsiveness to applied ethylene vary among Poa species that inherently differ in leaf elongation rates. Plant Physiol 129:1382–1390
Fong Chong B, Bonnett GD, Glassop D, O’ Shea MG, Brumbley SM (2007) Growth and metabolism in sugarcane are altered by the creation of a new hexose-phosphate sink. Plant Biotechnol J 5:240–253
Glasziou KT, Bull TA (1965) The relation between total invertase activity and internode expansion in sugarcane stalks. Proc Int Soc Sugar Cane Technol 12:575–581
Glasziou KT, Bull TA, Hatch MD, Whiteman PC (1965) Physiology of sugar-cane. VI. Effects of temperature, photoperiod duration, and diurnal and seasonal temperature changes on growth and ripening. Aust J Biol Sci 18:53–66
Glover J (1973) The dark respiration of sugar-cane and the loss of photosynthate during the growth of a crop. Ann Bot 37:845–852
Gosnell JM, Lonsdale JE (1974) Some effects of drying off before harvest on cane yield and quality. Proc Int Soc Sugar Cane Technol 15:701–712
Groenewald J-H, Botha FC (2008) Down-regulation of pyrophosphate: fructose 6-phosphate 1-phosphotransferase (PFP) activity in sugarcane enhances sucrose accumulation in immature internodes. Transgenic Res 17:85–92
Grof CPL, So CTE, Perroux JM, Bonnett GD, Forrester RI (2006) The five families of sucrose-phosphate synthase genes in Saccharum spp. are differentially expressed in leaves and stem. Funct Plant Biol 33:605–610
Inman-Bamber NG, Bonnett GD, Spillman MF, Hewitt ML, Jackson J (2008) Increasing sucrose accumulation in sugarcane by manipulating leaf extension and photosynthesis with irrigation. Aust J Agr Res 59:13–26
Iskandar HM, Simpson RS, Casu RE, Bonnett GD, Maclean DJ, Manners JM (2004) Comparison of reference genes for quantitative real-time polymerase chain reaction analysis of gene expression in sugarcane. Plant Mol Biol Rep 22:325–337
Keller CP, Stahlberg R, Barkawi LS, Cohen JD (2004) Long-term inhibition by auxin of leaf blade expansion in bean and Arabidopsis. Plant Physiol 134:1217–1226
Lingle SE (1997) Seasonal internode development and sugar metabolism in sugarcane. Crop Sci 37:1222–1227
Mamet LD, Galwey NW (1999) A relationship between stalk elongation and earliness of ripening in sugarcane. Exp Agr 35:283–291
McCormick AJ, Cramer MD, Watt DA (2006) Sink strength regulates photosynthesis in sugarcane. New Phytol 171:759–770
McCormick AJ, Cramer MD, Watt DA (2008a) Changes in photosynthetic rates and gene expression of leaves during a source-sink perturbation in sugarcane. Ann Bot 101:89–102
McCormick AJ, Cramer MD, Watt DA (2008b) Culm sucrose accumulation promotes physiological decline of mature leaves in ripening sugarcane. Field Crop Res 108:250–258
McDonald L (2006) The effect of time of ratooning on sugarcane growth in the Burdekin. Proc Aust Soc Sugar Cane Technol 28:261–272
Moore PH, Osgood RV (1989) Prevention of flowering and increasing sugar yield of sugarcane by application of ethephon (2-chloroethylphosphonic acid). J Plant Growth Regul 8:205–210
Moore PH, Botha FC, Furbank RT, Grof CPL (1997) Potential for overcoming physio-chemical limits to sucrose accumulation. In: Keating BA, Wilson JR (eds), Intensive sugarcane production: meeting the challenges beyond 2000. Proceedings of the sugar 2000 symposium. CAB International, Wallingford, pp 141-156
Morgan PW, Gausman HW (1966) Effects of ethylene on auxin transport. Plant Physiol 41:45–52
Morgan T, Jackson P, McDonald L, Holtum J (2007) Chemical ripeners increase early season sugar content in a range of sugarcane varieties. Aust J Agr Res 58:233–241
Morris DA, Arthur ED (1984) Invertase and auxin-induced elongation in internodal segments of Phaseolus vulgaris. Phytochemistry 23:2163–2167
Nayamuth AR, Cheeroo-Nayamuth BF, Soopramanien GC (1999) Agro-physiological characteristics underlying the sucrose accumulation pattern of early and late varieties. Proc S Afr Sug Technol Assoc 73:157–163
Pammenter NW, Allison JCS (2002) Effects of treatments potentially influencing the supply of assimilate on its partitioning in sugarcane. J Exp Bot 53:123–129
Pfaffl MW (2001) A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res 29:2002–2007
Pierik R, Tholen D, Poorter H, Visser EJW, Voesenek LACJ (2006) The Janus face of ethylene: growth inhibition and stimulation. Trends Plant Sci 11:176–183
