
Abstract
Genetic variation and gene flow in the scleractinian coral Goniastrea aspera (Verrill), found around the north–south Okinawa Islands, were studied using allozyme and starch gel electrophoresis. The relative contribution of sexual and asexual reproduction to recruitment was determined. Analysis of multilocus genotypes of samples, collected at least 3 m apart, identified a high number of unique genotypes (NG) relative to the number of individuals sampled (N) (mean NG: N=0.97±0.03 [SD]), and also highly observed genotypic diversity (GO) relative to expected genotypic diversity under Hardy–Weinberg equilibrium (GE) (mean GO: GE=0.95±0.06 [SD]). These results suggest that the collected G. aspera propagated predominantly by sexual reproduction in Okinawan populations. The UPGMA grouping of five populations in the Okinawa, based on Nei’s unbiased genetic distance, showed two clusters that were south and north Okinawa populations. AMOVA analyses that incorporated the data of populations from the Kerama and the Ishigaki Islands detected significant FSC values among the populations within groups in analyses distinguishing north–south Okinawa, Okinawa–Kerama, and Okinawa–Kerama–Ishigaki. Significant FCT values were neither detected between north–south Okinawa nor between the Okinawa–Kerama groups. The “local-recruits” hypothesis, which assumes that a substantial proportion of recruits is produced locally, appears to be applicable to coral populations in the Ryukyu Archipelago. On the other hand, detection of nonsignificant FCT values indicated the presence of genetic connectivity between north–south Okinawa and Kerama–Okinawa groups.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Coral bleaching is a global phenomenon that is probably linked to global climate change and increasing ocean temperature (Glynn 1991; Brown 1997; Hoegh-Guldberg 1999). The 1998 coral-bleaching event was the most extensive and severe that has yet been recorded (e.g., Hoegh-Guldberg 1999; Goreau et al. 2000), and coral populations in the Okinawa Islands, particularly populations of branching species, declined severely during the 1998 mass-bleaching event (Loya et al. 2001). The Kerama and the Ishigaki Islands, 30 and 400 km from the Okinawa Islands, respectively, maintained healthy coral populations on many reefs even after mass coral bleaching (Taniguchi and Iwao 2000; Ministry of the Environment of Japan 2002).
The data of the surface current, larval ecology, and population genetics strongly suggest that coral populations at the Kerama Islands will be the source of larval supply for the recovery of coral populations around the Okinawa Islands. Nadaoka et al. (2002a) studied the current flows around the Kerama and the Okinawa Islands using GPS buoys and high-frequency (HF) radar. They also cultured larvae of Acropora tenuis and Acropora nasuta under laboratory conditions to know the period before planula attachment after spawning. Nadaoka et al. (2002a) showed that the sea surface current flowed from the Kerama Islands to the Okinawa Islands during the spawning period of Acropora spp. (full moon in June), and the time required for the buoys to reach the west coast of Okinawa from the Kerama Islands corresponded to the length of the period between spawning and the peak of planula attachment (4–5 days). Population genetic study by Nishikawa et al. (2003) showed that gene flow between the Kerama and the west coast of Okinawa was higher than between the Yaeyama and the west coast of Okinawa for A. tenuis and Stylophora pistillata. We refer to this as the “source-sink” hypothesis in this paper. The study by Nishikawa et al. (2003) used the bleaching-susceptible species (“losers” in Loya et al. 2001). Thus, their analysis may include the effect of coral bleaching on genetic structure of coral populations. In the present study, we examine the coral Goniastrea aspera whose mortality due to bleaching was low (one of “winners” in Loya et al. 2001).
There is little information on the genetic connectivity of coral populations around the Okinawa Islands. Adjeroud and Tsuchiya (1999) studied genetic variation in the coral Pocillopora damicornis between population on the west and east coasts but only at the northern part of the Okinawa Islands before the 1998 mass coral bleaching. Nishikawa et al. (2003) and Nishikawa and Sakai (2003) studied A. tenuis, S. pistillata, and G. aspera but only on the west coast of Okinawa Islands after the bleaching. In the present study, we took samples from populations of G. aspera all around the Okinawa Islands to reveal the connectivity of populations around the Okinawa Islands.
