
Abstract
Exogenous brassinosteroid (BR) has been reported to improve plant resistance to abiotic stress, but little is known about the role of endogenous BR in plant stress responses. In this study we investigated the involvement of endogenous BR in salt stress response using BR mutants det2-1 and bin2-1 of Arabidopsis. Seed germination and seedling growth of det2-1 and bin2-1 were more sensitive to salt stress than that of Columbia wild type (WT). The transcript levels of salt- and ABA-induced genes COR78 and P5CS1 were less induced in det2-1 than in WT under 200 mM NaCl. In addition, the basal proline level and, to a lesser extent, the proline level induced by 200 mM NaCl or 50 μM ABA in both det2-1 and bin2-1 was enhanced, resulting in decreased proline accumulation. On the other hand, exogenous 24-epibrassinolide (EBR) could enhance proline accumulation, promote root elongation of WT, and partially rescue the growth of det2-1 under salt stress. These results suggested that endogenous BR is positively involved in the plant response to salt stress in Arabidopsis.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
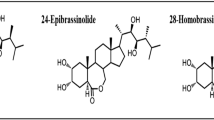
Phytohormones such as abscisic acid (ABA), ethylene, and jasmonic acid have been reported to be involved in plant responses to abiotic stress. Brassinosteroids (BRs) are a class of plant polyhydroxysteroids that are structurally similar to animal and insect steroid hormones and control a broad range of plant developmental processes (Clouse and Sasse 1998). Several BR signaling components have been identified and characterized using BR-deficient or BR-insensitive mutants. The membrane-localized receptors BRI1 and BAK1 transfer the BR signal into the cytoplasm to inhibit BIN2, a glycogen synthase kinase-3/SHAGGY-like kinase. BIN2 negatively regulates downstream signaling by phosphorylating two transcription factors BES1 and BZR1 and inhibiting their binding to BR target promoters (Wang and He 2004; Li and Jin 2006). Arabidopsis DET2 encodes a BR biosynthetic enzyme that shares significant sequence identity with the mammalian steroid 5α-reductase. The defects in root and leaf development of the mutant det2-1 could be rescued by the application of brassinolide (Li and others 1996).
Application of exogenous BR has been reported to improve plant resistance to many environmental stresses such as salinity stress (Anuradha and Rao 2001, 2003), drought stress (Kagale and others 2007), thermal stress (Dhaubhadel and others 2002; Singh and Shono 2005; Ogweno and others 2008), and heavy-metal stress (Hayat and others 2007). However, the function of endogenous BR under stress is not clear. Recently, a rice knockout mutant of a salt-responsive OsGSK1, an ortholog of Arabidopsis BIN2, was reported to be hypersensitive to BR and more tolerant to abiotic stress (Koh and others 2007).
From this study we provide preliminary evidence that endogenous BR is involved in salt stress response by demonstrating that the Arabidopsis BR-deficient mutant det2-1 and BR-insensitive mutant bin2-1 are hypersensitive to salinity stress during seed germination and/or the seedling growth stage. Moreover, the hypersensitivity correlated with the inhibited induction of stress-related genes COR78 and P5CS1 and proline accumulation under 200-mM-NaCl stress. Addition of 24-epibrassinolide (EBR) could improve the NaCl-induced proline accumulation and alleviate the inhibition of root elongation in wild type (WT). Our data suggested that BR plays an important role in the Arabidopsis response to environmental stresses.
Materials and Methods
Plant Materials and Growth Conditions
Arabidopsis thaliana ecotype Columbia was used as WT control. BR-deficient mutant det2-1 and BR-insensitive mutant bin2-1 were obtained from Dr. Zhiyong Wang and Dr. Jun Zhao. WT and det2-1 seeds were surface-sterilized in 10% sodium hypochlorite and rinsed five times in sterile water. The seeds were then sown on half-strength MS (1/2 MS) agar medium with 1% sucrose for normal germination, or on the same basic medium supplemented with different concentrations of NaCl and ABA for stress treatment. After being stored at 4°C for 72 h, seeds were germinated in a growth chamber at 22°C under 16 h of light (120 μEm−2 s−1), E represents Einstein.
