
Abstract
Archaeological wood charcoal analysis or anthracology has been applied for the first time in New Caledonia as part of an interdisciplinary research program examining Kanak pre-colonial landscape management in a valley on the northeast coast of the subtropical Pacific island. In contrast to previous hypotheses, this study demonstrated that when the Kanak traditional cultural complex emerged around ad 1000, following the initial 2,000 years of human presence on the island, the vegetation cover showed few signs of deforestation and the tropical rainforests were still prominent. The vegetation surrounding Kanak settlement sites evolved during the first half of the 2nd millennium ad towards a more open but more complex composition that included useful taxa. This was interpreted as showing a form of forest management and possible arboriculture (the cultivation and management of trees). However, in sharp contrast to its Melanesian neighbours, very little is known about arboricultural practices in New Caledonia. Through the interpretation of data from two sites in particular, a discussion of the two-step analytical process is used to argue for the existence of arboricultural practices associated with these sites: (1) using anthracological data to reconstruct the vegetational landscape, together with ethnobotanical, ethnohistorical and archaeological data; (2) allowing for the recognition of specific practices of forest domestication, based on the manipulation of plants and of spatial patterns of forests. At a time when archaeology is engaged in a process of post-colonial re-evaluation of its schemes of interpretation, it seems timely for archaeobotany to try integrating more indigenous systems of representation into its analyses. The approach presented here is an effort in this direction, trying to make sense of new data by reading them through a local lens.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
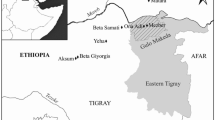
New Caledonia is a subtropical Pacific archipelago located at the southern tip of the so-called region of Melanesia, first peopled 3,000 years ago during the Lapita cultural expansion (Fig. 1; Sand 2010). Within this archipelago, the study of Kanak traditional societies has long been the field of ethnology, while analysis of traditional plant management practices has mainly been the focus of cultural geographers (Dotte 2010). Multidisciplinary and diachronic approaches on these questions have remained rare, with archaeology originally concentrating on issues of first settlement (Sand et al. 2008) and with sparse palaeoenvironmental data available (Sémah 1998; Stevenson 2004).

Location of New Caledonia and the Tiwaka valley, adapted from Dotte-Sarout (2010)
To start addressing these gaps in our knowledge, a multidisciplinary research project combining anthracology (wood charcoal analysis), settlement patterns in archaeology and ethnohistorical as well as ethnological analyses was conducted in the Tiwaka valley, northeastern Grande Terre (Fig. 1; Dotte-Sarout 2010; Dotte-Sarout et al. 2013a). The main goal of the project was to understand past changes in forest composition and uses by humans of vegetation resources, in order to define the dynamics of landscape transformation during the Kanak pre-colonial period, from around ad 1000 to the late 19th century (Sand et al. 2008).
Based on the subsistence model classically applied to New Caledonia and the Pacific islands in general (e.g. Haberle et al. 2010; Kirch 2000, 2004; Kirch and Hunt 1997; McWethy et al. 2010; Sand 2002), the initial use of an extensive slash-and-burn horticultural system with the arrival of people 3,000 years ago would have left strong signatures of deforestation on the vegetation. The subsequent development of cultivation techniques using irrigated terraces and drained mounds during the Kanak pre-colonial period is usually seen as a more sustainable way to exploit natural resources and to reduce the human impact on the environment (Sand 2002; Sand et al. 2008). Surprisingly, the results of the present study produced a more complex history in the Tiwaka valley, beginning in the 2nd millennium ad with Kanak land occupation patterns developing in a well-forested landscape showing few signs of deforestation. Rapid forest perturbations and savanna extension appear after the 17th century ad, when the human presence in the valley seems to have intensified with the development of settlement and horticultural sites (Dotte-Sarout et al. 2013a). These results stand in sharp contrast to the expectations produced by the classic model of itinerant slash-and-burn horticulture usually associated with the first millennia of human presence in Pacific archaeology narratives (e.g. Haberle et al. 2010; Kirch 2000, 2004; Kirch and Hunt 1997; McWethy et al. 2010; Sand 2002). However, and rather paradoxically, the results make sense in relation to the large body of ethnological data on traditional practices of arboriculture and on the existence of complex practices of forest “domestication” or management throughout Melanesia and southeast Asia (Barrau 1962; Blench 2005; Kennedy 2012; Latinis 2000; Walter and Sam 1999; Yen 1996), but only rarely referring to New Caledonia. Here, two case studies are presented from the research undertaken in the Tiwaka valley to discuss the two-step analytical process that was used to move from anthracological data to interpretation within local sociocultural frameworks, eventually leading to an argument for the existence of specific practices of forest domestication in New Caledonia.
Materials and methods
Methods
This project represents the first application of anthracology in New Caledonia, and in the Melanesian region generally (Dotte-Sarout et al. 2015). The archipelago is renowned for its highly diverse and peculiar flora, constituting, alone, one of the world’s most floristically diverse regions with more than 3,200 indigenous species of vascular plants, of which 77 % are endemic (Jaffré et al. 2004). The pioneering work of Rita Scheel-Ybert in a similarly diverse and complex tropical environment in Brazil (2000, 2001, 2002) has provided a set of essential references, especially with regard to diversity indices and size of assemblages. A local reference collection of 142 woody taxa from New Caledonia was first established by the author and a wood anatomy database developed (Dotte-Sarout 2010). To strategically prioritize certain taxa with regard to the extreme richness of the New Caledonian flora, botanical surveys were conducted in association with a review of the ethnobotanical and botanical literature (Dotte 2010). In accordance with the main ecological characteristics of the Tiwaka valley, and within the archaeological focus of the study, the collection was orientated towards species associated with rainforest (65 % of the taxa) or with human activities such as cultivation, symbolic use or as ruderals, with an over-representation of non-endemic species (close to 50 % of the collected taxa).
Three pre-colonial Kanak settlement sites located in different parts of the valley were chosen for excavation. Here, we focus on the two providing the richest anthracological records, Pwadaunu, a site located on a mountain slope at one of the higher elevations in the valley, and Komijièn, a site located on the coastal alluvial plain, close to the mouth of the river Tiwaka. Abandoned traditional Kanak habitation sites are visible in the landscape as clusters of round mounds on top of which were once erected the houses of individual nuclear families. One settlement mound was chosen at each of the sites for excavation, according to various criteria (Dotte-Sarout 2010). Test pits of 1 × 1 m to 2 × 1 m were excavated, so as to expose the three usual layers of a traditional Kanak house mound.
-
1.
the pre-construction layer, expected to yield an anthracological assemblage representative of vegetation clearance before construction;
-
2.
the mound construction layer, expected to hold an anthracological deposit related to the very first occupation of the site, as new house mounds were progressively constructed and the site expanded as the resident population increased (Dotte-Sarout et al. 2013b; Sand et al. 2008);
-
3.
the occupation(s) layer(s), expected to produce an anthracological assemblage representing the later phase(s) of occupation of the site, in particular the occupation(s) of the related house.