Ayre and Hughes (2000) studied the population genetics of five brooding and four spawning coral species across 1,800 km of the Great Barrier Reef (GBR) using allozyme electrophoresis. They found significant genetic variations among local populations within geographic groups (a few km scales) for all nine species (four spawning and five brooding species), but not among the geographic groups (1,800 km scales) for two spawning and three brooding species out of nine. They concluded that a substantial proportion of recruits in a given reef were produced in that reef (“local-recruits” hypothesis). However, they also argued that gene flow is sufficient to prevent the accumulation of fixed genetic differences among the groups. The Okinawa Island is elongated in the north–south direction and the distance between the northern and southern ends is about 100 km. Thus, populations of some coral species may also be genetically differentiated.
Some scleractinian corals may display asexual reproduction, through fragmentation (Ayre and Willis 1988), and the planula-brooding of asexual planula (Stoddart 1983). The degree of asexual reproduction that can lead to gene frequencies that are skewed from those expected under random mating has effects on the genetic structure of populations. Therefore, knowledge of the relative contribution of sexual and asexual reproduction to G. aspera recruitment in the regions is important for inferring gene flow from gene frequencies.
Goniastrea aspera (Verrill) is widely distributed in Indo–Pacific reefs. Abe (1937) and Motoda (1939) reported that G. aspera in Palau released brooded planulae, while Babcock (1984) reported that G. aspera in the GBR was a hermaphroditic spawner. In the Ryukyu Archipelago, an individual colony of G. aspera brooded planulae after spawning (Sakai 1997). Nishikawa and Sakai (2003) studied the gene flow of G. aspera among the islands of the Ryukyu Archipelago, and compared the degree of gene flow with two other species (A. tenuis and S. pistillata) that was already published in the same geographic range (Nishikawa et al. 2003). They showed that G. aspera had greater gene flow than the brooding species (S. pistillata) but smaller than the spawning ones (A. tenuis). They indicated that gene flow, attributed to spawned planulae, prevents the accumulation of fixed genetic differences by philopatric dispersal by brooded planulae. In addition, relatively low survival rates of spawned planulae may bring about limited gene flow compared to spawning species of A. tenuis.
This study determined the degree of gene flow of G. aspera around the Okinawa Islands. We incorporated the data from a previous publication that studied genetic differentiation among three populations of G. aspera in the west coast of the Okinawa Islands (Nishikawa and Sakai 2003), in which the same allozyme system as in the present study was used in the same species. Further, we discuss the relevance of the “source-sink” and “local-recruits” hypotheses by incorporating the data from the Kerama and the Ishigaki populations from the previous study (Nishikawa and Sakai 2003).
Materials and methods
Collection of specimens and electrophoresis
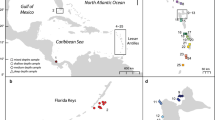
Five sampling sites were established in the Okinawa Islands: two new sites and three sites used previously (Nishikawa and Sakai 2003) (Fig. 1). Coral fragments in each population were taken from 33 to 50 colonies found along a swimming trajectory in 2002. We collected samples the same year as Nishikawa and Sakai (2003). Colonies from which the samples had been taken were separated by at least 3 m to avoid repeated collection of the same clone. The collected fragments were kept alive in a small plastic tank filled with seawater and brought to the laboratory of the Tropical Biosphere Research Center (TBRC) at the University of the Ryukyus. The fragments were reared in an outdoor holding tank (2.0×1.0×3.5 m) supplied with running seawater at TBRC. Coral fragments were frozen and stored at −30°C for at least 2 h before allozyme electrophoresis.