Growth Measurement
The salt sensitivity of mutant and wild-type seedlings was analyzed by transferring seedlings grown on 1/2 MS medium (1% sucrose) for 4 days (det2-1) or 7 days (bin2-1) onto NaCl-containing medium with or without different concentrations of EBR. The seedlings were allowed to grow horizontally for the desired period of time. The seedlings were then carefully pulled out from the medium for photographing, determination of fresh weight (dead seedlings were counted as zero), and root length measurement. The inhibition of fresh weight or root elongation of WT and mutants was calculated as follows: [(control seedlings − treated seedlings) × 100] ÷ control seedlings.
Salt and ABA Treatment
Sterilized seeds of mutant and WT were germinated on nylon mesh placed on 1/2 MS medium. The mesh with 10-day-old seedlings was lifted and incubated in liquid 1/2 MS medium containing 200 mM NaCl or 50 μM ABA, with or without preincubation for 3 days in medium with EBR. The seeds were harvested at the desired time points and frozen in liquid nitrogen for proline content measurement and RNA extraction.
Proline Content Determination
Proline content of the seedlings was measured according to the method of Bates and others (1973). Twenty to twenty-five seedlings were pooled in each replica for proline determination. Folds of proline accumulation were calculated as follows: proline content of treated sample ÷ proline content of control.
Isolation of RNA and Semiquantitative RT-PCR
Total RNA was extracted from 10-day-old seedlings as described by Hua and others (2001). Twenty to twenty-five seedlings were pooled for total RNA extraction. First-strand cDNA was synthesized by Superscript II following the manufacturer’s instructions (Invitrogen, Carlsbad, CA, USA). PCR, using the first-strand cDNA as a template, was performed using the following primers: COR78F: 5′-TGGAACAGAGGATGTAACGAG-3′ and COR78R: 5′-CGGAGTCAACTTCTCATCAAC-3′ to amplify a 1084-bp COR78 (At5g52310) fragment. RD22F: 5′-ATGGCGATTCGGCTTCCTCTG-3′ and RD22R: 5′-CTAGTAGCTGAACCACACAAC-3′ were used for full-length RD22 (At5g25610) cDNA. The primers for P5CS1 (At2g39800) were P5CS1F: 5′-AGTCT-ATGCTTGATTTGAGGGT-3′ and P5CS1R: 5′-AAGCTCATTAAGCACAGCATTC-3′. A 380-bp fragment of ACTIN7 (At5g09180), an internal control, was amplified by ACTIN7F: 5′-TGCACAAGTCATAACCATCGG-3′ and ACTIN7R: 5′-TGTGAACAATCGATGGACCTGAC-3′. PCR amplification of all four genes was performed by an initial denaturation at 94°C for 4 min followed by 25 cycles of denaturing at 94°C for 45 s, annealing at 55°C for 45 s, extension at 72°C for 60 s, and a final extension at 72°C for 7 min. Quantification of the amplified signals was performed by photographing the gel and analyzing it using the software Band Leader (Version 3.0, Magnitec Ltd.)
Results
Hypersensitivity of det2-1 to Salt Stress During Germination
To understand whether endogenous BR is involved in the salt response during seed germination, the seeds of det2-1 and WT were germinated on salt-containing medium. The germination rates were counted every day for 7 days. The germination rates of WT were 100% under all NaCl concentrations tested within 7 days. On the other hand, the germination rate of det2-1 was close to 100% only under control conditions and was only 84 and 75% under 50 and 100 mM NaCl, respectively (Fig. 1a). Under 200 mM NaCl, less than 45% of det2-1 seeds germinated whereas WT seeds reached 100% (data not shown). Not only the germination rate but also the speed of germination was reduced in det2-1 under salt stress. Under 0 mM NaCl, WT seeds germinated within the first day, whereas only 24% of det2-1 seeds germinated. Under salt stress, the germination of det2-1 was further delayed compared with that of WT (Fig. 1a).

