
Abstract
Charcoal fragments from five historic campsite locations in the Galápagos Islands were identified and radiocarbon dated to investigate postulated early human presence in the archipelago, historic fuel wood collection patterns and the resultant impact on native vegetation. A variety of taxa and fuel types were revealed to be present in the charcoal assemblages, indicating geographically driven rather than species-specific methods of collection. Historic anthropogenic impact was therefore spread amongst woody taxa in the lowland plant communities, with severity dependent on proximity to campsite location. All charred remains were found to date from within the historic period, supporting the preponderance of archaeological evidence indicating that human presence did not begin in Galápagos until after European discovery.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Human impact poses one of the greatest threats to the native biodiversity of the Galápagos Islands, an area renowned for its unique ecosystems (Snell et al. 2002). Recent habitat degradation can be attributed to increasing human habitation, agricultural development, anthropogenic introduction of non-native species, and increases in tourism and its associated infrastructure (Boersma et al. 2005). Little, however, is known about the history of early human impact in Galápagos. The first known human presence in this remote island archipelago, located 1,000 km west of the South American mainland, occurred with European discovery in a.d. 1535. Archaeological evidence of surface and subsurface earthenware pottery has led to speculation, most notably by Heyerdahl and Skjölsvold (1956), about earlier human habitation in Galápagos. They contended that some ceramic pot sherds discovered in coastal campsite locations were of pre-Columbian Amerindian origin. They referred in particular to a type of unglazed, thin-walled, red-slipped earthenware, which they called ‘aboriginal ware’. Their conclusions, however, have been questioned both on methodological grounds (Suggs 1967) and as a result of recent changes in the archaeological classification of South American pottery (Flett and Haberle 2008; Stothert 2007).
This paper examines charcoal fragments recovered during an archaeological investigation of coastal campsites in June 2005 (Anderson and Haberle 2005). Whilst most of the site locations examined had previously been investigated by earlier archaeological expeditions to Galápagos, including those of Heyerdahl and Skjölsvold (1956), our excavations were within areas of archaeologically undisturbed sediments (except for material from area L, James Bay described below) (Anderson 2005). Radiocarbon dating of charred woody remains found in context with putative aboriginal ceramics is used to examine the possibility of pre-European human presence in Galápagos. Identification of the charcoal fragments provides information on historic fuel wood collection, that is, whether particular species were favoured as a fuel source, and what impact this may have had on the native vegetation.
The Galápagos archipelago was discovered by chance in a.d. 1535 by the Bishop of Panama and documented visits to the rather inhospitable islands throughout the remainder of the 16th century were infrequent. Human presence increased throughout the 17th and 18th centuries with the arrival of buccaneers and whalers (Perry 1984; Slevin 1959), followed by the scientific voyages of the 19th century including Darwin’s famed expedition on the Beagle in 1835. Human occupation of the islands throughout this time was largely transient, with impacts centred upon temporary coastal campsites established at sites with good anchorage and near to scarce freshwater sources. Some small, short-lived settlements developed in the islands during the early 19th century, most notably on the islands of Floreana and San Cristóbal. Reported early Galápagos inhabitants include individuals left behind by passing ships, penal colonies of political deportees and convicts from mainland Ecuador, and settlers engaged in trade with visiting whalers (Hickman 1985). Permanent habitation of most of the larger islands did not begin until the late 19th and early 20th centuries.
Of the two islands examined in this study, Santa Cruz is the more developed, presently supporting the largest population centre in Galápagos and extensive agricultural development in the island’s interior (Snell et al. 2002). The earliest reported habitation on Santa Cruz is at Whale Bay on the west coast, where historical accounts describe the presence of some huts in the 1840s (Lundh 2004). Apart from a few small settlements, significant permanent human occupation on Santa Cruz did not begin until the development of farming and fishing communities in the 1920s and 1930s. Santiago Island is one of the largest uninhabited islands in the Galápagos archipelago. It was a favoured base for buccaneers in the 17th century, and in the 1920s and 1960s salt mining operations were established on the island, but soon abandoned. Whilst never developed for settlement or agriculture, the island’s vegetation has been indirectly impacted by humans through the introduction of feral goats as a food supply for passing ships.
