
Abstract
Coral larvae acquire populations of the symbiotic dinoflagellate Symbiodinium from the external environment (horizontal acquisition) or inherit their symbionts from the parent colony (maternal or vertical acquisition). The effect of the symbiont acquisition strategy on Symbiodinium-host associations has not been fully resolved. Previous studies have provided mixed results, probably due to factors such as low sample replication of Symbiodinium from a single coral host, biogeographic differences in Symbiodinium diversity, and the presence of some apparently host-specific symbiont lineages in coral with either symbiont acquisition strategies. This study set out to assess the effect of the symbiont acquisition strategy by sampling Symbiodinium from 10 coral species (five with a horizontal and five with a vertical symbiont acquisition strategy) across two adjacent reefs in the southern Great Barrier Reef. Symbiodinium diversity was assessed using single-stranded conformational polymorphism of partial nuclear large subunit rDNA and denaturing gradient gel electrophoresis of the internal transcribed spacer 2 region. The Symbiodinium population in hosts with a vertical symbiont acquisition strategy partitioned according to coral species, while hosts with a horizontal symbiont acquisition strategy shared a common symbiont type across the two reef environments. Comparative analysis of existing data from the southern Great Barrier Reef found that the majority of corals with a vertical symbiont acquisition strategy associated with distinct species- or genus-specific Symbiodinium lineages, but some could also associate with symbiont types that were more commonly found in hosts with a horizontal symbiont acquisition strategy.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Symbiodinium is a photosynthetic dinoflagellate genus that is found in symbiosis with some protists and a wide range of marine invertebrate hosts, including reef-building corals (Baker 2003). The mutualistic symbiosis between Symbiodinium and corals is considered to be responsible for the growth and formation of coral reefs through the energetic and nutritional advantages of algal–animal endosymbiosis (Muscatine et al. 1981).
There are eight divergent lineages within the genus Symbiodinium, designated clades A–H, which each contain numerous subclade types based on rDNA (reviewed in Coffroth and Santos 2005; Stat et al. 2006). Patterns of phylogeography are evident for Symbiodinium at the subclade level. Adaptive radiation and limited connectivity have been proposed to explain the large differences in Symbiodinium diversity seen between reefs in the Indo-Pacific compared to the Atlantic–Caribbean (LaJeunesse 2005). On smaller scales, different host–symbiont assemblages can be seen over latitudinal and longitudinal gradients (Baker et al. 1997; Rodriguez-Lanetty et al. 2001; LaJeunesse et al. 2004a, b; LaJeunesse 2005), and as a function of colony depth (Rowan and Knowlton 1995; Baker and Rowan 1997; Toller et al. 2001; LaJeunesse et al. 2003; Iglesias-Prieto et al. 2004) and the irradiance level in individual colonies (Rowan et al. 1997; van Oppen et al. 2001).
Coral hosts acquire Symbiodinium each generation, either by the dinoflagellate symbiont infecting juvenile corals from the ocean environment (horizontal acquisition; Trench 1987) or directly from the parent egg or brooded larvae (maternal or vertical acquisition; Trench 1987). There are conflicting data as to whether this symbiont acquisition strategy influences the final host–symbiont assemblage. Analysis of coral hosts and their associated Symbiodinium genotypes based on the internal transcribed spacer 2 (ITS2) region suggests a correlation between “specialist” Symbiodinium types and hosts with a vertical symbiont acquisition strategy (e.g., LaJeunesse et al. 2003, 2004a, b; LaJeunesse 2005; Thornhill et al. 2006a, b). A high number of unique symbiont types were found in coral hosts from Hawaii (LaJeunesse et al. 2004a), which was considered to be due to the predominance of coral hosts with a maternal symbiont acquisition strategy in Hawaiian reefs. Loh et al. (2001) showed that Seriatopora hystrix (maternal strategist) harbored unique symbiont types in different geographic locations within the Pacific, while Acropora longicyathus (horizontal strategist) had symbionts that were genetically identical between reefs from Australia, Malaysia, and Japan. A study by Barneah et al. (2004) on soft corals from the Red Sea found clade A symbionts associated with hosts that have a vertical symbiont acquisition strategy while clade C associated with horizontal strategists. In contrast to these observations that largely show symbionts to partition according to host symbiont acquisition strategy, van Oppen (2004) found a similar level of symbiont genetic diversity in Acropora sp. (horizontal strategist) and Montipora sp. (maternal strategist) from the central GBR. Montipora associated with some unique symbiont types inferred to have co-evolved with the host but also harbored symbionts commonly found in Acropora. Other studies employing different coral hosts have found some flexibility among maternal strategists, which sometimes harbor generalist types that are usually confined to symbiosis with horizontal strategists (e.g., Sampayo et al. 2007). In addition, some horizontal strategists can show evidence of specificity, as was observed in Fungia scutaria from Hawaii, which acquires its symbionts from the environment but was found to house the specialist Symbiodinium genotype ITS2 C1f (Weis et al. 2001; Rodriguez-Lanetty et al. 2004).