Effects of salt stress (a) or ABA treatment (b) on the germination rate of det2-1. Data are the mean ± standard deviation of three independent measurements of 300 seeds
It is well established that phytohormone ABA could inhibit seed germination and is involved in a plant’s response to salt stress. Because germination of det2-1 was hypersensitive to salt stress, its sensitivity to ABA treatment was also checked. The germination rate of det2-1 was reduced more than that of WT after 0.1-μM-ABA treatment (Fig. 1b), which is in agreement with a previous report (Steber and McCourt 2001) that the germination of det2-1 was hypersensitive to ABA. These results indicated that endogenous BR has a positive effect on seed germination under salt stress or ABA treatment.
Altered Salt Stress Sensitivity of det2-1 During Seedling Growth
To characterize the salt sensitivity of det2-1 at the seedling growth stage, 7-day-old seedlings were grown on 1/2 MS medium with 50, 100, and 150 mM NaCl for 3 days. On the control medium, det2-1 seedlings showed darker green leaves and shorter roots than WT. Under 100 mM NaCl, leaves of det2-1 seedlings turned pale rapidly, whereas the leaves of WT were still partially green (Fig. 2a, panels a and b).

Effects of salt stress on det2-1 seedling growth. a Effects of EBR on 100-mM-NaCl-stressed det2-1 and WT seedlings. Seedlings grown on normal medium for 7 days were transferred onto NaCl-containing medium and grown for another 3 days; (a) CK, (b) 100-mM-NaCl treatment, (c) cotreatment with 100 mM NaCl and 1 nM EBR, (d) cotreatment with 100 mM NaCl and 10 nM EBR. b Relative inhibition of fresh weight and root length of det2-1 (mean ± SD, n = 4) under salt stress. CK: seedlings grown on normal 1/2 MS without NaCl
We further examined the root length and fresh weight of det2-1. As shown in Fig. 2b, the root elongation of det2-1 seedlings was inhibited more severely than that of WT at all concentrations of NaCl. The inhibition of det2-1 root elongation was 86.7% under 100 mM NaCl, twice as much as that of WT. The fresh weight of det2-1 was inhibited more than that of WT under 50 and 100 mM NaCl (Fig. 2b). The inhibition of fresh weight of det2-1 was 100% because all the det2-1 seedlings were dead under 150 mM NaCl. These results indicate that det2-1 is hypersensitive to salt stress during the seedling stage.
To investigate whether the salt sensitivity of det2-1 is due to its lack of wild-type level of BR, different concentrations of EBR were added back to the salt-containing medium. As shown in Fig. 2a (panels c and d), the bleaching of the leaves of det2-1 was slowed greatly by 10 nM of EBR, suggesting that addition of BR can partially rescue the growth of det2-1 under salt stress.
Altered Salt Stress-Induced Gene Expression and Proline Accumulation of det2-1
To gain some insight into the possible molecular mechanism underlying the salt hypersensitivity of det2-1, the expression in det2-1 of some marker genes known to be induced by salt stress and ABA was compared with that in WT. COR78 and RD22 were reported to be induced by abiotic stress such as salinity, desiccation, cold, and osmotic stress (Zhu 2002; Yamaguchi-Shinozaki and Shinozaki 2006). As shown in Fig. 3a and b, during salt stress the induction of COR78 transcription in det2-1 was significantly decreased compared with that in WT, especially at 6 h after treatment, but the salt-induced RD22 expression was maintained in both genotypes.

Salt-induced gene expression and proline accumulation in det2-1. a RT-PCR analysis of COR78, RD22, and P5CS1 in det2-1 and WT under 200-mM-NaCl stress for 0, 4, and 6 h. ACTIN7 was used as an internal control. b Quantitative results of RT-PCR for transcript level (mean ± SD, n = 3 for COR78 and RD22; n = 4 for P5CS1). c Content and folds of proline accumulation of WT and det2-1 (mean ± SD, n = 5) under 200-mM-NaCl treatment for 24 h with or without 10 nM EBR. CK: seedlings grown on normal 1/2 MS medium
The expression of P5CS1, which encodes the key enzyme of proline biosynthesis, was also examined. The induction of P5CS1 in det2-1 after a 4-h treatment with 200 mM NaCl was greatly inhibited compared with that in WT (Fig. 3a, b). We have further examined the proline content of det2-1 under NaCl treatment and found that the proline level under the stress condition was not significantly different from that of WT. However, the folds of proline accumulation in det2-1 were less than in WT due to a higher basal proline content in det2-1 (Fig. 3c). Our results seem to suggest that endogenous BR may be involved in the salt response by modulating the expression of some stress-inducible genes and flux through the proline biosynthetic pathway.