All excavated sediments were collected by stratigraphic units and features and wet-sieved in a nearby river with 2 and 4 mm meshes. Because flotation proved to be inefficient due to the clayey nature of the sediment, all charcoal fragments were handpicked directly from the sieves, dried and bagged in the field. The two sites described in the present study did not yield any concentration features within the excavation test pits, so ‘assemblage’ hereafter relates only to samples created by the collection of dispersed charcoal from occupation, construction and pre-construction layers (Dotte-Sarout 2010, Dotte-Sarout et al. 2013a). Preparation, observation and identification of charcoal fragments followed standard anthracological techniques (Chabal et al. 1999), with reference to the local wood anatomy database (ESM). Based on an ideal size for charcoal assemblages from tropical environments suggested by Scheel-Ybert (2002), archaeological assemblages consisted of 300–400 identifiable fragments depending on the deposit charcoal richness, while pedo-anthracological assemblages from the pre-construction layers contained around 100 fragments each, as these were generally in poorer states of preservation. Exact numbers for the Pwadaunu and Komijièn sites are presented in Table 1. Saturation curves (taxa/fragments) and Gini-Lorenz indices (taxa rank/taxonomic diversity) were used to test the ecological diversity of the assemblages and guarantee their representativity (Chabal et al. 1999; Dotte-Sarout et al. 2015). Most of the Gini-Lorenz indices are between 25:75 and 29:71, in accordance with the ideal reference of 25:75 and lower limit of 30:70, as calculated for diverse tropical environments by Scheel-Ybert (2002) (Table 1). There are two low indices for Komijièn (32:68, 36:64) and these reflect assemblages where extreme diversity is likely to have resulted from two factors: (1) mixed origins of original wood samples from a composite environment of forests, mangroves and riverine as well as coastal vegetation; (2) depositional contexts favouring high taxonomic diversity (stratigraphy in Dotte-Sarout 2010, Dotte-Sarout et al. 2013a).
Overall, two thirds of the fragments analysed were identified, more than 50 % to genus level, representing 136 different taxa, with high taxonomic diversity in the assemblages (Table 1). Unidentified taxa comprise 70 different types, represented in proportions of less than 1 % of the assemblages, except for one type that reached 9.4 % in one assemblage from the third site, which is not considered here. Twelve taxa were found in half or more than half of all the assemblages analysed during the project, eight in total, and showed the highest occurrence in proportions of fragments, ranging from 2 to 13 % per assemblage on average (Table 2 and ESM). Among these, nine are typical rainforest taxa, and one is a savanna woody taxon, Melaleuca quinquenervia, which often spreads following fire. However, the most frequently occurring taxon in all the assemblages is from the Casuarinaceae family. The latter comprises two genera that demonstrate close anatomical similarity (Fig. 2). Only one species, Casuarina equisetifolia can be clearly differentiated, by the length and width of its rays (Détienne and Jacquet 1999; Dotte-Sarout 2010). It is associated with halophytic vegetation and is found in low frequency at Komijièn. The majority of the Casuarinaceae found in both assemblages could be either: (1) C. collina, a very wide-ranging species that inhabits wetlands, open, dry or secondary vegetation and which is planted close to houses; or (2) Gymnostoma nodiflorum, a species characteristic of deep valley riverside vegetation, and one of the main woody taxa in a specific type of rainforest, characterised by an open and light-demanding vegetation, called “photoxerophilous rainforest type” (Jaffré et al. 1994, 2004; Virot 1951).

Anatomy details of Casuarinaceae wood charcoals found in high frequencies in the study
Archaeological and theoretical background
On the Grande Terre, the Kanak territorial settlement system appears to have been organized as inter-connected spatial, social and economic units that had a certain degree of mobility within a fixed territory, known as “country” in the Kanak languages (Dotte et al. 2013b; Sand et al. 2008; Saussol 1990). This territory, ideally corresponding to an alluvial valley, was linked, according to anthropological studies, to a chiefdom and to an extended kinship group associated with the “house” society type (Bensa and Rivierre 1982, pp. 32–33).
Most archaeological occupation sites identified on the Grande Terre are related to artificial earth features for horticultural use of the land, either drained or irrigated. This division of the landscape between dry and wet areas corresponds to a spatial model widely applied throughout Oceania (Barrau 1962, 1965). For dry crop cultivation mainly of numerous local varieties of Dioscorea alata and D. esculenta (yams), these features consist of parallel elongated raised mounds separated by drainage trenches, arranged according to the topography (Barrau 1956). For growing wet crops mainly of numerous varieties of Colocasia esculenta (taro) associated with Saccharum officinarum (sugar cane) and Musa spp. (banana), the features consist of irrigated terraces that were built around the slopes of mountains and hills. Ethnohistorical accounts also report that extensive terraces and other types of irrigation structures existed in the alluvial plains or on smaller islands of the archipelago (Barrau 1956; Sand 2012).
According to current archaeological datasets, it has been proposed that the wet and dry types of horticultural features could have had different histories (Dotte-Sarout et al. 2013a). Indeed, the dry horticulture features could be seen as stemming from pre-Lapita traditions which had developed in Papua New Guinea from the early Holocene (Denham 2005), while irrigated fields appear to have emerged at the end of the 1st millennium ad on the Grande Terre and then developed on a particularly large scale there relative to other Pacific islands (Sand 2012). Until the present study, it was generally considered that these types of constructions represented a sustainable answer to the destructive effects of the progressive intensification of agricultural slash-and-burn practices before this. This interpretation was based mainly on the observation of some thick sedimentary layers and increased charcoal deposits directly pre-dating the development of the Kanak complex (Sand 2002; Sand et al. 2008), and on pollen analyses which showed an opening-up of the vegetation in the centuries following the arrival of people on New Caledonia in the 1st millennium bc (Sémah 1998; Stevenson 2004). Such data and explanation were easily integrated within the classic regional interpretative framework applied throughout the Pacific regarding the history of human deforestation on the islands (Kirch 2004).
In parallel, the work of cultural anthropologists has tended to emphasize the universal presence and multiple functions of plants, especially woody taxa, within Kanak societies (Bensa and Rivierre 1982; Guiart 1956; Leenhardt 1953; Saussol 1990). Although the importance of plants in the Kanak world is usually taken for granted, the actual presence of trees or plants around ancient occupation sites can be easily overlooked in archaeological studies that deal with spatial occupation patterns or in non-archaeological reconstructions of past environments.
To address these issues, and in order to help build a better theoretical framework for the anthracological analysis, a preliminary study was conducted to explore models of management of the vegetation within the territorial framework and the associated subsistence system of Kanak pre-colonial societies (Dotte 2010). Archaeological field surveys were done at the beginning of the research programme which were associated with botanical surveys and ethnobotanical observations. Comparison of the various types of field data with ethnological, archaeological, botanical or ethnobotanical literature resulted in a redefinition of the pre-colonial horticultural system. In particular, it was proposed that a specific territorial pattern existed which integrated trees and other woody plants with forested land. Three kinds of vegetation areas were characterised, based on taxonomic diversity and the degree of domestication, in the sense of human manipulation. Twenty species most often associated with pre-colonial Kanak habitation sites were also identified (Table 3).