The impact of early visitors on the native fauna of the islands, particularly hunting of the famed Galápagos tortoises which were taken as a source of fresh meat aboard ship and later hunted for their oil, has been well documented, but little is known about the impact early inhabitants may have had on the native vegetation. Historical accounts document both the use of local animals as a food source and of the collection of wood for fuel (Burney 1816 in Burney 2002; Dampier 1697 in Dampier 1998), but there is little information about the specific trees and habitats which were utilized. The collection method, either preferential harvesting by species based on factors such as burn quality or log size, or geographically determined patterns of gathering, would have had very different impacts upon the island vegetation.
Presently, the vegetation on the larger, more topographically complex islands of the Galápagos exhibits a distinct altitudinal zonation pattern which is commonly sub-divided into four zones: Littoral, Arid, Transition and Humid (Johnson and Raven 1973; McMullen 1999; Wiggins and Porter 1971). The woody vegetation growing along the coastal fringe in the Littoral Zone is dominated by four species of mangrove (Avicennia germinans, Conocarpus erectus, Laguncularia racemosa and Rhizophora mangle). The Arid Zone encompasses the dry lowland communities, typically extending inland up to an elevation of 120 m on wetter southern slopes and to over 300 m on northerly aspects (Wiggins and Porter 1971). The native vegetation is dominated by cactus at the lowest elevations, and by open-canopy forests of Bursera graveolens. Other characteristic woody plants include Acacia, Cordia lutea, Croton scouleri, Parkinsonia aculeata, Piscidia carthagenensis and Prosopis juliflora as well as numerous shrubs, often forming dense patches of undergrowth. Characteristic trees occurring in the higher elevation Transition and Humid Zones include Pisonia floribunda, Psidium galapageium and Zanthoxylum fagara (McMullen 1999). The Humid Zone may be sub-divided into three natural vegetation types: the endemic ‘giant daisy tree’ forests of Scalesia pedunculata, a community that has been severely impacted by agricultural development and grazing of introduced animals, Miconia scrub, and high elevation fern-sedge communities.
The aim of this study was to examine historic anthropogenic impact on the native vegetation of the Galápagos Islands, specifically:
-
(i)
To determine the age of charred woody remains recovered during archaeological excavations of historic coastal campsite locations in order to date human occupation.
-
(ii)
To identify the woody taxa being utilised.
-
(iii)
To determine historic fuel wood collection patterns in Galápagos and whether there was selection of specific wood types, geographic dispersal and possible impacts on the local vegetation composition.
Methods
Sample collection
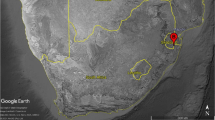
Charcoal fragments from archaeological campsite assemblages were excavated from five coastal sites in June 2005: James Bay and Buccaneer Bay on Santiago Island; Whale Bay, Las Palmitas and Cabo Colorado on Santa Cruz Island (Fig. 1). Sampling was undertaken at 10 cm depth intervals where possible, but most sites exhibited little or no stratigraphy. Charcoal was collected both from within defined hearths and also from the surrounding campsite locations. Representative samples were collected from all material found which appeared to be in primary context and dry-sieved to remove debris. The size and number of charcoal fragments recovered ranged widely between sites (Table 1).

Locations of the five archaeological sites, Santiago and Santa Cruz Islands, Galápagos
Although there are presently no permanent settlements on Santiago Island, anchorage had been sought at various times there since at least the 1681 visit by Captain Dampier to James Bay on the west coast (Fig. 1). James Bay is one of the few coastal locations in Galápagos with a relatively dependable source of fresh water (Slevin 1959) and Heyerdahl and Skjölsvold (1956) postulated that it is the best-suited location in the archipelago for supporting possible pre-European settlement. The excavation site is located on a raised coastal terrace with evidence of multiple campsites containing both earthenware pottery sherds, notably of ‘aboriginal ware’ and historic European remains, including glazed and unglazed ceramics, glass, nails and other iron fragments. Heyerdahl and Skjölsvold (1956) subdivided the site into twelve sampling sections designated A–L. That designation is followed here and we have analyzed charcoal obtained from three of the sections, Areas E, H and L. Surface charcoal samples were collected from Area E, a narrow, steeply-sloping shelf located above the central beach which may have been used as a refuse dump or simply caught archaeological material sliding down from a campsite nearby. Area H, containing a fireplace and associated midden, is located on a neighbouring hill slope. Charred remains were excavated from inside the fireplace at 10–20 cm depth and outside the fireplace at progressive depths (Table 1). Area L is a separate campsite located on the eastern edge of the James Bay site and the charcoal samples were obtained from a residual soil heap resulting from Heyerdahl and Skjölsvold’s original excavation.