The difficulty of resolving the effect of symbiont acquisition strategy on Symbiodinium diversity lies in the fact that the above studies did not set out to specifically address this issue or investigate the question using a variety of host species representing both modes of symbiont acquisition. The aim of this study was therefore to systematically sample 10 coral species, five with a horizontal and five with a vertical symbiont acquisition strategy, from two separate locations in the southern Great Barrier Reef, to determine how host symbiont acquisition strategy affects the diversity of their constituent Symbiodinium types.
Materials and methods
Sample collection
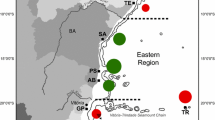
Samples were collected in August 2001 from 10 coral species (14–22 colonies/species, Table 1) at various reef locations surrounding One Tree Island (23°25′ S 151°55′ E) and Heron Island (23°25′ S 151°55′ E) located in the southern Great Barrier Reef (Fig. 1). Exceptions were colonies of S. hystrix collected from location B in January/February 2002 and colonies of Montipora digitata that were collected from location E in July 2002. Coral species that were abundant on these reefs and well characterized with respect to lifecycle were targeted. Conspecific colonies separated by at least 10 m (to reduce the possibility of collecting the same asexually produced individual) were sampled from intertidal reef flat zones and at depths of 0–4 and 12–15 m. A 3-cm piece from branching species or a 3-cm2 section from massive species was removed using a hammer and chisel. Samples were collected from three sections of each coral colony according to the level of light irradiance (low, intermediate, high) categorized by position on the colony and relative amount of shading. Coral tissue was removed from the skeleton with directed jets of high pressure air supplied using SCUBA, collected into sterile bags, and mixed with an equal amount of saturated salt-DMSO preserving buffer in a microfuge tube for transport and storage (Seutin et al. 1991).

Location of sites where corals were sampled for Symbiodinium genotyping. (a) The location of sampling sites within the Great Barrier Reef; (b) position of reefs within the Capricorn Bunker Group; (c) Heron Island reef sites A: intertidal reef flat, B: 0–4 m, C: 12–15 m, D: 12–15 m; (d) One Tree reef sites E: 0–4 m, F: 0–4 m, G: 0–4 m, H: 12–15 m, I: 12–15 m, J: 0–4 m
DNA extraction
Total DNA was extracted using the protocol described in Loh et al. (2001) with minor modifications. Preserved coral/algal tissue was incubated with 1% SDS at 65°C for 1 h followed by overnight incubation at 50°C with the addition of Proteinase K (0.5 mg ml−1). Cellular debris was removed by extraction with phenol–chloroform–isoamyl alcohol (25:24:1) followed by centrifugation. The supernatant was heated at 65°C for 15 min with 1% CTAB and re-centrifuged. An equal volume of chloroform–isoamyl alcohol (24:1) was then added to the supernatant, which was chilled on ice for 30 min and centrifuged again. DNA was precipitated from the final supernatant with an equal volume of isopropanol and 1/10 volume 3 M sodium acetate (pH 5.2). The DNA was washed in 70% ethanol, dried, and resuspended in TE Buffer (10 mM Tris–Borate, 1 μM EDTA, pH 8.0).
LSU amplification, SSCP, and sequence analysis
Dinoflagellate specific primers were used to amplify the D1/D2 region of the LSU rDNA gene from Symbiodinium (Zardoya et al. 1995; forward: 5′ CCC GCT GAA TTT AAG CAT ATA AGT AAG CGG 3′ and reverse: 5′ GTT AGA CTC CTT GGT CCG TGT TTC AAG A 3′). All PCR reactions contained 10 μl 10× PCR buffer (1 M Tris–HCl, 0.5 M KCl, 2.5 mM MgCl2, pH 8.3), 1 μl of 5 μM each dNTP, 40 μg bovine serum albumin, 20 pmol of each primer, and 0.5 μl of Taq DNA polymerase (5 unit μl−1) in a 100-μl reaction. PCR reactions were performed using the following conditions: 94°C for 5 min, followed by 30 cycles of 94°C for 1 min, 65°C for 40 s, and 72°C for 40 s, and a final extension at 72°C for 7 min. Partial LSU rDNA PCR product (20 μl) was mixed with an equal volume of stop solution (95% formamide, 10 mM NaOH, 0.25% bromophenol blue, and 0.25% xylene cyanol), heated at 95°C for 8 min and immediately chilled on ice for 10 min. A 0.5× MDE vertical gel, prepared according to the manufacturer’s instructions (BioWhittaker Molecular Applications, Rockland, ME, USA), was loaded with the samples and electrophoresed at 140 V for 14 h with 0.6% TBE running buffer (45 mM Tris–Borate, 1 mM EDTA, pH 8.0). The gel was stained in Syber gold (Molecular Probes, Eugene, OR, USA) for 20 min and viewed under UV transillumination. The image was captured on Polaroid 667 film. Single-stranded conformational polymorphism (SSCP) profiles were compared visually, and variants were selected and electrophoresed together on a single SSCP gel for final comparison. This was done to prevent gel-to-gel variation in SSCP profiles that can occur due to minor differences in gel composition or ambient temperature. All amplifications that gave unique SSCP profiles were directly sequenced. The partial LSU rDNA fragments were purified using GFX™ PCR DNA purification kits (Amersham Biosciences, New Jersey USA) prior to sequencing on an ABI 3730xl 96-capillary automated DNA sequencer at the Australian Genome Research Facility. DNA sequences were aligned using Clustal W (Thompson et al. 1994) and manually edited using GeneDoc (Nicholas et al. 1997).