The first area was characterized as the domestic space, one dominated by culture, in every sense of the word. This is where the occupation and horticultural structures or gardens are found, and where the pre-existing forest had been cleared to plant crops and symbolic or useful plants, according to a strict spatial organization (Bensa and Rivierre 1982; Guiart 1956; Leenhardt 1953; Saussol 1990). This domestic area comprises two distinctive spatial units that can mix. One is a strictly social and ritual space where only plants fulfilling a symbolic role are found, such as Araucaria columnaris, Cocos nucifera, Cordyline and Dendrocalamus. The other is the horticultural economic space consisting of small gardens adjacent to the settlement or more distant and extensive horticultural features where food crops are grown, including some trees such as Musa spp., Syzygium malaccense and Artocarpus altilis, and later on, post contact introductions such as Carica papaya (Table 3; Dotte 2010).
The regular occurrence of several crops or symbolic tree taxa on the edge of archaeological occupation sites seems to indicate the past frontier zone between the domestic area and the forest. This hybrid area, the second vegetation area identified as part of the territorial system, is characterized by the presence of rainforest taxa that also produce edible fruits and nuts, or woody plants with known ecological, ritual, useful or aesthetic functions. Several of these important attributes are often present in a single tree taxon, for example Hibiscus tiliaceus, Fagraea berteroana and Ficus spp. (Table 3; Dotte 2010).
Based on oral traditions and ethnographical studies, the forest is the domain of the “uncultivated”, rather than the “wild” in the Western sense of the term (Haudricourt 1964). This uncultivated land, the third vegetation area, is a space occupied by various elements that have left the “cultivated” world for a time only: ancestor spirits or abandoned settlements and horticultural structures in fallow. This forest is still part of the daily landscape though, crossed by the paths that connect related occupation sites and horticultural features, or places that are used for hunting. These trips and specialized visits facilitate the collection of fruits, nuts or other edible plant parts, as well as timber (Barrau 1962; Leenhardt 1953) and, we can hypothesize, firewood.
In neighbouring islands and archipelagos where detailed ethnobotanical studies on arboriculture have been undertaken, specific practices of resource management within the forest have been recorded. The location of the most useful trees is precisely known by local communities and transmitted within a social group. Specific trees are tended by pruning or protection during slash and burns, and useful taxa can be transplanted within or between determined forest areas to extend their range or distribution, or, in the case of important economic crops such as fruit trees, be planted along paths (Kennedy 2012; Latinis 2000; Walter and Sam 1999; Yen 1996). Several observations made in the field in New Caledonia tend to show that similar practices may have existed during pre-colonial times (Dotte 2010); in line with Haudricourt’s definition of the forest as the uncultivated, that is, not part of the social world, rather than the natural or wild space, the forested landscapes surrounding Kanak habitation sites could have been profoundly manipulated environments. However, until the present study, no precise archaeobotanical data could directly confirm or refute such a model.
Results
Pwadaunu
The occupation site of Pwadaunu is situated at ca. 500 m a.s.l. on Mwîdip mountain, which reaches 959 m a.s.l. A total of 88 different taxa were identified including 15 indeterminate types within the three assemblages studied in the anthracological analysis.
The oldest assemblage comes from SU3, a stratigraphic unit (SU) at a level lying directly on the natural rocky substrate and preceding the mound construction level. These deposits were archaeologically sterile with the exception of signs of a fire event dated to ca. 11th–12th century ad, interpreted as the result of forest clearance for the establishment of the first occupation mounds in the area (Dotte-Sarout 2010, pp. 237–251; Dotte-Sarout et al. 2013a). An unequivocally strong rainforest signature characterizes the anthracological assemblage and there is a notable absence of savanna or ruderal taxa (Fig. 3). All of the identified taxa are found in New Caledonian rainforests; the most abundant families in the assemblage are typical of these vegetation types, Euphorbiaceae, Myrtaceae, Clusiaceae, Rubiaceae, Meliaceae and Cunoniaceae (Jaffré et al. 1994, 2004). The assemblage displays an average taxonomic diversity for its relatively small size, in comparison with the medium number of taxa and diversity recorded for New Caledonia during the study (Dotte-Sarout 2010; Dotte-Sarout et al. 2015). It comprises 33 taxa from 94 fragments and its Gini–Lorenz index of 29:71 falls within the tropical reference range.


Anthracological spectra from Pwadaunu
There appears to be an over-representation of the two most frequently occurring taxa, Type 1 cf. Symplocos sp./Nemuaron vieillardii representing two endemic taxa with strong anatomical similarities, and a member of the Casuarinaceae, cf. Casuarina collina/Gymnostoma nodiflorum. Both Symplocos sp. and Nemuaron vieillardii are typical of dense tropical rainforests. The two Casuarinaceae share a common association with open, light-demanding and relatively dry vegetation, as mentioned above. Moreover, a small number of taxa, including the secondary taxa Carpolepis laurifolia and Dysoxilum sp, are characteristic of the low and medium altitude photoxerophilous rainforest (Jaffré et al. 1994; Virot 1951). This forest type is distinguished by the presence of more light-seeking taxa adapted to drier conditions than is usually the case in typical rainforests, and is characterized by the dominance of Gymnostoma (Jaffré et al. 1994; Virot 1951). Hence, it appears that the vegetation covering the area just before the establishment of the Kanak occupation site at the turn of the 2nd millennium ad was a diverse rainforest, but a specific open type.
While we observe the presence of this open rainforest, it is interesting to note that, at the turn of the 11th century, the regional climate seems to have changed relatively abruptly from generally dry conditions to the establishment of cyclonic and wet conditions, which continued until the mid 15th century. Following these changes, it appears that the climate entered a phase marked by a strong irregularity in rainfall patterns which has continued up to the present day (Dotte-Sarout 2010).
In the overlying mound construction deposits (SU 2), the first traces of human activity are recorded in the form of (1) pre-contact archaeological remains (Dotte-Sarout et al. 2013a) and (2) the appearance of woody taxa commonly associated with people, such as economic and symbolic trees (Fig. 3, see taxa grouped under “anthropogenic vegetation”). According to the development patterns of Kanak pre-colonial occupation sites, this deposit is interpreted as representing remains from surrounding domestic activities on some early mounds, between the 13th and 14th centuries cal ad. The mound itself is thought to have been constructed at some point during the 15th century ad (Dotte-Sarout et al. 2013a).
These developments appear to have resulted in a substantial transformation of the surrounding forested land. Indeed, the assemblage from SU 2 shows, as well as purely socio-cultural taxa, the appearance and multiplication of trees characterized by a wide variety of ecological conditions and reflecting a clear opening of the forest associated with human impact. There are secondary taxa such as Geissois spp. and savanna taxa such as Melaleuca quinquenervia (Fig. 3). Nevertheless, this transformation happened in parallel with a general persistence of the rainforest, with the photoxerophilous type still being strongly represented and Casuarina collina/Gymnostoma nodiflorum type remaining the most frequent taxon in the assemblage. The most frequent families continue to be those of the main rainforest woody species, Euphorbiaceae and Myrtaceae, then Cunoniaceae, Apocynaceae and Sapindaceae. A third of the taxa (a quarter of the fragments) are restricted to rainforests. The assemblage is a large and rich one with 61 taxa from 300 fragments, and with a satisfactory Gini-Lorenz index of 29:71.