Buccaneer Bay is on the northwest tip of Santiago (Fig. 1). It is another known historic campsite location most famously occupied by Charles Darwin in 1835 on one of his longest sojourns ashore in Galápagos (Darwin 1839 in Darwin 1989). The site contains multiple hearths and ‘aboriginal ware’ as well as early 19th century remains. Charcoal samples were recovered from archaeological ‘Test Pit 4’, located in an area containing the putative early pottery, at 30–40 cm depth and also from ‘Hearth 2’, a 4 m2 excavation site containing multiple fireplaces, an associated midden and ceramics. Samples were collected from Fireplace 1 at 40–60 cm depth and Fireplace 2 at 50 cm (Anderson 2005).
Three sites were excavated on Santa Cruz Island (Fig. 1). Whale Bay is located on the west side of the island, a site of reported historic occupation (Lundh 2004; Slevin 1959). The archaeological site, located on a plateau above the beach, contains abundant ‘aboriginal ware’ and early 19th century material (Heyerdahl and Skjölsvold 1956). The surface of the site was sampled on a 5 m × 5 m grid (square A1 in the southeast corner), with test excavations of 0.5 m × 0.5 m and sampling at 10 cm intervals in selected places. Surface charcoal fragments were analysed from sampling squares G4, G5, G10 and H9. The second site on Santa Cruz, Las Palmitas, is a sheltered harbour on the west coast, located approximately 8 km south of Whale Bay. This site had not been excavated previously. A surface fireplace with a midden of chiton and turtle shells and early 20th century bottle-glass remains was discovered there. The site is approximately 10 m long and all of the charcoal was recovered from a small surface fireplace. The final campsite location examined was Cabo Colorado on the north-eastern coast of Santa Cruz, a site of reputed aboriginal pottery discovered by the Walt Disney Galápagos expedition in 1954. Material, including European glass and ceramics and sherds of ‘aboriginal ware’ was collected from the surface and logged in a grid system, as at Whale Bay, and test pits of 0.5 m × 0.5 m were excavated in selected areas. Surface charcoal samples were collected from within and adjacent to a fireplace with an associated midden.
Identification of charred remains
Reference material
A wood anatomy reference collection was developed in the Long-term Ecology Laboratory, School of Geography and the Environment, University of Oxford. Wood slices 12–25 μm thick were taken using a sliding microtome from branch and stem material of 29 of the most common native woody plants on Santa Cruz and Santiago Islands. Anatomical descriptions and photographs available in the InsideWood online wood anatomy database (InsideWood 2004) and reference slides in the Jodrell Laboratory, Kew Gardens were used to examine other native and introduced taxa occurring in Galápagos. Although branch wood is known to be more anatomically variable than that of tree boles (Schweingruber 1990), collection restrictions necessitated its use and samples were shown to exhibit a full range of material from bark to pith. Tissue sections along three axes (transverse, tangential and radial) were stained using safranin and astrablue, washed with ethanol and xylene, fixed with Canada balsam and dried overnight at 60°C. Anatomical characteristics were described for each species according to the International Association of Wood Anatomists (IAWA) standard (1989). The complete IAWA list of anatomical features and sectional view photographs of each species have been included in the InsideWood database (InsideWood 2004) and may be accessed at http://insidewood.lib.ncsu.edu/search.
Charcoal identification
Charcoal identification follows the methodology of Leney and Casteel (1975). Fragments were split with a razor blade to obtain transverse, tangential and radial sections and examined under a reflected light microscope at ×50, ×100, ×200 and ×500 magnification.