We also tested whether addition of BR could rescue proline accumulation in det2-1. As demonstrated in Fig. 3c, the basal level of proline in det2-1 was reduced to a level similar to that in WT, which resulted in enhanced proline accumulation.
Hypersensitivity to Salt Stress and Proline Accumulation of bin2-1 Seedling
To obtain more evidence about the involvement of BR in a plant’s salt stress responses, we tested the salt sensitivity of BR-insensitive mutant bin2-1 at the seedling stage. Because the homozygous bin2-1 mutant is sterile, we used the seeds derived from the heterozygous bin2-1, which would segregate out plants corresponding to homozygous bin2-1, heterozygous bin2-1, and WT. Seven days after germination the seedlings of heterozygous bin2-1 phenotype were selected and transferred to 1/2 MS medium containing different NaCl concentrations. Almost all bin2-1 seedlings were bleached at 150 and 200 mM NaCl (Fig. 4a), and root growth was more severely inhibited than in WT at each NaCl concentration tested. The survival rate (Fig. 4b), relative root elongation (Fig. 4c), and fresh weight (Fig. 4d) all showed that heterozygous bin2-1seedlings are more sensitive to salt stress than WT. Similar to the results in det2-1, 200-mM-NaCl-induced P5CS1 upregulation (Fig. 4e, f) and proline accumulation (Fig. 4g) were also inhibited in bin2-1 seedlings.

Altered sensitivity to salt stress and proline accumulation of bin2-1 seedlings. a Growth of bin2-1 seedlings on medium with different NaCl concentrations. Seedlings grown on normal medium for 7 days were transferred onto NaCl-containing medium and grown for another 3 days. Scale bars: 2 cm. Survival rate (b), relative inhibition of root length (c), and fresh weight (d) of bin2-1 (mean ± SD, n = 4) under different concentrations of NaCl. e RT-PCR analysis of P5CS1 transcript level in WT and bin2-1 at 0 h (lanes 1, 4), 4 h control (lanes 2, 5), and 200-mM-NaCl treatment for 4 h (lanes 3, 6), ACTIN7 was used as an internal control. f Quantitative results of RT-PCR for P5CS1 transcript level (mean ± SD, n = 4). g Content and folds of proline accumulation in WT and bin2-1 (mean ± SD, n = 5) treated with 200 mM NaCl or 50 μM ABA. CK: seedlings grown on 1/2 MS medium
Exogenous BR Enhanced the Seedlings Growth and Proline Accumulation Under Salt Stress
The above results indicated that the WT level of endogenous BR and BR signaling are both necessary for normal seedling growth during salt stress. Thus, we further tested whether exogenous applied EBR could improve the growth of WT seedlings under salt stress. Four-day-old wild-type seedlings were grown on medium containing 75 mM NaCl, with or without 0.1 μM EBR for 7 days. The result showed that addition of 0.1 μM EBR in the growth medium under 75 mM NaCl could significantly improve the root elongation of the seedlings, but no improvement was observed at higher BR concentrations (Fig. 5a).

Effect of EBR on seedling root elongation and proline accumulation under salt stress. a Effect of EBR on root length of salt-stressed (75 mM) seedlings (mean ± SD, n = 4); root length was measured at 7 days after stress. b Effect of EBR pretreatment on proline accumulation of WT under 200-mM-NaCl stress (mean ± SD, n = 5). Seedlings were preincubated with different concentrations of EBR for 3 days before addition of 200 mM NaCl; CK: seedlings pretreated with 1/2 MS medium with 70% ethanol added (solvent of EBR) for 3 days
The effect of BR on salt-induced proline accumulation was also analyzed. Ten-day-old wild-type seedlings were first pretreated with 0.1, 0.3, or 0.5 μM of EBR for 3 days before being incubated under 200 mM NaCl. Proline accumulation induced by salt treatment was enhanced by EBR in a dose-dependent manner (Fig. 5b). These results showed that exogenous BR could enhance salt tolerance and salt-induced proline accumulation.