A suite of taxa seems to represent various forms of human impact on the forest, which could be linked to the increasingly open type of vegetation there, in spite of the wetter climatic conditions recorded for the period. Firstly, the appearance of the savanna species Melaleuca quinquenervia and of pioneer rainforest taxa such as Sapindaceae cf. Guioa villosa, Pagiantha serifera and Geissois spp., is notable. Secondly, the assemblage includes rainforest taxa with specific socio-economic value such as Aleurites moluccana, also represented by nut remains, Fagraea berteroana, Ficus sp., Araucariaceae cf. Araucaria columnaris and Syzygium cf. malaccense, three of which produce edible fruits or nuts. There is also the possible presence of important Kanak crops, with Cocos nucifera nut remains, monocotyledon cf. Cordyline sp., monocotyledon cf. Dendrocalamus sp. and Moraceae cf. Artocarpus altilis.
Similarly, SU1, the occupation level of the Pwadaunu mound records the presence of the traditional vegetation associated with pre-colonial Kanak occupation sites, with remains of useful or culturally important taxa such as Cordyline or coconut, as well as hybrid taxa associated with both rainforest and vegetation modified by humans, such as Fagraea berteroana or Ficus spp. (Fig. 3; Table 3). The most abundant families are the Euphorbiaceae, Cunoniaceae, Meliaceae, Apocynaceae and Myrtaceae and these are typically the dominant families represented in the New Caledonian rainforests (Jaffré et al. 1994). Again, there is a clear dominance of photoxerophilous and particularly open and/or dry rainforest types (Fig. 3). It is a large assemblage of 400 fragments that displays a typical high taxonomic diversity for tropical vegetation types, with 66 taxa recorded. The Gini-Lorenz index of 28:72 falls well within the tropical reference range. Nevertheless, the relative proportions of taxa are less differentiated in this phase than in the construction and pre-construction sequences. As well as Casuarina collina/Gymnostoma nodiflorum type, Montrouziera cauliflora is one of the most frequent taxa, one of the Clusiaceae typical of New Caledonia medium altitude rainforests and with a valued timber of high socio-cultural value. The savanna taxon Melaleuca quinquenervia increases in frequency compared to earlier assemblages, showing a more severe disturbance of the forests at the time of occupation of the mound, between the 15th and 18th centuries.
Komijièn
The site of Komijièn is located on an alluvial plain close to the mouth of the river Tiwaka. It was completely surrounded by several hectares of horticultural dry gardening mounds, and there is evidence of considerable landscape changes that included the diversion of streams and the creation of drainage canals. In the three anthracological assemblages studied, a total of 120 plant taxa were identified, including 15 indeterminate types.
In addition to the archaeological excavations described above, three auger cores were taken in a transect across the plain, in order to understand its sedimentary history (Dotte-Sarout 2010, pp. 326–331; Dotte-Sarout et al. 2013a). This survey indicated that what is now a plain used to be a wide estuary bordered by mangroves and a swamp adjacent to the mountainsides. Unfortunately, no dating material was recovered from the deepest sedimentary levels, but radiocarbon dates obtained from charcoal samples from the upper alluvial levels indicate that build-up continued at varying rates of deposition until the 15th century ad, when a Kanak pre-colonial occupation is first recorded (Dotte-Sarout 2010, pp. 326–331). The Kanak occupation appears to have lasted throughout the 16th century, and can be recognized in the SU 4 sedimentary sequences recorded during excavations on the Komijièn mound. The SU 4 deposits and associated anthracological assemblage appear to have resulted from human activities that occurred on the plain before the construction of the settlement and the horticultural structures currently visible in the landscape. The deposits may also have incorporated material from the sides of the adjacent mountains, transported and deposited onto the plain by the nearby streams, before they were channelled in later periods (Dotte-Sarout 2010, pp. 326–331).
The assemblage records the strong presence of rainforest, although of a relatively complex type with taxa adapted to open and dry vegetation (Fig. 4). However mangrove, riverine and halophytic vegetation types, typical of the diverse estuarine ecotones, are also well represented in the assemblage. This ‘mixing’ of ecological zones is well illustrated by the dominance in the anthracological record of families such as Myrtaceae and Sapindaceae that are frequently found in both rainforests and dry forests, Cunoniaceae with taxa that grow in wet rainforest in association with the mangrove family Rhizophoraceae, and the Casuarinaceae family of halophytic, forest and riverine taxa. The assemblage displays a moderately high taxonomic diversity for a relatively small sized sample due to the pedo-anthracological depositional context, with 36 taxa from 100 fragments. The low Gini-Lorenz Index of 36:64 can be linked to the small size of the assemblage in relation to its high taxonomic diversity, which can be principally attributed to the mixture of different vegetation types, from mangrove to rainforest, represented by the charcoal deposit.


Anthracological spectra from Komijien
The absence of the savanna taxon Melaleuca quinquenervia and other taxa associated with vegetation disturbance indicates that the forests had been little affected by human activities at this time, though the archaeological and anthracological data clearly record a human presence. Indeed, although ruderal taxa are absent from the assemblage, a suite of woody taxa is represented which were either cultivated, planted or of socio-cultural importance. These include rainforest or estuarine taxa also traditionally associated with Kanak occupation sites such as Hibiscus tiliaceus, Casuarina equisetifolia, Calophyllum inophyllum or Agathis sp. and Montrouziera cf. cauliflora. The fruit-bearing tree Syzygium malaccense and a monocotyledon, possibly Cordyline, were also present (Fig. 4; Table 3). It is therefore likely that the plain was covered by diverse woody vegetation of mangrove-dominated swamp forest along tidal riverbanks, gradually changing further inland to a rich rainforest on the mountain slopes, interspersed with small pockets of useful or symbolic plant communities.
In the following 17th century ad, a range of archaeological and ethnohistorical data demonstrates an intensification of human occupation on the plain (Dotte-Sarout 2010, pp. 314–362). At this time, it is likely that there were successive extensions to horticultural and habitation structures, with progressive channelling of all streams running through the plain. The SU 3 deposits might record some of these events through the infilling of the mound construction, an event dated to the mid 18th century ad (Dotte-Sarout et al. 2013a). The excavations and material culture show that the anthracological assemblage from this level represents the vegetation exploited by the inhabitants of a Kanak settlement site, maybe consisting of the first mounds of the Komijièn site (Dotte-Sarout 2010, pp. 316–338).
The assemblage is still dominated by rainforest and estuarine vegetation taxa, of which approximately one third also have a ruderal or sociocultural status (Fig. 4). The richest families in the assemblages continue to be those associated with open rainforest and with diverse forest types associated with riverine wet environments or the more exposed mountain slopes, Euphorbiaceae, Myrtaceae, Cunoniaceae, Apocynaceae, Rubiaceae and Sapindaceae. The assemblage is characterized by a high diversity with 74 taxa, and a low Gini-Lorenz index of 32:68 despite the small number of 300 fragments studied. This is again probably related to the varied types of vegetation surrounding the site. Relative proportions between taxa are hence little differentiated, with no taxon being represented by more than 6 % of the fragments. While the diversity of the estuarine vegetation decreases in comparison to previous phases of activity, the forest vegetation becomes more complex. Most of the dominant taxa in the assemblage are commonly associated with open or dry forest types, showing the presence of an open type of lowland forest. It is noteworthy that among the two most frequently occurring taxa is the savanna tree Melaleuca quinquenervia, along with the less well represented Pagiantha serifera or Guioa villosa, typically found on the disturbed edges of rainforests.