Map of the Ryukyu Archipelago and Okinawa Islands showing five collection sites indicated (*). (OHD Ohdo, SES Sesoko, HED Hedo, GES Gesashi, KIN Kin). Arrows indicate Kuroshio Current flows
Frozen fragments were ground in a solution of 0.1% B mercaptoethanol, 10% sucrose, 0.1% bromphenol blue, and 0.25 mg/ml NADP; the resulting homogenate was centrifuged at 3,000 rpm for 3 min. The supernatants were absorbed onto Whatman No. 3 chromatography paper wicks (3×10 mm) and then loaded onto horizontal starch gels (12%). We initially screened for genetic variation in ten enzyme systems, following the methods of Selander et al. (1971) and Hillis et al. (1996), using horizontal starch gels (12%: at 100–180 V and 36–48 mA for 6–9 h). Based on these results, five enzyme systems were used: malate dehydrogenase (Mdh; E.C.#1.1.1.37), phosphoglucomutase (Pgm; E.C.#5.4.2.2), hexokinase (Hk; E.C.#2.7.1.1), and peptidase using leucyl-tyrosine as a substrate (Lt; E.C.#3.4.11), and leucyl-glycyl-glycine as a substrate (Lgg; E.C.#3.4.11). Tris EDTA citrate buffer (pH 7.5) was used for Mdh, Pgm, and Hk, and Tris citrate buffer (pH 8.0) was used for Lgg and Lt.
Statistical analysis
Genetic variability, allele frequencies, and genetic distance were analyzed using TFPGA (Miller 1997). Genetic variability, which included the mean number of alleles per locus, the percentage of polymorphic loci (95% criterion), the observed heterozygosity, and the expected heterozygosity under Hardy–Weinberg equilibrium (HWE), were calculated for each population. The magnitude and direction of departures from HWE at each locus were also assessed for each population. These departures were expressed as Wright’s fixation index (F), where positive and negative values represented deficits and excesses of heterozygotes, respectively. Five tests were conducted and Bonferroni’s adjustment was used to test the significance of departures from HWE (significant: P<0.01). The dendrogram based on unbiased genetic distance (Nei 1978) was conducted using the UPGMA.
Two measures were used to assess the possible effects of asexually derived recruits on the genotypic diversity of the collections. First, each colony was assigned a multilocus (clonal) genotype. The number of multilocus genotypes detected, NG, was an estimate of the minimum number of clones present within a population; the ratio NG: N, where N is the number of individual colonies collected, provided a simple index of the effect of asexual reproduction on genotypic diversity. Second, the ratio of observed multilocus genotypic diversity (GO) to that expected under conditions of sexual reproduction (GE) was calculated following Stoddart and Taylor (1988). Departure of GO: GE from unity was used as an index of the combined effect of departures from singlelocus HWE and multilocus linkage disequilibrium. A genetically variable population with high levels of asexual recruitment would have a low ratio of observed to expected genotypic diversity.
We calculated a hierarchical analysis of molecular variance (AMOVA). AMOVA enables investigation of population structure by testing F-statistics at different levels of spatial hierarchy; those are among local populations within the group of sampling sites (FSC) and among the groups of sampling sites (FCT). Tests of statistical significance of the AMOVA were based on nonparametric permutation procedures using 16,000 permutations (Excoffier et al. 1992). Gene flow was estimated using Wright’s (1969) Island model (Nem=[(1/θ)−1]/4, where Ne is the effective population size and m is the proportion of migrants per generation).
Results
The mean number of alleles per locus in each subpopulation ranged from 3.2 to 4.0, and the percentage of polymorphic loci (P95) ranged from 80% to 100% (Table 1). The observed mean heterozygosity in each population was lower than the expected heterozygosity assuming HWE (paired t-test, P<0.001). Only one fixed locus was found, the Lt-2 locus in the GES (Gesashi) population (Table 2). Significant heterozygosity deficits from HWE were found in three of five loci; Mdh, Lt-1, and Lt-2 (Table 3). In particular, Lt-1 and Lt-2 loci often exhibited significant deviations from HWE. Heterozygote deficits occurred in 21 out of 24 cases, and significant deviations from HWE were detected in seven cases.