Discussion
Most plant growth hormones are first discovered as substances that stimulate plant development; however, some, such as abscisic acid (Shinozaki and others 2003; Zhang and others 2006; Lin and others 2007), ethylene (Mizoguchi and others 1997; Wang and others 2007), and jasmonic acid (Takahashi and others 2007; Walia and others 2007), are later found to also modulate the plant response to environmental stimuli. Previous reports on BR’s function in plant stress tolerance were based mostly on its exogenous application. The involvement of endogenous BR in stress response can best be investigated using mutants deficient in endogenous BR production or BR signaling. To this end, BR mutants det2-1 and bin2-1 were used to study the function of endogenous BR in salt tolerance. Examination of seed germination rates and measurements of relative root elongation and relative fresh weight of the seedlings demonstrated that det2-1 was hypersensitive to salinity at both germination and the early growth stage, and this growth defect can be partially rescued by the exogenous addition of 10 nM EBR. In addition, the seedling growth of the BR-insensitive mutant bin2-1 also showed hypersensitivity to salt stress, suggesting that endogenous BR and BR signaling are important for salt tolerance of Arabidopsis.
It has been widely reported that there might be an antagonistic interaction between BR and ABA signaling pathways. For example, expression of three closely related BR-responsive bHLH transcription factors, BEE1, BEE2, and BEE3, was inhibited by ABA, and overexpressing BEE1 could reduce ABA sensitivity in Arabidopsis (Friedrichsen and others 2002). bin2-1 displayed a hypersensitivity to ABA in root elongation profiles (Li and others 2001). Recently, it has been reported that ABA could inhibit BR signaling by modulating the phosphorylation status of BES1 and BR-responsive gene expression (Zhang and others 2009).
Altered sensitivity of det2-1 seedlings to salt stress was manifested not only in germination and seedling growth, but also in the expression of genes known to be related to salt response. Salt induction of COR78 and P5CS1 expression is decreased in det2-1, suggesting that BR might promote salt tolerance by upregulating a subset of salt-responsive genes. In a genome-wide search for BR-regulated genes, COR78 is among the few stress-related genes that are responsive to BR, whose expression in the BR-deficient mutant dwf1-6 was found to be slightly decreased compared to that in WT, but downregulated by BR treatment (Müssig and others 2001). Therefore, it is not clear whether BR directly regulates COR78 expression. In our data, the expression of COR78 was undetectable under control conditions in both the wild type and det2-1. However, its salt-induced expression was reduced. We reason from these data that BR may not directly regulate the promoter of COR78 and P5CS1 (which is not on the list of BR-regulated genes in the above-cited article); rather, BR may interact with a salt signaling component to affect the salt-induced expression of COR78 and P5CS1. Another possibility is that BR may regulate COR78 and P5CS1 promoters only under stress conditions. The E-box consensus sequence (CANNTG), present in many BR-induced promoters (Nemhauser and others 2004), was found in the promoter of P5CS1 (at −86 and −128 bp) and COR78 (at −297 bp).
Proline accumulation appears to play a protective role in plants under salt and drought stresses (Savouré and others 1995) and is regulated by signal molecules such as calcium (Knight and others 1997), PLC (Parre and others 2007), and PLD (Thiery and others 2004), which are integrated in ABA-dependent and ABA-independent pathways (Savouré and others 1997; Strizhov and others 1997).