ITS2 amplification and DGGE
ITS2 analysis was undertaken to integrate the data presented in this study with the extensive ITS2 database (e.g., LaJeunesse 2001, 2002, 2005; LaJeunesse et al. 2003, 2004a, b; Thornhill et al. 2006a, b; Pochon et al. 2007; Sampayo et al. 2007) and to compare the diversity and population structure of Symbiodinium between the two genes. The ITS2 region was amplified as described in LaJeunesse and Trench (2000). Touchdown PCR using the primers ITS2 clamp (5′ CGC CCG CCG CGC CCC GCG CCC GTC CCG CCG CCC CCG CCC GGG ATC CAT ATG CTT AAG TTC AGC GGG T 3′) and ITSintfor2 (5′ GAA TTG CAG AAC TCC GTG 3′) was performed starting with an annealing temperature of 62°C, which was decreased by 0.5°C per cycle to 52°C and then held for an additional 20 cycles. For denaturing gradient gel electrophoresis (DGGE) analysis, 10 μl of ITS2 PCR product was loaded onto an 8% polyacrylamide denaturing gradient gel (45–80% urea–formamide gradient; 100% consists of 7 mol l−1 urea and 40% deionized formamide) and separated by electrophoresis for 9.5 h at 150 V at a constant temperature of 60°C (C.B.S. Scientific, California USA; LaJeunesse 2002). Profiles were compared to reference samples of known ITS2 types following the protocol of LaJeunesse and Trench (2000), and representative bands of an ITS2 type were confirmed by sequencing following the protocol of LaJeunesse (2002) on a Biodsystems ABI Prism 377™ at the University of Georgia. The gel was stained with Sybr Green (Molecular Probes) for 25 min according to the manufacturer’s directions and photographed using 667 Polaroid film.
Statistical analyses
Analysis of molecular variance (AMOVA) using conventional F-statistics was implemented using the software Arlequin (Excoffier et al. 2005). AMOVA was used to determine the partitioning of SSCP types in (1) hosts with a vertical symbiont acquisition strategy and in (2) hosts with a horizontal symbiont acquisition strategy.
Results
SSCP and sequence analysis of the partial LSU rDNA region from Symbiodinium
Amplification of the LSU D1/D2 region produced a single fragment of approximately 630 nucleotides from each coral sample. Fifteen different SSCP profiles were identified among the 549 samples (three samples from each of 183 colonies; Fig. 2a). Sequence analysis characterized these as genotypes within Symbiodinium clade C and clade D, and the profiles were designated Clsu1–Clsu13 and Dlsu1–Dlsu2, accordingly. Replicate sequencing was done on SSCP profiles that appeared to be the same but were found in different host colonies or species to confirm that they had the same sequence. This resulted in a total of 23 replicate sequences for Clsu3 (EF372063), 9 for Clsu4 (EF372065), 5 for Clsu5 (EF372066), 6 for Clsu10 (EF372070), and 2 each for Clsu6 (EF372067), Clsu7 (EF372068), and Clsu8 (EF372069). Unambiguous sequence data were obtained from SSCP profiles Clsu4, Clsu5, Clsu7, Clsu8, Clsu10, Dlsu1 (EF372071), and Dlsu2 (EF372072). One sequence originating from the Clsu3 profile amplified from G. favulus differed by a single base change to other sequences derived from this profile, and Clsu6 contained three polymorphic sites. Unambiguous sequences could not be obtained for Clsu1, Clsu2, Clsu9, Clsu11, Clsu12, and Clsu13, indicating multiple symbiont genotypes or a relatively equal abundance of more than one dominant LSU rDNA sequence within the genome of the symbiont (e.g. Santos et al. 2001; LaJeunesse 2002; Diekmann et al. 2003; van Oppen et al. 2005; Thornhill et al. 2007).