Tree crops or woody taxa associated with Kanak occupation sites are also recorded in this phase of activity. These include Artocarpus altilis and Syzygium malaccense, Cocos nucifera and other species with a double halophytic status such as Hibiscus tiliaceus, Calophyllum inophyllum or Thespesia populnea, a suite of rainforest taxa also known for their socioeconomic role like Ficus spp., Elaeocarpus angustifolius, Aleurites moluccana (only represented by nut remains), Araucariaceae cf. Agathis sp. or Montrouziera cf. cauliflora, and the more ubiquitous Fagraea berteroana, Morinda citrifolia and Pandanus sp. As a whole, the SU 3 assemblage demonstrates clear signs of landscape transformation by humans through direct clearance of forest and the establishment of socioeconomic gardens, as well as more subtle manipulation of the taxonomic composition and spatial organisation of the surrounding forest.
The archaeological analysis of Komijièn suggests that the upper layer, recorded as SU 2, is the result of successive late occupations on the mound in the late 18th or early 19th century cal ad. This includes a destruction layer involving a fire that possibly relates to a punitive colonial military expedition against the Kanak sites on the plain in 1862, documented in ethnohistorical records (Dotte-Sarout 2010, pp. 316–338).
The SU 2 assemblage consists of 77 different taxa from 400 fragments, with a very satisfactory Gini-Lorenz index of 25:75. The SU 2 anthracological assemblage is still dominated by taxa of the rainforest, ruderal and socio-economic vegetation and halophytic vegetation (Fig. 4). In the meantime, the number of taxa associated with human activities increases in comparison to the preceding layers, while the taxonomic diversity of the halophytic vegetation decreases. In addition, the evidence for an open and drier type of rainforest becomes stronger. Indeed, the most abundant families in the assemblage are the Euphorbiaceae, Rubiaceae, Sapindaceae, Meliaceae, Myrtaceae and Moraceae, which are a mixture of wet rainforest and dry or open forest types (Jaffré et al. 1994, 2004). The dominant taxa of the assemblage each represent between 5 and 10 % of the fragments (Fig. 4). Apart from the most frequent one, which is an indeterminate type, these are two Rhizophoraceae (mangroves), a group of Casuarinaceae taxa (Casuarina collina/Gymnostoma nodiflora and some fragments of Casuarina cf. equisetifolia) associated with halophilic, riparian and ruderal vegetation types as well as the photoxerophilous rainforest. Also common is Montrouziera cf. cauliflora, typical of the photoxerophilous rainforest but also of great sociocultural importance, valued for its use in the construction of traditional houses. There is a possibility that the charcoal fragments of this taxon in the assemblage represent the remains of such timber destroyed by the fire event. The Type 1 taxon Symplocos arborea/Nemuaron vieillardii also represents rainforest. In addition, the savanna species Melaleuca quinquenervia is represented in just under 5 % of the charcoal fragments. Finally, the SU 2 assemblage records the presence of a suite of ruderal and/or useful trees with various ecological associations, such as Artocarpus altilis, Ficus spp., Aleurites moluccana, Fagraea berteroana, Cocos nucifera, Hibiscus tiliaceus, Calophyllum inophyllum and Pandanus sp.
Hence, the change in the vegetation represented in the anthracological assemblage from SU 2 follows the general trend that is apparent from the first occupation of the mound onwards. The rainforest became an increasingly open and complex vegetation and the diversity in estuarine vegetation decreased through time. The proportion of vegetation associated with human activities increases, not only in the form of savanna and secondary vegetation, but also as a suite of useful trees with low frequency but relatively high diversity.
The upper deposits at the Komijièn site, recorded as SU 1, represent a fairly recent alluvial deposit, substantially disturbed due to post-colonial agricultural and pastoral activities; these were not considered for anthracological analysis.
Discussion
In the two sites presented here, the comparative analysis of the anthracological assemblages reveals a general transformation of the vegetation over time that began early in the 2nd millennium ad, with a clear dominance of rainforest-only taxa. This vegetation changed to a more complex composition with an increased proportion of taxa which represent secondary or ruderal vegetation and crops. Overall, taxa associated with rainforests still remain dominant throughout the time period, from the 12th to the 19th century cal ad, while taxa associated with more open or drier vegetation types are present from the beginning and show a continuous increase over time. At the same time, the number of useful or cultivated taxa increases. Finally, the occurrence of taxa representing savanna and secondary vegetation is clearly associated with the development of the Kanak settlement system and extensive use of horticultural structures, according to associated archaeological data.
By comparing the anthracological results from Pwadaunu and Komijièn with the initial table of useful taxa in Table 3, it is possible to identify the three vegetation areas characterized in the horticultural model defined earlier, from the cultivated to the uncultivated. Indeed, taxa were identified in the assemblages that correspond with the cultivated area and are traditionally planted on Kanak habitation sites for specific symbolic reasons, or as crops in gardens in horticultural areas. Other socio-economic taxa recorded in our literature and field surveys as being associated with the hybrid or forest areas were also identified within the assemblages (Table 3).
In particular, the persistence of photoxerophilous or open rainforests during the 2nd millennium is striking, although the climatic conditions were then progressing towards an increase in rainfall and cyclonic activities. The co-occurrence of many socioeconomic taxa amongst the typical rainforest community suggests that this particular type and taxonomic composition of the forests is a direct result of human manipulation in the form of
-
1.
Construction of drainage channels and associated stream diversion strategies identified during surveys, linked to horticultural features within and around the forests (Dotte-Sarout 2010; Dotte-Sarout et al. 2013a).
-
2.
Precise minimal use of slash-and-burn, and the transplanting or specific care of selected taxa to create an open forest in which more light encouraged production of fruits and nuts, incorporating a variety of significant socioeconomic and symbolic taxa.
Such practices as planting, maintaining and/or protecting certain trees, applying controlled and precise slash-and-burn techniques and the selective cutting and burning of potential plant competitors are described in tropical Asia or in northern Melanesia and Micronesia as traditional methods used to create managed or “domesticated” forests (Athens et al. 1996; Kennedy 2012; Kennedy and Clarke 2007; Latinis 2000; Michon 2005; Walter and Sam 1999). Interestingly, according to the local Kanak word view there is no such thing as “nature”; however, there is a distinction between the forest that is exploited and experienced by people on a daily basis and the “deep forest”, which is not “socialized” and is inhabited by supernatural beings and spirits (Leblic 2005). The forests represented in the anthracological assemblages from the sites of Pwadaunu and Komijièn appear to be those which were regularly visited and exploited by people, in particular for firewood, applying the concept of daily itineraries as defined in Salavert and Dufraisse (2014) and can thus be considered as belonging to the forest domain within the social world. However, the deep forest, which was regarded as dangerous and was not socialized, probably corresponds to the higher altitude dense wet rainforest ecosystems that are not represented in the anthracological assemblages. According to local ethnographic information, one such forest still exists on the Mwîdip mountain top, close to Pwadaunu. Until recently, it was only frequented on very special occasions, for example when a complete tree of Montrouziera cauliflora was needed for the construction of the central pillar of a men’s house, or when a large trunk was to be obtained to build a canoe (Dotte-Sarout 2010, pp. 380–386). However, it was avoided during daily activities and its absence from the anthracological assemblages is to be expected.