Our results showed that the induction of the P5CS1 transcript level in det2-1 and bin2-1 was attenuated compared with that in WT after treatment with 200 mM NaCl (Figs. 3a, 4e). However, the absolute proline level during stress was not significantly changed or even slightly increased. The decreased proline accumulation is mainly due to the enhanced basal proline level (Figs. 3c, 4g). It is possible that here the P5CS1 expression level is not the sole determinant for the proline level. It seemed that expression of the proline dehydrogenase gene was also decreased in det2-1 compared with that in WT (data not shown). The role of proline accumulation in protecting the stressed plant is a matter of debate. It is not clear whether the role of proline accumulation is achieved by the proline molecule itself or by the proline biosynthetic process, which consumes large amounts of NADPH+ reported to accumulate during many kinds of environmental stresses (Hare and Cress 1997).
On the other hand, exogenous EBR could increase proline accumulation of WT under 200 mM NaCl (Fig. 5b), supporting a previous report that exogenous EBR could enhance proline accumulation induced by 20% PEG treatment in Sorghum (Vardhini and Rao 2003). The proline accumulations of both bin2-1 (Fig. 4g) and det2-1 (data not shown) under 50-μM-ABA treatment were less than that of WT, raising the possibility that decreased proline accumulation in det2-1 and bin2-1 under 200 mM NaCl may result from the antagonistic effect of BR on the ABA-dependent pathway. Taken together, these results suggest that the role of BR in the salt tolerance of Arabidopsis may be accomplished partially by regulating proline accumulation; however, more work needs to be done to elucidate the precise role of BR in stress-induced proline accumulation.
In contrast to our results, Abraham and others (2003) have shown that the induction of P5CS1 transcription after NaCl treatment for 24 h was inhibited by EBR in WT and enhanced in det2-1. The apparent discrepancy may be due to different growth conditions, especially different light regimes. In our experiment, Arabidopsis seedlings were grown under a 16-h light/8-h dark cycle, whereas the seedlings of Abraham and others were grown under an 8-h light/16-h dark cycle. The activity of BR could be affected by light (Neff and others 1999; Turk and others 2005). We have also tried BR treatment under short-day conditions; preliminary results showed that BR could suppress proline accumulation. Different lengths of treatment may also affect the induction of gene expression because P5CS1 has been shown to be mostly induced at the onset of stress, reaching a peak at about 4–8 h after 1% NaCl stress (Savouré and others 1995). All our results on P5CS1 expression shown here were obtained with 4-h NaCl treatment.
In conclusion, we have presented evidence that endogenous BR and BR signaling are important for the response of Arabidopsis to salt stress. BR could enhance salt tolerance by affecting the expression of some stress-related genes and proline accumulation. Our results, therefore, have provided more insights into the role of BR as a plant growth hormone not only in plant development but also in adaptation to environmental stresses.
References
Abraham E, Rigo G, Szekely G, Nagy R, Koncz C, Szabados L (2003) Light-dependent induction of proline biosynthesis by abscisic acid and salt stress is inhibited by brassinosteroid in Arabidopsis. Plant Mol Biol 51:363–372
Anuradha S, Rao SSR (2001) Effect of brassinosteroids on salinity stress induced inhibition of seed germination and seedling growth of rice (Oryza sativa L.). Plant Growth Regul 33:151–153
Anuradha S, Rao SSR (2003) Application of brassinosteroids to rice seeds (Oryza sativa L.) reduced the impact of salt stress on growth, prevented photosynthetic pigment loss and increased nitrate reductase activity. Plant Growth Regul 40:29–32