Single-stranded conformational polymorphism gel showing Symbiodinium partial nuclear large subunit rDNA fingerprints. Clsu1–Clsu13 and Dlsu1–Dlsu2 represent Symbiodinium genotypes within the phylogenetic groups clades C and D, respectively (a). Clsu11 is distinguished from Clsu10 by two small bands, each indicated by an asterisk. Similarly, Dlsu1 and Dlsu2 are distinguished by two small bands, each indicated by an asterisk. SSCP profiles Clsu10 and Clsu11 originating from the same colony of Montipora digitata sampled at an area of high (1), intermediate (2), and low (3) irradiance (b). Asterisks indicate bands associated with SSCP type Clsu11. M: marker
Diversity and distribution of Symbiodinium
Table 1 shows the Symbiodinium SSCP types associated with each of the coral species at the various different locations, and Fig. 3 represents the SSCP types associated with each host species. Clsu3 was the most abundant, occurring in 72 different colonies and at all of the reef sites except Heron Island site B. Although Clsu3 occurred in all hosts with a horizontal symbiont transmission strategy, it was not seen in any of the hosts with maternal symbiont transmission. Clsu1, Clsu2, Dlsu1, and Dlsu2 also occurred in hosts with a horizontal symbiont transmission strategy only. Clsu1 was common to the massive species Favites abdita (n = 3), Goniastrea favulus (n = 1), and Lobophyllia corymbosa (n = 7) and was found in colonies located at all depths and at both reefs. Dlsu1 was found in a single colony of G. favulus from the Heron Island reef flat (site A). A single colony of Acropora millepora from One Tree Island reef site H contained Clsu2, and two colonies of Acropora palifera at Heron Island reef contained Dlsu2.

Symbiodinium genotypes associated with each of the coral hosts sampled in the study based on LSU rDNA analysis. Each segment within the pie chart represents one coral. Multi-colored pie segments indicate a colony that associated with two SSCP rDNA Symbiodinium types: Clsu10/11 in Montipora digitata and Clsu3/Dlsu2 in Acropora palifera
Unique SSCP types were found in nearly all of the coral hosts with a maternal transmission strategy. The exception was Clsu10, which associated with both M. digitata (n = 17) and Porites cylindrica (n = 17). Clsu10 was the only symbiont type found in these host species, apart from a single colony of M. digitata that contained both Clsu10 and Clsu11 (see below). All colonies of S. hystrix (n = 20) harbored Clsu9. Pocillopora damicornis colonies present at all depths harbored Clsu12 (n = 14), and six colonies at 12–15 m contained Clsu13. Stylophora pistillata associated with five SSCP types: Clsu4 (n = 10), Clsu5 (n = 5), Clsu6 (n = 2), Clsu7 (n = 1), and Clsu8 (n = 1). Clsu4 was present in colonies located at depths of 0–4 and 12–15 m at both reefs. Clsu5 was only found in colonies on the reef flat at Heron Island. Clsu6 occurred in two colonies in shallow water (0–4 m) at One Tree Island, and Clsu7 and Clsu8 were each found in a single colony at Heron Island reef site C.
Samples obtained from single coral colonies produced identical SSCP profiles regardless of whether they were exposed to high, medium, or low irradiance within the colony. The exception was a single colony of M. digitata that contained both Clsu10 and Clsu11 (Fig. 2b). In this colony, Clsu10 appeared dominant at the region of highest irradiance on the colony; an equal abundance of Clsu10 and Clsu11 occurred at intermediate irradiance; and Clsu11 dominated at the region of lowest irradiance. One colony of A. palifera contained Clsu3 and Dlsu2, but these were equally present throughout the coral colony.
DGGE analysis of Symbiodinium ITS2
The ITS2 region was amplified from a subset of the samples and the diversity assessed using DGGE, allowing the integration of previously published ITS2 genotypes and data from the current study. Figure 4 shows representative ITS2-DGGE types identified in the study. Samples typed using DGGE of the ITS2 region included Clus1 (from G. favulus: n = 1 and L. corymbosa: n = 1), Clsu3 (from A. millepora: n = 13, A. palifera: n = 15, F. abdita: n = 3, G. favulus: n = 2, and L. corymbosa: n = 3), Clsu4 (from S. pistillata: n = 3), Clsu5 (from S. pistillata: n = 3), Clsu6 (from S. pistillata: n = 2), Clsu7 (from S. pistillata: n = 1), Clsu8 (from S. pistillata; n = 1), Clsu9 (from S. hytrix: n = 13), Clsu10 (from M. digitata: n = 14, and P. cylindrica: n = 17), Clsu11 (from M. digitata: n = 1), Clsu12 (from P. damicornis: n = 3), Clsu13 (from P. damicornis: n = 3), Dlsu1 (from G. favulus: n = 1), and Dlsu2 (from A. palifera: n = 2). DGGE profiles correlated with SSCP profiles so that a single DGGE profile was observed for all samples producing a given SSCP profile. Exceptions were Clsu12 which produced the ITS2-DGGE patterns representing C42/C42a and C42/C42a/C42b, and Clsu5 and Clsu6 which both gave ITS2 pattern C78.

Internal transcribed sequence 2 (ITS2) DGGE profiles of Symbiodinium identified in the coral hosts sampled in the study. The corresponding LSU rDNA type is indicated at the top of each lane. M: ITS2-DGGE standards
Community structure of Symbiodinium
Partitioning of LSU SSCP types according to host symbiont acquisition strategy was found to be highly significant (Likelihood ratio test; 255.81, df = 14, P < 0.0001). There was a significant association between coral species with a vertical symbiont acquisition strategy and the associated symbiont SSCP type (Likelihood ratio test; 267.43, df = 36, P < 0.0001) indicating host–symbiont specificity within this group of corals. In contrast, there was no significant association between coral species with a horizontal symbiont acquisition strategy and the associated symbiont SSCP type, indicating a shared symbiont type for this group of corals.