This study demonstrates that it is possible to reconstruct the changing spatial structure of the forests based on anthracological data, using a detailed knowledge of the particular local sociocultural systems and social landscape. By understanding the archaeological data within the context of the local and regional environmental and ethnological framework, it is even possible to identify specific forest management practices, especially through two observations:
-
1.
The identification of a suite of taxa with known economic or symbolic uses correlating with archaeological remains of human occupation.
-
2.
The recognition of a specific open type of rainforest, which is seen as the result of human actions, as detailed above.
The fact that this more open forest seems to precede the development of Kanak occupation sites could indicate that specific practices of managing the forest emerged during the preceding millennia of human presence on the island. This, together with the absence of ruderal, savanna or pioneer taxa in the oldest levels analysed, demonstrates that rather than simple deforestation, the major human impact on the forests of New Caledonia until the development of the Kanak sociocultural complex could have been in the form of a precise, but substantial, manipulation of the taxonomic composition and structure of the forests. This can be seen as “domesticating forests” (Michon 2005) or practising a subtle arboriculture or sylviculture that transformed tropical forests into gardens, or “artefacts” as per Barton et al. (2012).
Such a model of vegetation resources management and exploitation makes sense when compared with other regional palaeoenvironmental studies that have shown how the importance of subsistence systems based on trees in Oceania span the entire archaeological sequence, from the late Pleistocene to the late Holocene (Barker 2013; Blench 2005; Denham 2004, 2011; Huebert 2014; Kennedy 2012; Kennedy and Clarke 2007; Kirch and Yen 1982; Latinis 2000; Lepofsky 1992; Michon 2005; Oliveira 2008; Piper and Rabett 2014). Moreover, a comparison between the pan-Pacific plants of socioeconomic importance identified in the New Caledonian assemblages with those from other regional archaeobotanical studies shows interesting correlations. Taxa such as Aleurites moluccana, Artocarpus altilis, Cordia sp., Crossotylis sp., Hibiscus sp., Ficus spp., Syzygium malaccense, Sapindaceae, Terminalia sp. and Thespesia sp., as well as various monocotyledons rarely identified to genera, except for Cocos nucifera, Pandanus sp. and fragments of Araceae tubers, represent an arboricultural corpus that is regularly found in Pacific assemblages, from the early Holocene in Timor and Bismarck, to the late pre-colonial period in Micronesia, New Caledonia and East Polynesia (Athens et al. 1996; Huebert 2014; Kahn et al. 2015; Kirch et al. 2015; Lepofsky 1992; Lepofsky et al. 1996; Millerstrom and Coil 2008; Orliac 1997, 2000, 2003; Oliveira 2008; Weisler 2001).
Moreover, the existence of traditions of forest management makes sense in the context of the immensely diverse and peculiar tropical world, where subsistence systems are based on horticulture, which is the exploitation of diversity and the specific care of individual plants, especially tubers such as Colocasia spp. and Dioscorea spp., essentially associated with the rainforest (Barrau 1962; Barton et al. 2012; Denham 2011).
The central argument presented through the case studies of Komijièn and Pwadaunu is the importance of understanding the anthracological data within their sociocultural as well as socioecological context. This allows proposing interpretations that are based, as much as possible, on local ways and knowledge rather than exclusively on the classical Western academic point of view (Dwyer 1996; Brunois 2005). In this example, this means reconsidering the classical paradigms of what agriculture is, what is domesticated and what is wild, especially so in the tropics. Indeed, numerous scholars have demonstrated that discriminating between domesticated or cultivated and wild or natural taxa is an artificial notion, typically produced by pre-conceived ideas which are derived from cereal-based agricultures and deeply rooted in “Western”, or maybe even non-tropical, representations of the world (Barrau 1962, 1965; Barton et al. 2012; Denham et al. 2007; Descola 2005; Haudricourt 1964; Kennedy and Clarke 2007; Latinis 2000; Michon 2005; Yen 1996). Such a binary division is utterly unadapted to tropical vegetation, landscapes and subsistence modes. It is also foreign to sociocultural representations of the Indo-Pacific region that integrate hybrids of nature and culture with fundamental links between society and the environment (Brunois 2005; Dwyer 1996; Leblic 2005).
Conclusions
There is now in Oceania a considerable body of evidence that demonstrates the existence of horticultural systems which are largely based on the precise management of forests, necessarily avoiding extensive forest clearance. These systems are currently known in most islands of the Indo-Pacific region, although still poorly represented by archaeobotanical data, but have been little studied in New Caledonia, so far. This paper seeks to demonstrate how anthracological data and the use of local interpretative frameworks can be combined to identify the long-term existence of forest-based horticultural systems on the island. This conclusion requires consolidation and refinement by further work. It also calls for attention to several aspects of current classical models used to characterize subsistence systems throughout Oceania, especially at the time of first human settlement. Most notably, it appears that the subtle processes of complex forest transformation should be investigated more thoroughly, considering the non-linear evolution of human impact on the vegetation over time, rather than simply identifying changes in forest structure and composition as reflecting deforestation and destructive “agricultural” practices following initial human settlement. In addition, a single universal model should not be applied to all the islands and sub-regions of the Pacific and Island Southeast Asia. For example, the subtle long-term manipulation of the forests described here seems to be commonly shared by the islands of the western Pacific, while islands of eastern Polynesia tend to demonstrate more abrupt human impacts on the indigenous vegetation associated with arboriculture.
This case study also demonstrates how anthracological studies can go beyond the reconstruction of palaeoenvironment and firewood collection strategies, and be applied to investigate questions of past landscape management and subsistence systems, as demonstrated for instance by Delhon et al. (2009) or Asouti et al. (2015). By comparing the anthracological data with local botanical, ethnological and archaeological data, it is possible to move from the anthracological spectra to perceptions of the spatial organisation of the vegetation and then to interpretations about the horticultural system. Trying to interpret the data within an indigenous socioecological framework not only provides a powerful way of acquiring a deep understanding of past interactions between societies and the vegetation, but it can also help tropical archaeobotany to free itself from long-standing patterns of thoughts that are inherited from its origins in classical Near Eastern and Mediterranean agricultural studies.