Radiocarbon dating
Nine charcoal samples obtained from four of the analysed sites were radiocarbon dated to determine the age of the woody material and the inferred date of campsite occupation (Table 2). Calendar ages were calibrated based on the SHCal04 dataset (McCormac et al. 2004) and calculated at the 2σ level using the probability distribution method, CALIB 5.0.1 (Stuiver and Reimer 1993). Median calendar ages and standard deviations are based on a 95% minimum probability of occurrence and are rounded to the nearest 5 year interval. In two cases, individual fragments were too small for dating and two fragments had to be combined within the radiocarbon sample (fireplace 1, hearth 2 at Buccaneer Bay; and Cabo Colorado, surface collection, outside the fireplace). In each case, two charred fragments of the same taxon exhibiting similar characteristics (collection position, size, colour, patterning and other physical qualities, that is, powdery or glassy, hard or flaky) were selected to increase the likelihood that the remains had come from the same original source material.
Results
A total of 127 charcoal fragments were examined (Table 1). 15 fragments were found to be too degraded for analysis. 11 distinct wood anatomical types (Table 3; Fig. 2) were identified from the remaining 107 specimens: six positive and one provisional species identifications were made, whilst four distinct types remained unidentified. One of these was tentatively classified as Chiococca alba, but the identification could not be positively confirmed from only the single fragment recovered. The effect of carbonization is known to distort some wood anatomical features, but most major features are generally preserved well enough to allow taxonomic identification (Rossen and Olsen 1985). Anatomical features observed in the charcoal samples for each identified taxon are summarised in Table 4. Three of the four remaining unidentified wood types were found not to be abundant within the campsite assemblages, with only one or two fragments present at single locations (Table 3). A fourth unidentified taxon, however, was recovered from two separate archaeological sites on Santa Cruz, Cabo Colorado and Las Palmitas, with 11 fragments of this type comprising the entire Las Palmitas assemblage. The charcoal fragments of this type exhibit short uniseriate rays (tangential longitudinal view), square and upright ray cells (radial longitudinal view), vessels both solitary and in pairs (transverse view), scanty paratracheal and in places aliform and confluent parenchyma (transverse view), and alternate intervessel pitting (tangential longitudinal view). It is likely that this species may be Croton scouleri, an endemic small tree or shrub the most common variety of which, Croton scouleri var. scouleri, is found in the arid lowlands throughout the Galápagos archipelago, although a positive identification could not be made from the charred remains.


Microscopic view of charcoal fragments of the seven identified and four unidentified taxa discovered within the archaeological remains. a Vallesia glabra (TS, ×200); b Bursera cf. graveolens (TS, ×100); c Leguminosae subfamily Mimosoideae (TS, ×100); d Piscidia carthagenensis (TS, ×100); e Scutia spicata cf. (TLS, ×200); f Cinchona pubescens (TS, ×100); g monocot (TS, ×100); h unidentified taxon 1 (Chiococca alba cf.) (TS, ×100); i unidentified taxon 2 (TS, ×100); j unidentified taxon 3 (TS, ×100); k unidentified taxon 4 (TS, ×100); l unidentified taxon 4 (RLS, ×200). TS transverse section, TLS tangential longitudinal section, RLS radial longitudinal section; (images: J.A. Lee)
Three charcoal types were identified to be woody material derived from species of large trees, Bursera cf. graveolens, an Arid Zone dominant, Piscidia carthagenensis and Cinchona pubescens, an introduced species. Abundant charcoal from a Mimosoid legume was also revealed (Fig. 2), but could not be identified to species level. This is likely to be from either Acacia or Prosopis, both of which are common small trees of the coastal Arid Zone. Charcoal fragments of a number of shrubs were found, including Vallesia glabra, Scutia spicata var. pauciflora cf. and possibly also Chiococca alba, in addition to the charred remains of an unidentified monocot.
The radiocarbon results show that all of the dated charcoal fragments, apart from one sample at James Bay, originated during the historic period, with median calibrated ages ranging between a.d. 1575 and a.d. 1825 (Table 2). The James Bay sample, a charcoal piece excavated from a fireplace within sampling Area H (fragment JAM-HFIR_1), was found to be beyond the range of the radiocarbon calibration curve and can therefore only be identified as modern (post-1950) material (Table 2). This sample is one of eight fragments of monocot recovered from the James Bay site (Table 3). The charred remains are probably the result of recent use of the campsite and burning of either an introduced taxon or imported material. All of the radiocarbon samples in the campsite assemblages post-date European discovery of the Galápagos Islands. The full range of calibrated ages of the charcoal samples, conservatively calculated at the 2σ level with a 95% probability of occurrence, range between a.d. 1500 and a.d. 1950 (Fig. 3).