Symbiodinium SSCP types were grouped by host and reef for AMOVA (Table 2). Two independent AMOVAs were performed: (1) for hosts with a vertical symbiont acquisition strategy and (2) for hosts with a horizontal symbiont acquisition strategy. In hosts with a vertical symbiont acquisition strategy, there was a significant effect of host on the community of Symbiodinium (ϕCT = 0.712, P = 0.002, Table 2) and a significant effect of reef nested within host (ϕSC = 0.140, P = 0.036, Table 2). In contrast, for coral with a horizontal symbiont acquisition strategy, partitioning of Symbiodinium due to host or by reef nested within host was not significant (Table 2).
Discussion
This study supports the hypothesis that the mechanism by which corals obtain their symbionts strongly influences host–symbiont assemblages. Symbiodinium genotypes from corals with a maternal symbiont acquisition strategy were largely host-specific, while all corals included with horizontal symbiont acquisition strategies shared a lower diversity of symbiont types.
Although hosts employing a maternal symbiont acquisition strategy all harbored distinct Symbiodinium genotypes, differences in specificity and diversity were evident within this group of corals. M. digitata and P. cylindrica both harbored almost exclusively Clsu10 (ITS2 C15), a symbiont specific to these corals and found in several other species of Porites and the hydroid Aglaophenia sp. in the region (this study; LaJeunesse et al. 2003). M. digitata also harbored Clsu11 (ITS2 C73), which has not been found in any other coral species (LaJeunesse 2005). S. hystrix, harbored Clsu9 (ITS2 C3n-t) exclusively, although it has also been shown to associate with the global generalist symbiont ITS2 C3 when sampled in this region (LaJeunesse et al. 2003; Sampayo et al. 2007). Numerous Symbiodinium types were found in S. pistillata and P. damicornis. S. pistillata harbored symbiont types Clsu4 (ITS2 C8/C8a), Clsu5 (ITS2 C78), Clsu6 (ITS2 C78), Clsu7 (ITS2 C35/C35a), and Clsu8 (ITS2 C79). In addition to these, the global generalist symbiont ITS2 C1 has been detected in colonies of S. pistillata (LaJeunesse et al. 2003; Sampayo et al. 2007). Similarly, in addition to symbiont types Clsu12 (ITS2 C42/C42a/C42b) and Clsu13 (ITS2 C33) found in P. damicornis, the global generalist C1 and other symbionts not specific to P. damicornis (ITS2 C1b and C1c) have been found in this vertical strategist (LaJeunesse et al. 2003; Sampayo et al. 2007). A faint band at the migration endpoint for ITS2 C1 in our DGGE profiles for Clsu12/13 also points to the presence of C1 in this host species. Collectively, these findings show that host-specific symbiont lineages predominate in coral with a vertical symbiont acquisition strategy in the southern Great Barrier Reef and that generalist symbionts can also associate with these corals, as has been suggested by van Oppen (2004). ITS2 types C3 and C1 have been called the “living ancestors” within the Symbiodinium clade C lineage (LaJeunesse 2005). Thus, even though host–symbiont specificity has evolved in coral lineages with a vertical symbiont acquisition strategy, the recognition between these ancestral types (from which the specialist symbionts are hypothesized to have evolved) and this group of corals appears to remain a viable interaction.
Genotypes Clsu1 (ITS2 C21) and Clsu3 (ITS2 C3) were common to hosts with horizontal symbiont acquisition strategies and were truly ubiquitous, being present in corals at both study sites and at various depths. It is likely that retaining the ability to maintain symbiotic interactions with a wide range of host species and to persist when outside the host has restricted the genetic differentiation, and hence speciation of these horizontally transmitted symbionts. Although host-specific symbiont lineages were not detected in this group of corals in the current study, specificity between the horizontal strategist F. scutaria and the specialist genotype ITS2 C1f has been shown in coral originating from Hawaii (Weis et al. 2001; Rodriguez-Lanetty et al. 2004). This suggests that specific host–symbiont assemblages can also occur in corals that acquire their symbionts from the environment so that a particular symbiont is selected over the generalist type (e.g., C1 and/or C3), but this is apparently a rare scenario.
Spatial distribution of different symbiont types within a coral colony and with colony depth was rare. A single colony of A. palifera harbored both Clsu3 (ITS2 C3) and Dlsu2 (ITS2 D1) and one colony of M. digitata harbored Clsu10 (ITS2 C15) and Clsu11 (ITS2 C73), with the latter appearing to be distributed over the colony according to relative light levels. The ability to accommodate symbionts that are genetically variable may provide the host with benefit over a wider environmental range (Douglas 1998). However, P. damicornis and S. pistillata appear to be more susceptible to bleaching than M. digitata and P. cylindrica, which associate with a single Symbiodinium genotype but are relatively bleach resistant (Loya et al. 2001; LaJeunesse et al. 2003). Recently, Mieog et al. (2007) found colonies of Acropora contained multiple symbiont types that could not be detected using standard PCR and fingerprinting techniques. It is possible that this finding may extend to many more host taxa. These cryptic symbiont populations may be kept at low levels, as mixed Symbiodinium communities have been shown to be less beneficial to the host animal (Loram et al. 2007).