References
Asouti E, Kabukcu C, White EC, Kuijt I, Finlayson B, Makarewicz C (2015) Early Holocene woodland vegetation and human impacts in the arid zone of the southern Levant. Holocene 25:1,565–1,580
Athens S, Ward JV, Murakami GM (1996) Development of an agroforest on a Micronesian high island: prehistoric Kosraean agriculture. Antiquity 70:834–846
Barker G (2013) Rainforest foraging and farming in island southeast Asia: The archaeology of the Niah Caves, Sarawak, vol 1. (McDonald Institute Monographs) McDonald Institute for Archaeological Research, Cambridge and Sarawak Museums
Barrau J (1956) L’agriculture Vivrière Autochtone de la Nouvelle-Calédonie. Commission du Pacifique Sud, Nouméa
Barrau J (1962) Les plantes alimentaires de l’Océanie. Origines, Distribution et usages. Musée Colonial, Marseille
Barrau J (1965) L’Humide et le Sec: an essay on ethnobiological adaptation to contrastive environments in the Indo-Pacific area. J Polynesian Soc 74:329–346
Barton H, Denham T, Neumann K, Arroyo-Kalin M (2012) Long-term perspectives on human occupation of tropical rainforests: an introductory overview. Quat Int 249:1–3
Bensa A, Rivierre J-C (1982) Les Chemins de l’Alliance, l’organisation sociale et ses représentations en Nouvelle-Calédonie. SELAF, Marseille
Blench R (2005) Fruits and arboriculture in the Indo-Pacific Region. Bull Indo Pac Pre Hi 24:31–50
Brunois F (2005) Pour une approche interactive des savoirs locaux: l’ethno-éthologie. Journal de la Société des Océanistes 120–121:31–40
Chabal L, Fabre L, Terral J-F, Théry-Parisot I (1999) L’anthracologie. In: Bourquin-Mignot C et al (eds) La Botanique. Errance, Paris, pp 43–104
Delhon C, Thiébault S, Berger J-F (2009) Environment and landscape management during the Middle Neolithic in Southern France: evidence for agro-sylvo-pastoral systems in the Middle Rhone Valley. Quat Int 200:50–65
Denham T (2004) The roots of agriculture and arboriculture in New Guinea: looking beyond Austronesian expansion, Neolithic packages and indigenous origins. World Archaeol 36:610–620
Denham T (2005) Agricultural origins and the emergence of rectilinear ditch networks in the highlands of New Guinea. In: Pawley A, Attenborough R, Golson J, Hide R (eds) Papuan pasts: cultural, linguistic and biological histories of Papuan-speaking peoples. Pacific Linguistics 572. ANU, Canberra, pp 329–361
Denham T (2011) Early agriculture and plant domestication in New Guinea and Island Southeast Asia. Curr Anthropol 52(Suppl 4):S379–S395
Denham T, Iriarte J, Vrydaghs L (2007) Rethinking agriculture: archaeological and ethnoarchaeological perspectives. Left Coast Press, Walnut Creek
Descola P (2005) Par-delà nature et culture. Gallimard, Paris
Détienne P, Jacquet P (1999) Manuel d’identification des bois de Polynésie. CIRAD-Forêt, Montpellier
Dotte E (2010) Modes d’exploitation et intégration au sein des territoires kanak pré-coloniaux des ressources végétales forestières. Approche ethno-archéo-anthracologique en Nouvelle-Calédonie. In: Nicolas T, Salavert A (eds) Territoires et Economies. Publications de la Sorbonne, Paris, pp 155–189
Dotte-Sarout E (2010) «The Ancestor Wood» . Trees, forests and precolonial Kanak settlement on New Caledonia Grande Terre: case study and anthracological approach in the Tiwaka valley. Unpubl. doctoral thesis. Université Paris I Sorbonne and the Australian National University
Dotte-Sarout E, Ouetcho J, Bolé J, Baret D, Sand C (2013a) Of charcoals and forests: Results and perspectives from an archaeobotanical investigation of precolonial kanak settlement sites in New Caledonia. In: Summerhayes G, Buckley H (eds) Pacific archaeology: Documenting the past 50,000 years, vol 25. University of Otago Series in Archaeology, Otago, pp 120–130
Dotte-Sarout E, Sand C, Bolé J, Ouetcho J (2013b) Archéogéographie et processus de territorialisation pré-contact au Nord de la Grande Terre (Nouvelle-Calédonie). Journal de la Société des Océanistes 136–137:139–152
Dotte-Sarout E, Carah X, Byrne C (2015) Not just carbon. Assessment and prospects for the application of anthracology in Oceania. Archaeol Ocean 50:1–227
Dwyer P (1996) The Invention of nature. In: Ellen ER, Fukui K (eds) Redefining nature: ecology, culture and domestication. Berg, Oxford, pp 157–186
Guiart J (1956) L’organisation sociale et coutumière de la population autochtone de la Nouvelle-Calédonie. In: Commission South Pacific (ed) L’agriculture vivrière autochtone de la Nouvelle-Calédonie. Commission du Pacifique Sud, Nouméa
Haberle S, Stevenson J, Prebble M (2010) Altered ecologies: fire, climate and human influence on terrestrial landscapes. Terra Australis 32. ANU Press, Canberra
Haudricourt A-G (1964) Nature et culture dans la civilisation de l’igname: l’origine des clones et des clans. L’Homme 4:93–104
Huebert J (2014) Evaluating forest transformations and the rise of arboreal subsistence strategies in highly variable environments: The anthropogenic forests of the Marquesas Islands, French Polynesia. Unpubl. doctoral thesis, The University of Auckland, Auckland
Jaffré T, Morat P, Veillon J-M (1994) La Flore de Nouvelle-Calédonie. Caractéristiques et composition floristique des principales formations végétales. Bois et Forêts des Tropiques 242:8–28
Jaffré T, Morat P, Veillon J-M, Rigault F, Dagostini G (2004) Composition et caractérisation de la flore indigène de Nouvelle-Calédonie, 2nd edn. IRD, Nouméa
Kahn J, Dotte-Sarout E, Molle G, Conte E (2015) Landscape change, settlement histories, and agricultural practices on Maupiti, Society Islands (Central Eastern Polynesia). J Island Coast Archaeol 10:363–391
Kennedy J (2012) Agricultural systems in the tropical forest: a critique framed by tree crops of Papua New Guinea. Quat Int 249:140–150
Kennedy J, Clarke W (2007) Trees and crops and the cultivated landscapes of the southwest Pacific. In: Connell J, Waddell E (eds) Environment, development and change in rural Asia-Pacific: between local and global. Routledge, New York, pp 76–90
Kirch PV (2000) On the road of the winds: An archaeological history of the Pacific Islands before European contact. University of California Press, Oakland
Kirch PV (2004) Oceanic islands: microcosms of “global change”. In: Redman CL et al (eds) The archaeology of global change. Smithsonian Books, Washington, pp 13–27
Kirch PV, Hunt T (1997) Historical ecology in the Pacific Islands. University Press, Yale
Kirch PV, Yen D (1982) Tikopia. The prehistory and ecology of a Polynesian outlier. (Bishop Museum Bulletin 238) Bishop Museum Press, Honolulu
Kirch PV, Molle G, Nickelsen C, Mills P, Dotte-Sarout E, Swift J, Wolfe A, Horrocks M (2015) Human ecodynamics in the Mangareva Islands: a stratified sequence from Nenega-Iti Rock Shelter (site AGA-3, Agakauitai Island). Archaeol Ocean 50:23–42
Latinis DK (2000) The development of subsistence system models for Island Southeast Asia and Near Oceania: the nature and role of arboriculture and arboreal-based economies. World Archaeol 32:41–67