Bates L, Waldren RP, Teare ID (1973) Rapid determination of free proline for water stress studies. Plant Soil 39:107–205
Clouse SD, Sasse JM (1998) Brassinosteroids: essential regulators of plant growth and development. Annu Rev Plant Physiol Plant Mol Biol 49:427–451
Dhaubhadel S, Browning KS, Gallie DR, Krishna P (2002) Brassinosteroid functions to protect the translational machinery and heat-shock protein synthesis following thermal stress. Plant J 29:681–691
Friedrichsen DM, Nemhauser J, Muramitsu T, Maloof JN, Alonso J, Ecker JR, Furuya M, Chory J (2002) Three redundant brassinosteroid early response genes encode putative bHLH transcription factors required for normal growth. Genetics 162:1445–1456
Hare PD, Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants. Plant Growth Regul 21:79–102
Hayat S, Ali B, Aiman Hasan S, Ahmad A (2007) Brassinosteroid enhanced the level of antioxidants under cadmium stress in Brassica juncea. Environ Exp Bot 60:33–41
Hua XJ, Van De Cotte B, Van Montagu M, Verbruggen N (2001) The 5′ untranslated region of the At-P5R is involved in both transcriptional and post-transcriptional regulation. Plant J 26:157–169
Kagale S, Divi UK, Krochko JE, Keller WA, Krishna P (2007) Brassinosteroid confers tolerance in Arabidopsis thaliana and Brassica napus to a range of abiotic stresses. Planta 225:353–364
Knight H, Trewavas AJ, Knight MR (1997) Calcium signalling in Arabidopsis thaliana responding to drought and salinity. Plant J 12(5):1067–1078
Koh S, Lee SC, Kim MK, Koh JH, Lee S, An G, Choe S, Kim SR (2007) T-DNA tagged knockout mutation of rice OsGSK1, an orthologue of Arabidopsis BIN2, with enhanced tolerance to various abiotic stresses. Plant Mol Biol 65:453–466
Li J, Jin H (2006) Regulation of brassinosteroid signaling. Trends Plant Sci 12:37–41
Li J, Nagpal P, Vitart V, McMorris TC, Chory J (1996) A role for brassinosteroids light-dependent development of Arabidopsis. Science 272:398–401
Li J, Nam KH, Vafeados D, Chory J (2001) BIN2, a new brassinosteroid-insensitive locus in Arabidopsis. Plant Physiol 127:14–22
Lin PC, Hwang SG, Endo A, Okamoto M, Koshib T, Cheng WH (2007) Ectopic expression of ABSCISIC ACID2/GLUCOSE INSENSITIVE1 in Arabidopsis promotes seed dormancy and stress tolerance. Plant Physiol 143:745–758
Mizoguchi T, Ichimura K, Shinozaki K (1997) Environmental stress response in plants: the role of mitogen-activated protein kinases. Trends Biotechnol 15:15–19
Müssig C, Fischer S, Altmann T (2001) Brassinosteroid-regulated gene expression. Plant Physiol 129:1241–1251
Neff MM, Nguyen SM, Malancharuvil E, Fujioka S, Noguchi T, Seto H, Tsubuki M, Honda T, Takatsuto S, Yoshida S, Chory J (1999) BAS1: a gene regulating brassinosteroid levels and light responsiveness in Arabidopsis. Proc Natl Acad Sci USA 96:15316–15323
Nemhauser JL, Mockler TC, Chory J (2004) Interdependency of brassinosteroid and auxin signaling in Arabidopsis. PLOS Biol 2:1460–1471
Ogweno JO, Song XS, Shi K, Hu WH, Mao WH, Zhou YH, Yu JQ, Nogués S (2008) Brassinosteroids alleviate heat-induced inhibition of photosynthesis by increasing carboxylation efficiency and enhancing antioxidant systems in Lycopersicon esculentum. J Plant Growth Regul 27:49–57
Parre E, Ghars MA, Leprince AS, Thiery L, Lefebvre D, Bordenave M, Richard L, Mazars C, Abdelly C, Savouré A (2007) Calcium signaling via phospholipase C is essential for proline accumulation upon ionic but not nonionic hyperosmotic stresses in Arabidopsis. Plant Physiol 144:503–512
Savouré A, Jauoa S, Hua XJ, Ardiles W, van Montague M, Verbruggen N (1995) Isolation, characterization, and chromosomal location of a gene encoding the △1-pyrroline-5-carboxylate synthetase in Arabidopsis thaliana. FEBS Lett 372:13–19