Charcoal fragment age determinations. Calibrated age ranges displayed provide a minimum 95% probability of occurrence
Discussion
The charcoal identifications reveal the variety of taxa utilized as fuel wood at the archaeological sites (Table 3). Las Palmitas was the only site containing charred remains derived from only a single taxon. The other sites had more diverse charcoal assemblages, with up to six taxa shown to be present at James Bay. Acacia/Prosopis was the most common type utilized, with charred remains present at five of the six sites. The assemblages were shown to contain a mix of types of burned material, with charcoal of larger trees such as Bursera graveolens and Piscidia carthagenenis, smaller common tree taxa such as Acacia/Prosopis, and shrubby species including Scutia spicata and Vallesia glabra.
The charcoal identification reveals a general ‘least effort’ pattern of firewood collection in Galápagos. The remains of eleven distinct taxa reveal the burning of a range of common lowland trees and woody shrubs. There do not appear to be any particularly favoured types, the selective harvesting of which could have led historically to observable impacts on heavily utilized native species. Rather than species-based impacts, anthropogenic effects on the native Galápagos vegetation would instead have been geographical, centred around historic campsite locations. Acacia/Prosopis, both good burning and abundant Arid Zone taxa, was the most common type discovered in the charcoal assemblages. Evidence for the burning of larger trees such as Bursera graveolens and Piscidia carthagenensis was also revealed in the charred remains, although these species were less dominant in the campsite assemblages than might have been expected. The use of shrubs is probably under-represented in the charcoal remains as a result of the more complete combustion of smaller woody material. The presence of charcoal of small trees and shrubs supports the geographic collection hypothesis, rather than a bias towards larger trees to burn as logs. There is no evidence of more distant collection from the interior of the islands.
The small amount of Bursera wood that was found in the charcoal assemblages from throughout the island campsites is surprising, considering the ubiquitous distribution and large log size of this Arid Zone tree. In fact, Bursera charcoal was only discovered at one site, James Bay, at which it accounted for only 24% of the total assemblage (Table 3). The size and availability of Bursera would make it one of the more likely taxa for collection, if a concerted effort was to be made to harvest larger fireplace logs. Bursera is known to be a favoured firewood in mainland Ecuador, both because it burns well and also as a result of its mosquito-repelling properties (Jaramillo, personal communication). The lack of charred Bursera remains in the Galápagos assemblages indicates that the sites were only used as temporary camps where firewood collection was restricted to local, expedient gathering, rather than the targeted harvesting of larger logs as might be expected with longer-term habitation.
Charred remains of Piscidia carthagenensis were discovered at sites on both Santa Cruz and Santiago (Table 3). Piscidia is a native of Santa Cruz and San Cristóbal islands in the archipelago, as well as Central and South America, but it is not known to occur on Santiago. Whilst found in relative abundance in the campsite assemblages of the Santa Cruz Island sites, the sample recovered at Buccaneer Bay on Santiago is a single, small fragment. Piscidia is an extremely hard wood favoured for housing and boat construction in Galápagos (McMullen 1999). The common local use of the wood, coupled with discovery of only the single charred piece, suggests that the Piscidia remains found on Santiago are probably the result of the burning of ship’s timbers or other transported wooden products which had been brought over from a neighbouring island or the mainland, although the possibility that it may represent evidence of the past local occurrence of the species on Santiago cannot be discounted.
Burning of the non-native species Cinchona pubescens was revealed in the surface campsite collection at James Bay (Table 3). Cinchona is an escaped cultivar introduced to Galápagos ca. 1946 (Hamann 1974), which has since spread throughout the highlands of Santa Cruz Island. The presence of Cinchona charcoal at James Bay on Santiago Island, where the species does not occur, indicates both the transport of an object or fuel wood for burning and also the likelihood of more recent use of the campsite. This is supported by the radiocarbon evidence which revealed burning at James Bay during the modern period (Table 2). The charcoal could have originated from recent use of the campsite by goat hunters, park wardens and scientists (Tye, personal communication).