The LSU rDNA and ITS2 regions are linked in the rDNA array (Long and Dawid 1980). ITS2 is under less functional constraint than the LSU region and is therefore likely to be more variable (LaJeunesse 2001). The level of Symbiodinium diversity resolved in this study using these two markers was similar, and the few exceptions could be the result of primer bias, or because different techniques were used to analyze the diversity of the amplified fragments. The difference in Symbiodinium diversity resolved in this study compared to others performed in the Southern Great Barrier Reef, and particularly the lack of generalist types seen among the vertical strategists in our analysis compared to others (LaJeunesse et al. 2003; Sampayo et al. 2007) is likely due to differences in the number of colonies sampled in the study (where increased sampling can reveal rare host–symbiont partnerships; e.g., Sampayo et al. 2007), the site at which the colony was collected (fine-scale niche partitioning; e.g., Sampayo et al. (2007)), or the multi-copy nature of rDNA and interpretation of DGGE gels. For example, the ITS2-DGGE profile for Clsu9 contained bands C3, C3n, and C3t. Are these all from the same genome or do they represent different symbiont types? In addition, some bands were fainter than others within a single profile, such as the C1 band present in the Clsu12/13 samples. Researchers may differ in how they interpret the significance of these minor bands and whether they warrant inclusion in the analysis, which affects the number of symbiont types inferred to be present in a sample. The challenges presented by the multi-copy nature of the ITS2 gene and using DGGE to interpret Symbiodinium diversity have previously been recognized (Apprill and Gates 2007; Thornhill et al. 2007) and argue for the development of new approaches such as multilocus sequence typing (MLST; e.g., Adiri et al. 2003), based on a suite of single copy loci, to characterize functional groups of Symbiodinium.
Mutualisms employing vertical transmission strategies may be more beneficial given that a host with a horizontal acquisition system may fail to acquire symbionts (Genkai-Kato and Yamamura 1999). Indeed, the evolution of the mutualistic symbiosis between macrotermitine termites and their ectosymbiotic fungi reveals vertical transmission as the derived transmission strategy (Aanen et al. 2002). The localization of Symbiodinium during the development from the host egg to mature planulae is different between species, suggesting that vertical symbiont acquisition has evolved multiple times (Hirose and Hidaka 2006). If vertical symbiont acquisition is a more risk-free approach to acquiring symbionts, then why do the majority of coral species possess a horizontal mode of symbiont acquisition? One argument is that horizontal symbiont acquisition allows plasticity in the symbiosis, where hosts may acquire different genetic varieties of symbionts and occupy numerous environmental niches or adapt with environmental change (Buddemeier and Fautin 1993; Douglas 1998). The ability to “switch” symbionts has been proposed (Buddemeier and Fautin 1993; Baker 2001; Baker et al. 2004), but evidence in support of the acquisition of completely new symbiont types in natural coral ecosystems over ecological time scales is currently absent (Hoegh-Guldberg et al. 2002; Stat et al. 2006). If acquisition of exogenous symbionts can occur in adult colonies, the data presented here indicate acquisition of generalist types may be possible for most coral species, but specialist symbionts (which make up the majority of diversity within Symbiodinium) are restricted to their coral lineage. Ultimately, questions on the degree of host–symbiont flexibility and the adaptive potential of coral will become increasingly important, as we strive to understand and project the changes of coral reefs to global climate change.