The ratio of the number of observed genotypes (NG) to the number of individuals sampled (N) ranged from 0.92 to 1.00, and the ratio of observed multilocus genotypic diversity (GO) to that expected under conditions of sexual reproduction (GE) among collection sites ranged from 0.87 to 1.01 (Table 4). Both ratios of NG: N and GO: GE were high (0.97±0.03 and 0.95±0.06, respectively; mean ± SD), and there were no significant differences between GO and GE using unpaired t-test (following Stoddart and Taylor 1988). Both these measures (NG: N and GO: GE) indicate a high degree of sexual reproduction within the five populations examined.
The unweighted pairwise group method using arithmetic averages (UPGMA), based on Nei’s (1978) unbiased genetic distance, identified two clusters: one containing the OHD and KIN populations (south Okinawa, hereafter referred to as S-Okinawa) and the other the SES, HED, and GES populations (north Okinawa, hereafter referred to as N-Okinawa) (Fig. 2).

The UPGMA dendrogram showing relationship among populations in the Okinawa based on Nei’s (1978) unbiased genetic distance D
The AMOVA analysis assessed variation 2 hierarchies (among group and among populations within group) in three analyses using different geographic groups, S–N Okinawa, Okinawa–Kerama, and Okinawa–Kerama–Ishigaki (Table 5). The percent of molecular variation was greater among populations within groups than among groups. F-statistics among populations within group (FSC) were significant in all three geographic groupings while those among groups (FCT) were not detected in S–N Okinawa and Okinawa–Kerama. FSC values were greater than FCT in all three geographic groups. The number of migrants per generation at equilibrium (Nem) was 3.8 to 6.2 and 7.1 to 12.9 among populations within group and among group, respectively.
Discussion
The genotypic diversity of the five populations of G. aspera from the Okinawa Islands indicates that the sampled colonies, all of which were separated by more than 3 m, were produced primarily by sexual reproduction. Populations dominated by asexual reproduction (i.e., fragmentation or larvae of asexual origin) generally have low ratios for both the number of multilocus genotypes to the number of individuals (NG: N) and the observed genotypic diversity to the expected genotypic diversity (GO: GE) (Stoddart 1984; Ayre and Willis 1988). Stoddart (1984, Pocillopora damicornis) and Ayre and Willis (1988, Pavona cactus) found mean values for NG: N and GO: GE of 0.40 and 0.27, and 0.35 and 0.35, respectively, and both studies concluded that these low ratios are typical of coral populations dominated by asexual recruitment. By contrast, Dai et al. (2000, Mycedium elephantotus) and Nishikawa and Sakai (2003, G. aspera) found mean values for NG: N and GO: GE of 0.98 and 0.94, and 0.90 and 0.84, respectively, suggesting the predominance of sexual reproduction in these coral populations. Here, we found high values of NG: N and GO: GE (0.97± 0.03 and 0.95±0.06) in the populations of G. aspera. Together with a lack of histological evidence for asexually produced larvae in G. aspera (Sakai 1997), these results indicate that asexual recruitment has contributed very little to the maintenance of G. aspera populations around the Okinawa Islands.
It is unlikely that frequent coral recruitments in G. aspera occur around the Okinawa Islands. UPGMA based on Nei’s (1978) genetic distance produced two clusters: one consisting of populations from the N-Okinawa (SES, HED, and GES) and the other consisting of populations from the S-Okinawa (OHD and KIN). This suggests that the coral populations are subdivided between the north and the south populations around the Okinawa Islands. On the other hand, the value of FCT among the N-S Okinawa groups was not significant (0.026 and 9.4 in FCT and Ne m, respectively). Significance was detected in FSC values among populations within groups (0.039 and 6.2). These results support the local-recruit hypothesis (see below) but also suggest the presence of genetic connectivity between the N–S Okinawa groups around the Okinawa Islands.