The range of probable ages for the nine radiocarbon dated charcoal fragments (Fig. 3) demonstrates that all originated within the historic period. Calendar age determinations were calculated using methodologies providing a high probability of inclusion within the calibrated period, thus ensuring accuracy, but also leading to increasing age range estimates. Most of the uncalibrated 14C dates fall within the modern plateau for radiocarbon dating, leading to decreased precision in the calibration (Table 2). An additional factor in dating charcoal is that the fragments being dated may have come from large trees, thus possibly yielding dates of origination which may be several centuries older than the actual burn period. The oldest charcoal remains revealed by this study (Whale Bay, grid square H9) were from a sample of the large tree species Piscidia carthagenensis dating to between a.d. 1500 and 1650, still predominantly within the historic period. Despite the combination of factors tending to increase calibrated age range estimates, the campsite assemblage results remain remarkably similar—clearly demonstrating that the burning took place within the historic period.
The sites analysed contained ‘aboriginal ware’ in association with the dated charcoal assemblages. As the sites consist largely of remains lying on the surface, or within the uppermost sediments, the association of pottery and charcoal was generally surficial, and therefore not as strong as that from within sealed stratigraphy. However, where samples were sealed stratigraphically, as at James Bay and Buccaneer Bay, the radiocarbon dates were equally young. The radiocarbon results contradict inferences of pre-European occupation drawn from typological analysis of pottery by Heyerdahl and Skjölsvold (1956) and support the conclusion developed upon the preponderance of recent archaeological data (Anderson 2005; Stothert 2007; Flett and Haberle 2008) that human impact did not begin in Galápagos until after European discovery.
References
Anderson AJ (2005) Fieldbook and analytical notes and plans of the 2005 Galapagos Islands expedition. Department of Archaeology and Natural History, Australian National University, Australia
Anderson AJ, Haberle SG (2005) Human colonisation and environmental change on the Galapagos Islands, Republic of Ecuador: report to the El Instituto Nacional de Patrimonio Cultural (INPC), Guayaquil, Ecuador, May–June 2005
Boersma PD, Vargas H, Merlen G (2005) Living laboratory in peril. Science 308:925
Burney J (1816) A chronological history of the discoveries in the South Sea or Pacific Ocean, vol 4. History of the Buccaneers of America. G & W Nicol, London
Burney J (2002) History of the Buccaneers of America. Dover Publications, Mineola
Dampier W (1697) A new voyage round the world. Printed for James Knapton, London
Dampier W (1998) A new voyage round the world: the journal of an English Buccaneer; foreword by Giles Milton. Hummingbird Press, London
Darwin C (1839) Journal of researches into the natural history and geology of the countries visited during the voyage of H.M.S. “Beagle” round the World, under the command of Capt. Fitzroy, R.N., from 1832 to 1836. Henry Colburn, London
Darwin C (1989) Voyage of the Beagle: Charles Darwin’s journal of researches; edited and abridged with an introduction by Janet Browne and Michael Neve. Penguin, London
Detienne P, Jacquet P (1983) Atlas d’identification des bois de l’Amazonie et des regions voisines. Centre Technique Forestier Tropical, Nogent-sur-Marne
Evans JA, Gasson PE, Lewis GP (2006) Wood anatomy of the Mimosoideae (Leguminosae). (IAWA Journal Supplement 5). IAWA, Leiden
Flett I, Haberle SG (2008) East of Easter: traces of human impact in the far-eastern Pacific. In: Clark G, Leach F, O’Connor S (eds) Islands of inquiry—colonisation, seafaring and the archaeology of maritime landscapes. ANU E. Press, Canberra, pp 281–299
Gasson P, Wray E, Schrire BD (2004) Wood anatomy of the tribe Millettieae with comments on related papilionoid Leguminosae. IAWA J 25:485–545