References
Aanen DK, Eggleton P, Rouland-Lefevre C, Guldberg-Froslev T, Rosendahl S, Boomsma JJ (2002) The evolution of fungus-growing termites and their mutualistic fungal symbionts. Proc Natl Acad Sci USA 99:14887–14892
Adiri RS, Gophna U, Ron EZ (2003) Multilocus sequence typing (MLST) of Escherichia coli 078 strains. FEMS Microbiol Lett 222:199–203
Apprill AM, Gates RD (2007) Recognizing the diversity in coral symbiotic dinoflagellate communities. Mol Ecol 16:1127–1134
Baker AC (2001) Reef corals bleach to survive change. Nature 411:765–766
Baker A (2003) Flexibility and specificity in coral-algal symbiosis: diversity, ecology, and biogeography. Annual Review of Ecology, Evolution, and Systematics 34:661–689
Baker AC, Rowan R (1997) Diversity of symbiotic dinoflagellates (zooxanthellae) in scleractinian corals of the Caribbean and Eastern Pacific. Proc 8th Intl Coral Reef Symp 2:1301–1306
Baker AC, Rowan R, Knowlton N (1997) Symbiosis ecology of two Caribbean Acroporid corals. Proc 8th Intl Coral Reef Symp 2:1295–1300
Baker AC, Starger CJ, McClanahan TR, Glynn PW (2004) Corals’ adaptive response to climate change. Nature 430:741
Barneah O, Weis VM, Perez S, Benayahu Y (2004) Diversity of dinoflagellate symbionts in Red Sea soft corals: mode of symbiont acquisition matters. Mar Ecol Prog Ser 275:89–95
Buddemeier R, Fautin D (1993) Coral bleaching as an adaptive mechanism. Bioscience 43:320–325
Coffroth MA, Santos SR (2005) Genetic diversity of symbiotic dinoflagellates in the genus Symbiodinium. Protist 156:19–34
Diekmann OE, Olsen JL, Stam WT, Bak RPM (2003) Genetic variation within Symbiodinium clade B from the coral genus Madracis in the Caribbean (Netherlands Antilles). Coral Reefs 22:29–33
Douglas A (1998) Host benefit and the evolution of specialization in symbiosis. Heredity 81:599–603
Excoffier L, Laval G, Schneider S (2005) Arlequin ver 3.0: an integrated software package for population genetic data analysis. Evol Bioinformatics Online 1:47–50
Genkai-Kato M, Yamamura N (1999) Evolution of mutualistic symbiosis without vertical transmission. Theor Popul Biol 5:309–323
Hirose M, Hidaka M (2006) Early development of zooxanthella-containing eggs of the corals Porites cylindrica and Montipora digitata: the endodermal localization of zooxanthellae. Zool Sci 23:873–881
Hoegh-Guldberg O, Jones RJ, Ward S, Loh WK (2002) Is coral bleaching really adaptive? Nature 415:601–602
Iglesias-Prieto R, Beltran VH, LaJeunesse TC, Reyes-Bonilla H, Thome PE (2004) Different algal symbionts explain the vertical distribution of dominant reef corals in the eastern Pacific. Proc R Soc Lond, B 271:1757–1763
LaJeunesse TC (2001) Investigating the biodiversity, ecology, and phylogeny of endosymbiotic dinoflagellates in the genus Symbiodinium using the ITS region: in search of a “species” level marker. J Phycol 37:866–880
LaJeunesse TC (2002) Diversity and community structure of symbiotic dinoflagellates from Caribbean coral reefs. Mar Biol 141:387–400
LaJeunesse TC (2005) “Species” radiations of symbiotic dinoflagellates in the Atlantic and Indo-Pacific since the Miocene-Pliocene Transition. Mol Biol Evol 22:570–581
LaJeunesse TC, Trench RK (2000) Biogeography of two species of Symbiodinium (Freudenthal) inhabiting the intertidal sea anemone Anthopleura elegantissima (Brandt). Biol Bull 199:126–134
LaJeunesse TC, Loh WKW, van Woesik R, Hoegh-Guldberg O, Schmidt GW, Fitt WK (2003) Low symbiont diversity in southern Great Barrier Reef corals, relative to those of the Caribbean. Limnol Oceanogr 48:2046–2054
LaJeunesse TC, Thornhill DJ, Cox EF, Stanton FG, Fitt WK, Schmidt GW (2004a) High diversity and host specificity observed among symbiotic dinoflagellates in reef communities from Hawaii. Coral Reefs 23:596–603
LaJeunesse TC, Bhagooli R, Hidaka M, de Vantier L, Done T, Schmidt GW, Fitt WK, Hoegh-Guldberg O (2004b) Closely related Symbiodinium spp. differ in relative dominance in coral reef host communities across environmental, latitudinal and biogeographic gradients. Mar Ecol Prog Ser 284:147–161
Loh WKW, Loi T, Carter D, Hoegh-Guldberg O (2001) Genetic variability of the symbiotic dinoflagellates from the wide ranging coral species Seriatopora hystrix and Acropora longicyathus in the Indo-West Pacific. Mar Ecol Prog Ser 222:97–107
Long EO, Dawid IB (1980) Repeated genes in eukaryotes. Annu Rev Biochem 49:727–764
Loram JE, Trapido-Rosenthal H, Douglas AE (2007) Functional significance of genetically different symbiotic algae Symbiodinium in a coral reef symbiosis. Mol Ecol 16:4849–4857
Loya Y, Sakai K, Yamazato K, Nakano Y, Sambali H, van Woesik R (2001) Coral bleaching: the winners and the losers. Ecol Lett 4:122–131
Mieog JC, van Oppen JH, Cantin NE, Stam WT, Olsen JL (2007) Real-time PCR reveals a high incidence of Symbiodinium clade D at low levels in four scleractinian corals across the Great Barrier Reef: implications for symbiont shuffling. Coral Reefs 26:449–457