Porra RJ, Thompson WA, Kriedermann PE (1989) Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: verification of the concentration of chlorophyll standards by atomic absorption spectrophotometry. Biochim Biophys Acta 975:384–394
Quackenbush J, Liang F, Holt I, Pertea G, Upton J (2000) The TIGR Gene Indices: reconstruction and representation of expressed gene sequences. Nucleic Acids Res 28:141–145
Roitsch T, Balibrea ME, Hofmann M, Proels R, Sinha AK (2003) Extracellular invertase: key metabolic enzyme in PR protein. J Exp Bot 54:513–524
Rostron H (1973) The effect of chemical ripeners on the growth, yield and quality of sugarcane in South Africa and Swaziland. Proc S Afr Sug Technol Assoc 47:191–200
Rostron H (1977) Prolonged chemical ripening of sugarcane following multiple applications of ethrel. Proc Int Soc Sugar Cane Technol 16:1743–1753
Sacher JA, Hatch MD, Glasziou KT (1963) Regulation of invertase synthesis in sugar cane by an auxin- and sugar-mediated control system. Physiol Plantarum 16:836–842
Shani E, Yanai O, Ori N (2006) The role of hormones in shoot apical meristem function. Curr Opin Plant Biol 9:484–489
Solomon S, Singh I, Madam VK (1998) Effect of 2-chloroethyl phosphonic acid on early growth and advancement of maturity in sugarcane. Proc Sug Technol Assoc India 60:A10–A16
Stepanova AN, Robertson-Hoyt J, Yun J, Benavente LM, Xie D.-Y, Dolezal K, Schlereth A, Jurgens G, Alonso JM (2008) TAA1-mediated auxin biosynthesis is essential for hormone crosstalk and plant development. Cell 133:177–191
Stewart DW, Dwyer LM (1999) Mathematical characterization of leaf shape and area of maize hybrids. Crop Sci 39:422–427
Swarup R, Perry P, Hagenbeek D, Van Der Straeten D, Beemster GTS, Sandberg G, Bhalerao R, Ljung K, Bennett MJ (2007) Ethylene upregulates auxin biosynthesis in Arabidopsis seedlings to enhance inhibition of root cell elongation. Plant Cell 19:2186–2196
Tanaka Y, Sano T, Tamaoki M, Nakajima N, Kondo N, Hasezawa S (2005) Ethylene inhibits abscisic acid-induced stomatal closure in Arabidopsis. Plant Physiol 138:2337–2343
Untergasser A, Hijveen H, Rao X, Bisseling T, Geurts R, Leunissen JAM (2007) Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res 35:W71–W74
Updegraff DM (1969) Semimicro determination of cellulose in biological materials. Anal Biochem 32:420–424
Van Andel OM (1973) Morphogenetic effects on vegetative plants of Poa pratensis L. of 6-azauracil, (2-chloroethyl)-phosphonic acid, and (2-chloroethyl)trimethyl-ammonium chloride and their interaction with gibberellic acid. J Exp Bot 24:245–257
Van Andel OM, Verkerke DR (1978) Stimulation and inhibition by ethephon of stem and leaf growth of some Gramineae at different stages of development. J Exp Bot 29:639–651
Van Dillewijn C (1952) Botany of sugarcane. Chronica Botanica, Waltham, 371 pp
Warner HL, Leopold AC (1971) Timing of growth regulator responses in peas. Biochem Biophys Res 44:989–994
Weyers JDB, Johansen LG (1985) Accurate estimation of stomatal aperture from silicone rubber impressions. New Phytol 101:109–115
Whitelaw CA, Lyssenko NN, Chen L, Zhou D, Mattoo AK, Tucker ML (2002) Delayed abscission and shorter internodes correlate with a reduction in the ethylene receptor LeETR1 transcript in transgenic tomato. Plant Physiol 128:978–987
Whittaker A, Botha FC (1999) Pyrophosphate: d-fructose-6-phosphate 1-phosphotransferase activity patterns in relation to sucrose storage across sugarcane varieties. Physiol Plantarum 107:379–386
Zhu YJ, Komor E, Moore PH (1997) Sucrose accumulation in the sugarcane stem is regulated by the difference between the activities of soluble acid invertase and sucrose phosphate synthase. Plant Physiol 115:609–616
Acknowledgments
This work was supported by the Cooperative Research Centre for Sugar Industry Innovation through Biotechnology and the Queensland Department of State Development, Trade and Innovation (through a Smart State Fellowship to AG).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Fong Chong, B., Mills, E., Bonnett, G.D. et al. Early Exposure to Ethylene Modifies Shoot Development and Increases Sucrose Accumulation Rate in Sugarcane. J Plant Growth Regul 29, 149–163 (2010). https://doi.org/10.1007/s00344-009-9118-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00344-009-9118-3