Leblic I (2005) Sites sacrés et représentatiosn Païcî de la nature (Nouvelle-Calédonie). Journal de la Société des Océanistes 120–121:95–112
Leenhardt M (1953) Gens de la Grande Terre, 2nd edn. Gallimard, Paris
Lepofsky D (1992) Arboriculture in the Mussau Islands, Bismarck Archipelago. Econ Bot 46:192–211
Lepofsky D, Kirch PV, Lertzman KP (1996) Stratigraphic and paleobotanical evidence for prehistoric human-induced environmental disturbance on Mo’orea, French Polynesia. Pac Sci 50:253–273
McKee HS (1985) Les plantes introduites et cultivées en Nouvelle-Calédonie. Hors-série 1, 2nd edn. Museum National d’Histoire Naturelle, Paris
McWethy DB, Whitlock C, Wilmshurst JM et al (2010) Rapid landscape transformation in South Island, New Zealand following initial Polynesian settlement. Proc Natl Acad Sci 107:21,343–21,348
Michon G (2005) Domesticating forests. How farmers manage forest resources. Subur Printing, IRD, CIFOR, ICRAF, Indonesia
Millerstrom S, Coil J (2008) Pre-contact arboriculture and vegetation in the Marquesas Islands, French Polynesia: charcoal identification and radiocarbon dates from Hatiheu Valley, Nuku Hiva. Asian Perspect 47:330–351
Oliveira N. (2008) please add reference Subsistence archaeobotany: food production and the agricultural transition in East Timor. Unpublished doctoral thesis, Australian National University, Canberra
Orliac M (1997) Human occupation and environmental modifications in the Papeno’o Valley, Tahiti. In: Kirch P, Hunt T (eds) Historical ecology in the Pacific Islands. University Press, Yale, pp 200–229
Orliac C (2000) The woody vegetation of Easter Island between the early 14th to the mid-17th centuries AD. In: Stevenson CM, Ayres WS (eds) Easter Island archaeology, research on early Rapa Nui culture. Easter Island Foundation, Los Osos, pp 199–207
Orliac M (2003) Un aspect de la flore de Mangareva au XIIème siècle (archipel Gambier, Polynésie française). In: Orliac C (ed) Archéologie en Océanie Insulaire: Peuplement, Société et Paysages. Art’com, Paris, pp 150–171
Piper PJ, Rabett RJ (2014) Late Pleistocene subsistence strategies in Island Southeast Asia and their implications for understanding the development of modern human behaviour. In: Porr M, Dennell R (eds) Southern Asia, Australia, and the search for human origins. Cambridge University Press, Cambridge, pp 118–134
Salavert A, Dufraisse A (2014) Understanding the impact of socio-economic activities on archaeological charcoal assemblages in temperate areas: a comparative analysis of firewood management in two Neolithic societies in Western Europe (Belgium, France). J Anthropol Archaeol 35:153–163
Sand C (2002) Creations and transformations of prehistoric landscapes in New Caledonia, the southernmost Melanesian Islands. In: Ladefoged T, Graves M (eds) Pacific landscapes: archaeological approaches in Oceania. Easter Island Foundation, Los Osos, pp 11–34
Sand C (2010) Le Lapita calédonien. Archéologie d’un premier peuplement insulaire océanien. Société des Océanistes, Paris
Sand C (2012) ‘Certainly the most technically complex pondfield irrigation within Melanesia’: wet taro field systems of New Caledonia. In: Spriggs M, Addison D, Mathews PJ (eds) Irrigated taro (Colocasia esculenta) in the Indo-Pacific. (Senri Ethnological studies 78). National Museum of Ethnology, Osaka, pp 167–188
Sand C, Bolé J, Ouetcho AJ, Baret D (2008) Parcours archéologique. Deux décennies de recherches du Département Archéologie de Nouvelle-Calédonie (1991-2007). Les Cahiers de l’Archéologie en Nouvelle-Calédonie 17. Département Archéologie, Nouméa
Saussol A (1990) Le Pays Kanak. In: Boulay R (ed) La Maison Kanak. Parenthèses. ADCK-ORSTOM, Nouméa, pp 21–30
Scheel-Ybert R (2000) Vegetation stability in the Southeastern Brazilian coastal area from 5500 to 1400 14C year bp deduced from charcoal analysis. Rev Palaeobot Palynol 110:111–138
Scheel-Ybert R (2001) Man and vegetation in the Southeastern Brazil during the Late Holocene. J Archaeol Sci 28:471–480
Scheel-Ybert R (2002) Evaluation of sample reliability in extant and fossil assemblages. In: Thiébault S (ed) Charcoal analysis: Methodological approaches, Palaeo-ecological results and wood uses. Proceedings of the 2nd international meeting in anthracology, Paris 2000. BAR Internat Ser 1063. Archaeopress, Oxford, pp 9–16
Sémah A-M (1998) Recherche des traces de la première Conquête des Vallées dans Le Nord de la Grande Terre (Nouvelle-Calédonie). Journal de la Société des Océanistes 107:169–178
Stevenson J (2004) A late-Holocene record of human impact from the southwest coast of New Caledonia. Holocene 14:888–898
Virot R (1951) Associations végétales de la Nouvelle-Calédonie et leur protection. Journal de la Société des Océanistes 7:263–269
Walter A, Sam C (1999) Fruits d’Océanie. IRD Editions, Paris
Weisler M (2001) Life on the edge: prehistoric settlement and economy on Utrok Atoll, northern Marshall Islands. Archaeol Ocean 36:109–133
Yen DE (1996) Melanesian arboriculture: historical perspectives with emphasis on the genus Canarium. In: Stevens ML, Bourke RM, Evans BR (eds) South Pacific indigenous nuts. (ACIAR Proceedings 69) ACIAR, Canberra, pp 36–44
Acknowledgments
The research program was funded by an Award doctoral Scholarship from the French Ministry of Research (Allocation de Recherche Doctorale 2005-2008), a cotutelle fellowship from the French Embassy in Australia (2007) and a Cotutelle Lavoisier grant from the French Ministry of Foreign Affairs (2008), with support from the Center for Archaeological Research at the Australian National University, the Groupe De Recherche 2834 and the Equipe d’Ethnologie Préhistorique of the French Centre National de la Recherche Scientifique. Collaboration with the Institute of Archaeology of New Caledonia and the Pacific has been paramount and I also wish to thank Daniel and Irène Létocart for their help and incredible botanical knowledge. I acknowledge the assistance of the Institut de Recherche pour le Développement in Nouméa. The Northern Province of New Caledonia, the Conseils coutumiers of Bopope and Pombéi, and the people of the Tiwaka valley gave permission to work on their lands and are gratefully acknowledged for this, for offering us hospitality and sharing their knowledge of the land. Christelle Carlier provided the background for Fig. 1. Special thanks go to Ingrid Ward and Philip Piper for their helpful editing of the manuscript and to Rita Scheel-Ybert for providing the impetus for the writing of this paper. Two reviewers also gave constructive comments to improve the quality of the paper. Part of this research was conducted at the University of Western Australia.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by A. Fairbairn.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Dotte-Sarout, E. Evidence of forest management and arboriculture from wood charcoal data: an anthracological case study from two New Caledonia Kanak pre-colonial sites. Veget Hist Archaeobot 26, 195–211 (2017). https://doi.org/10.1007/s00334-016-0580-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-016-0580-0