Muscatine L, McCloskey LR, Marian RE (1981) Estimating the daily contribution of carbon from zooxanthellae to coral animal respiration. Limnol Oceanogr 26:601–611
Nicholas KB, Nicholas HB Jr, Deerfield DW II (1997) GeneDoc: analysis and visualization of genetic variation. EMBNEW.NEWS 4:14, http://www.nrbsc.org/downloads/
Pochon X, Garcia-Cuetos L, Baker AC, Castella E, Pawlowski J (2007) One-year survey of a single Micronesian reef reveals extraordinarily rich diversity of Symbiodinium types in soritid foramnifera. Coral Reefs 26:867–882
Rodriguez-Lanetty M, Loh W, Carter D, Hoegh-Guldberg O (2001) Latitudinal variability in symbiont specificity within the widespread scleractinian coral Plesiastrea versipora. Mar Biol 138:1175–1181
Rodriguez-Lanetty M, Krupp DA, Weis VM (2004) Distinct ITS2 types of Symbiodinium in clade C correlate with cnidarian/dinoflagellate specificity during onset of symbiosis. Mar Ecol Prog Ser 275:97–102
Rowan R, Knowlton N (1995) Intraspecific diversity and ecological zonation in coral-algal symbiosis. Proc Natl Acad Sci USA 92:2850–2853
Rowan R, Knowlton N, Baker A, Jara J (1997) Landscape ecology of algal symbionts creates variation in episodes of coral bleaching. Nature 388:265–269
Sampayo EM, Franceschinis L, Hoegh-Guldberg O, Dove S (2007) Niche partitioning of closely related symbiotic dinoflagellates. Mol Ecol 16:3721–3733
Santos SR, Taylor DJ, Coffroth MA (2001) Genetic comparisons of freshly isolated versus cultured symbiotic dinoflagellates: Implications for extrapolating to the intact symbiosis. J Phycol 37:900–912
Seutin G, White BN, Boag PT (1991) Preservation of avian blood and tissue samples for DNA analyses. Can J Zool 69:82–90
Stat M, Carter D, Hoegh-Guldberg O (2006) The evolutionary history of Symbiodinium and scleractinian hosts - symbiosis, diversity, and the effect of climate change. Perspect Plant Ecol Evol Syst 8:23–43
Thompson JD, Higgins DG, Gibson TJ (1994) ClustalW: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix code. Nucleic Acids Res 22:4673–80
Thornhill DJ, Fitt WK, Schmidt GW (2006a) Highly stable symbiosis among western Atlantic brooding corals. Coral Reefs 25:515–519
Thornhill DJ, LaJeunesse TC, Kemp DW, Fitt WK, Schmidt GW (2006b) Multi-year, seasonal genotypic surveys of coral-algal symbioses reveal prevalent stability of post-bleaching reversion. Mar Biol 148:711–722
Thornhill DJ, LaJeunesse TC, Santos SR (2007) Measuring rDNA diversity on eukaryotic microbial systems: how intragenomic variation, pseudogenes, and PCR artifacts confound biodiversity estimates. Mol Ecol 16:5326–5340
Toller WW, Rowan R, Knowlton N (2001) Zooxanthellae of the Montastraea annularis species complex: patterns of distribution of four taxa of Symbiodinium on different reefs and across depths. Biol Bull 201:348–359
Trench RK (1987) Dinoflagellates in non-parasitic symbioses. In: Taylor F (ed) The biology of dinoflagellates. Blackwell, Oxford, pp 530–570
van Oppen MJH (2004) Mode of zooxanthella transmission does not affect zooxanthella diversity in acroporid corals. Mar Biol 144:1–7
van Oppen MJH, Palstra FP, Piquet AM-T, Miller DJ (2001) Patterns of coral-dinoflagellate associations in Acropora: significance of local availability and physiology of Symbiodinium strains and host-symbiont selectivity. Proc R Soc Lond, B 268:1759–1767
van Oppen MJH, Mieog JC, Sanchez CA, Fabricius KE (2005) Diversity of algal endosymbionts (zooxanthellae) in octocorals: the roles of geography and host relationships. Mol Ecol 14:2403–2417
Weis VM, Reynolds WS, deBoer MD, Krupp DA (2001) Host-symbiont specificity during onset of symbiosis between the dinoflagellates Symbiodinium spp. and planula larvae of the scleractinian coral Fungia scutaria. Coral Reefs 20:301–308
Zardoya R, Costas E, Lopez-Rodas V, Garrido-Pertierra A, Bautista JM (1995) Revised dinoflagellate phylogeny inferred from molecular analysis of large-subunit ribosomal RNA gene sequences. J Mol Evol 51:26–40
Acknowledgments
The authors would like to thank T. Bui, K. Low, X. Pochon, T. LaJeunesse, W. Fitt, G. Schmidt, R. Gates, the staff at One Tree and Heron Island, and the reviewers who helped to improve this manuscript. This research was supported by an Australian Postgraduate Award to M. Stat and funded by an Australian Research Council grant (A10009205) to O. Hoegh-Guldberg and D. Carter, an Australian Biological Resources Study grant 204-53 to D. Carter, and by the GEF-World Bank Coral Reef Targeted Research Project ( www.gefcoral.org).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Biology Editor Dr. Michael Lesser
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License ( https://creativecommons.org/licenses/by-nc/2.0 ), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Stat, M., Loh, W.K.W., Hoegh-Guldberg, O. et al. Symbiont acquisition strategy drives host–symbiont associations in the southern Great Barrier Reef. Coral Reefs 27, 763–772 (2008). https://doi.org/10.1007/s00338-008-0412-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00338-008-0412-5