The significant FSC values (among populations within groups) and nonsignificant FCT values (among group) were also found in the relatively small geographic scale of part of Ryukyu Archipelago (30–100 km). The significant FSC values (among population within group) were detected in all three geographic groups (0.039 and 6.2, 0.061 and 3.8, and 0.057 and 4.1 in FSC and Ne m in N–S Okinawa, Okinawa–Kerama, and Okinawa–Kerama–Ishigaki group, respectively) but those of FCT (among group) were not significant in the N–S Okinawa and the Okinawa–Kerama groups. Additionally, the percentage of molecular variation was greater among populations within the groups than among the groups. These results are similar to those by Ayre and Hughes (2000); they found significant genetic variation among local populations within geographic groups for all nine species but not among the groups for five out of nine species. The “local-recruits” hypothesis, which assumes that a substantial proportion of recruits is produced locally, is applicable to coral populations in the Ryukyu Archipelago. On the other hand, Ayre and Hughes (2000) also concluded that gene flow is sufficient to prevent the accumulation of fixed genetic differences among vast geographic scales along the GBR. We suggest that the nonsignificant value in the small scale of the present study is probably due to mostly local dispersal coupled with rare dispersal between populations in different groups, so that we might detect significance of F-statistics among populations within groups but not among groups.
The present study also supports the “source-sink” hypothesis, which assumes that coral populations in the Kerama Islands have been the source of larval supply for the recovery of coral populations around the Okinawa Islands since the 1998 mass coral bleaching. The nonsignificant value of FCT for the Kerama–Okinawa group in the present study suggests some gene flow between the Kerama and the Okinawa Islands. In addition, Nadaoka et al. (2002a, b) demonstrated that a slick of coral eggs and larvae produced in the Kerama Islands was transported to the west coast of Okinawa Islands by the surface currents, suggesting the dispersal of coral larvae produced at the Kerama Islands to the Okinawa Islands. Our data suggest that both “local-recruits” and “source-sink” hypotheses are relevant to G. aspera that was affected little by the1998 mass coral bleaching (Loya et al. 2001). For coral species that declined severely due to the bleaching such as Acropora spp. (Loya et al. 2001), adult populations that produce larvae for “local recruits” are not present around the Okinawa Islands, and “source-sink” hypothesis is especially relevant.
References
Abe N (1937) Postlarval development of the coral Fungia actiniformis var palawensis Döderlein. Palao Trop Biol Stat Stud 1:73–93
Adjeroud M, Tsuchiya M (1999) Genetic variation and clonal structure in the scleractinian coral Pocillopora damicornis in the Ryukyu Archipelago, southern Japan. Mar Biol 134:753–760
Ayre DJ, Hughes TP (2000) Genotypic diversity and gene flow in brooding and spawning corals along the Great Barrier Reef, Australia. Evolution 54(5):1590–1605
Ayre DJ, Willis BL (1988) Population structure in the coral Pavona cactus: clonal genotypes show little phenotypic plasticity. Mar Biol 99:495–505
Babcock RC (1984) Reproduction and distribution of two species of Goniastrea (Scleractinia) from the Great Barrier Reef province. Coral Reefs 2:187–204
Brown BE (1997) Coral bleaching: causes and consequences. Coral reefs 16:129–138
Dai CF, Fan TY, Yu JK (2000) Reproductive isolation and genetic differentiation of a scleractinian coral Mycedium elephantotus. Mar Ecol Prog Ser 201:179–187
Exocoffier L, Smouse P, Quattro J (1992) Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochondrial DNA restriction data. Genetics 131:479–491
Glynn PW (1991) Coral reef bleaching in the 1980s and possible connections with global warning. Trends Ecol Evol 6:175–179