Savouré A, Hua XJ, Bertauche N, Van Montagu M, Verbruggen N (1997) Abscisic acid-independent and abscisic acid-dependent regulation of proline biosynthesis following cold and osmotic stresses in Arabidopsis thaliana. Mol Gen Genet 254:104–109
Shinozaki K, Yamaguchi-Shinozaki K, Sekiz M (2003) Regulatory network of gene expression in the drought and cold stress responses. Curr Opin Plant Biol 6:410–417
Singh I, Shono M (2005) Physiological and molecular effects of 24-epibrassinolide, a brassinosteroid on thermotolerance of tomato. Plant Growth Regul 47:111–119
Steber CM, McCourt P (2001) A role for brassinosteroids in germination in Arabidopsis. Plant Physiol 125:763–769
Strizhov N, Ábraham E, Okresz L, Blickling S, Zilberstein A, Schell J, Koncz C, Szabados L (1997) Differential expression of two P5CS genes controlling proline accumulation during salt-stress requires ABA and is regulated by ABA1, ABI1 and AXR2 in Arabidopsis. Plant J 12:557–569
Takahashi F, Yoshida R, Ichimura K, Mizoguchi T, Seo S, Yonezawa M, Maruyama K, Yamaguchi-Shinozaki K, Shinozaki K (2007) The mitogen-activated protein kinase cascade MKK3–MPK6 is an important part of the jasmonate signal transduction pathway in Arabidopsis. Plant Cell 19:805–818
Thiery L, Leprince AS, Lefebvre D, Ghars MA, Debarbieux E, Savouré A (2004) Phospholipase D is a negative regulator of proline biosynthesis in Arabidopsis thaliana. J Biol Chem 279:14812–14818
Turk EM, Fujioka S, Hideharu S, Shimada Y, Takatsuto S, Yoshida S, Wang H, Torres QI, Ward JM, Murthy G, Zhang J, Walker JC, Neff MM (2005) BAS1 and SOB7 act redundantly to modulate Arabidopsis photomorphogenesis via unique brassinosteroid inactivation mechamisms. Plant J 42:23–43
Vardhini BV, Rao SSR (2003) Amelioration of osmotic stress by brassinosteroids on seed germination and seedling growth of three varieties of sorghum. Plant Growth Regul 41:25–31
Walia H, Wilson C, Condamine P, Liu X, Ismail AM, Close TJ (2007) Large-scale expression profiling and physiological characterization of jasmonic acid-mediated adaptation of barley to salinity stress. Plant Cell Environ 30:410–421
Wang ZY, He JX (2004) Brassinosteroid signal transduction—choices of signals and receptors. Trends Plant Sci 9:92–96
Wang Y, Liu C, Li K, Sun F, Hu H, Li X, Zhao Y, Han C, Zhang W, Duan Y, Liu M, Li X (2007) Arabidopsis EIN2 modulates stress response through abscisic acid response pathway. Plant Mol Biol 64:633–644
Yamaguchi-Shinozaki K, Shinozaki K (2006) Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu Rev Plant Biol 57:781–803
Zhang J, Jia W, Yang J, Ismail AM (2006) Role of ABA in integrating plant responses to drought and salt stresses. Field Crops Res 97:111–119
Zhang S, Cai Z, Wang X (2009) The primary signaling outputs of brassinosteroids are regulated by abscisic acid signaling. Proc Natl Acad Sci U S A 106:4543–4548
Zhu JK (2002) Salt and drought stress signal transduction in plants. Annu Rev Plant Biol 53:247–273
Acknowledgments
The authors are grateful to Dr. Zhiyong Wang (Stanford University) and Dr. Jun Zhao (The Chinese Academy of Agriculture Sciences) for providing Arabidopsis mutant seeds. This work was supported by the Chinese National Key Basic Research Project (#2006CB100100) from the Ministry of Science and Technology of China and the Hundred Talent Program of the Chinese Academy of Science.
Author information
Authors and Affiliations
Corresponding author
Additional information
Haitao Zeng and Qi Tang contributed equally to this work.
Rights and permissions
About this article
Cite this article
Zeng, H., Tang, Q. & Hua, X. Arabidopsis Brassinosteroid Mutants det2-1 and bin2-1 Display Altered Salt Tolerance. J Plant Growth Regul 29, 44–52 (2010). https://doi.org/10.1007/s00344-009-9111-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00344-009-9111-x