Hamann O (1974) Contributions to the flora and vegetation of the Galápagos Islands. III. Five new floristic records. Botaniska Notiser 127:309–316
Heyerdahl T, Skjölsvold A (1956) Archaeological evidence of pre-Spanish visits to the Galápagos Islands. (Memoirs of the Society for American Archaeology 12). Society for American Archaeology, Salt Lake City
Hickman J (1985) The enchanted islands: the Galapagos discovered. Anthony Nelson, London
IAWA (International Association of Wood Anatomists) Committee (1989) IAWA list of microscopic features for hardwood identification. IAWA Bull 10:219–332
InsideWood (2004-onwards) Published on the Internet. http://insidewood.lib.ncsu.edu/search
Johnson MP, Raven PH (1973) Species number and endemism: the Galápagos archipelago revisited. Science 179:893–895
Leney L, Casteel RW (1975) Simplified procedure for examining charcoal specimens for identification. J Archaeol Sci 2:153–159
Lundh JP (2004) Galápagos: a brief history. Unpublished manuscript. Oslo, Norway. http://www.galapagos.to/TEXTS/LUNDH-0.HTM
McCormac FG, Hogg AG, Blackwell PG, Buck CE, Higham TFG, Reimer PJ (2004) SHCal04 southern hemisphere calibration 0–11.0 cal kyr BP. Radiocarbon 46:1087–1092
McMullen CK (1999) Flowering plants of the Galápagos. Cornell University Press, Ithaca
Neumann K, Schoch W, Detienne P, Schweingruber FH (2001) Woods of the Sahara and Sahel: an anatomical atlas. Paul Haupt, Bern
Perry R (1984) The islands and their history. In: Perry R (ed) Galapagos. Pergamon, Oxford, pp 1–15
Rossen J, Olsen J (1985) The controlled carbonization and archaeological analysis of SE U.S. wood charcoals. J Field Archaeol 12:445–465
Schweingruber FH (1990) Microscopic wood anatomy: structural variability of stems and twigs in recent and subfossil woods from central Europe. Eidgenössische Forschungsanstalt WSL, Birmensdorf
Slevin JR (1959) The Galapagos Islands: a history of their exploration. (Occasional papers of the California Academy of Sciences No. 25). California Academy of Sciences, San Francisco
Snell HL, Tye A, Causton CE, Bensted-Smith R (2002) The status of and threats to terrestrial biodiversity. In: Bensted-Smith R (ed) A biodiversity vision for the Galapagos Islands. Charles Darwin Foundation and World Wildlife Fund, Puerto Ayora, Galapagos, pp 30–47
Stothert KE (2007) Report on pottery from Galapagos (2005 Galapagos expedition). The University of Texas, San Antonio
Stuiver M, Reimer PJ (1993) Extended 14C database and revised CALIB 3.0 14C age calibration program. Radiocarbon 35:215–230
Suggs RC (1967) A reanalysis of Galapagos ceramics data. Zeitschrift für Ethnologie 92:239–247
Wiggins IL, Porter DM (1971) Flora of the Galápagos Islands. Stanford University Press, Stanford
Acknowledgments
This research was funded by the UK Natural Environment Research Council (grant NE/C510667/1 awarded to K.J. Willis and C.A. Froyd) and by the Australian Research Council (grant DP0449560 awarded to S.G. Haberle and A.J. Anderson). The authors would like to thank the Charles Darwin Research Station and the Galápagos National Park Service for their support of the project. The research was carried out under Autorizacion No. 002.SRL.INPC.2005 issued by the Instituto Nacional Patrimonio Cultural, Ecuador, to whom thanks are also due We gratefully acknowledge the contributions of additional archaeological colleagues in the 2005 expedition; Rosanne Anderson, Helene Martinsson-Wallin, Karen Stothert, Paul Wallin and César Vientamille and also Julia Sonsin Oliveira for assistance in charcoal identification. We would like to thank Pim van der Knaap, Alan Tye and an anonymous reviewer for their helpful comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by F. Bittmann.
Rights and permissions
About this article
Cite this article
Froyd, C.A., Lee, J.A., Anderson, A.J. et al. Historic fuel wood use in the Galápagos Islands: identification of charred remains. Veget Hist Archaeobot 19, 207–217 (2010). https://doi.org/10.1007/s00334-010-0239-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-010-0239-1