Goreau TJ, McClanahan TR, Hayes RL, Strong A (2000) Conservation of coral reefs after the 1998 global bleaching event. Conserv Biol 14:5–15
Hillis DM, Moritz C, Barbara KM (1996) Molecular Systematics. Sinauer Associates, incorporated
Hoegh-Guldberg O (1999) Coral bleaching, climate change, and the future of the world’s coral reefs. J Mar Fresh Res 50:839–866
Loya Y, Sakai K, Yamazato K, Nakano Y, Sambali H, Van Woesik R (2001) Coral bleaching: the winners and losers. Ecol Lett 4:122–131
Miller MP (1997) Tools for population genetic analysis (TFPGA) 1.3: a Windows program for the analysis of allozyme and molecular population genetic data. Computer software distributed by the author
Ministry of the Environment of Japan (2002) Report of the international Coral Reef Research and Monitoring Center (in Japanese). International Coral Reef Research and Monitoring Center, Ishigaki, pp 13–34
Motoda S (1939) Observation of period of emergence of planulae of Goniastrea aspera Verrill (in Japanese, title translated by KS). Kagaku Nanyo 1:113–115
Nadaoka K, Harii S, Mitsui J, Tamura H, Hanada G, Paringit E, Nihei Y, Fujii S, Sato K, Matsuoka T, Kakuma S, Ikema T, Iwao K, and Takahashi T (2002a) Larval tracking using small drifters and larval settlement experiments to examine long-distance larval transport of corals (in Japanese). Proc Coastal Eng JSCE 49:366–370
Nadaoka K, Harii S, Ikema T, Paringit E, Mitsui J, Tamura H, Iwao K, Kakuma S (2002b) Coral Spawning and Slick Observations in Kerama Island, Okinawa (in Japanese). Proc Coastal Eng JSCE 49:1176–1180
Nei M (1978) Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 89:583–190
Nishikawa A, Sakai K (2003) Genetic variation and gene flow of broadcast spawning and planula brooding coral, Goniastrea aspera (Scleractinia) in the Ryukyu Archipelago, southern Japan. Zool Sci 20:1031–1038
Nishikawa A, Katoh M, Sakai K (2003) Larval settlement rates and gene flow of broadcast-spawning (Acropora tenuis) and planula-brooding (Stylophora pistillata) corals (Scleractinia). Mar Ecol Prog Ser 256:87–97
Sakai K (1997) Gametogenesis, spawning, and planula brooding by the reef coral Goniastrea aspera (Scleractinia) in Okinawa, Japan. Mar Ecol Prog Ser 151:67–72
Selander RK, Smith MH, Yank SY, Johnston WE, Gentry JR (1971) Biochemical polymorphism and systematics in the genus Peromyscus I Variation in the old-field mouse (Peromyscus polionotus). Stud Genet VI University of Texas Pub 7103:49–90
Stoddart JA (1983) Asexual production of planulae in the coral Pocillopora damicornis. Mar Biol 76:279–284
Stoddart JA (1984) Genetic structure within populations of the coral Pocillopora damicornis. Mar Biol 81:19–30
Stoddart JA, Taylor JF (1988) Genotypic diversity: estimation and prediction in samples. Genetics 118:705–711
Taniguchi H, Iwao K (2000) Change of coral number and density in Aka Island (In Japanese). Rep Akajima Mar Sci Lab 11:22–23
Wright S (1969) The evolution and genetics of populations. University of Chicago press, Chicago
Acknowledgements
We are indebted to the staff of Sesoko Station, Tropical Biosphere Research Center, University of the Ryukyus. Thanks are due to many students in TBRC for their encouragement, help, and advice. We are very grateful for two anonymous reviewers who helped improve the manuscript. This study was supported by Grant-in-Aid of Scientific Research (B) (No. 14205071) to K. Nadaoka and (C) (No. 12640619) to K. Sakai from the Ministry of Education, Culture, Sports, Science and Technology, Japan.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Biological Editor H.R. Lasker
Rights and permissions
About this article
Cite this article
Nishikawa, A., Sakai, K. Genetic connectivity of the scleractinian coral Goniastrea aspera around the Okinawa Islands. Coral Reefs 24, 318–323 (2005). https://doi.org/10.1007/s00338-005-0484-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00338-005-0484-4