
Abstract
The study of plant macro-remains from archaeological sites provides substantial information on the activities occurring in a settlement and living conditions during its occupation. This article reports the plant macro-remains (charred seeds and wood charcoal) recovered from a rock shelter in the temperate forests of southern Chile (Latitude 39°S). The main goal of the study was to assess the potential of these remains to indicate collection, use and consumption of plant resources available in this ecosystem by hunter-gatherer groups during the Holocene. Remains from three cultural periods were collected using wet sieving and flotation techniques. Macro-botanical remains found in the rock shelter represented a variety of native trees, shrubs and herbs, both native and introduced. Charred seeds included 19 plant taxa, divided into three groups according to their source: collected foods, including pulses (1 species) and grains (3 taxa, 1 probably cultivated); seeds introduced with food items, including fruit stones and nutshells from several native shrubs and trees; seeds introduced with non-food items, mainly from tree species with no evident use. Wood charcoal fragments represented 28 different native taxa (26 dicotyledons, 1 monocotyledon and 1 gymnosperm), none with a specific identifiable use. Analysis of seed remains indicated the use of grains and, to some extent, collection of fruits and nuts over summer and autumn. The analysis of dispersed charcoal fragments and those accumulated in hearths were especially useful to describe the vegetation surrounding the rock shelter, infer successional events, and identify taxa undetectable in the seed record.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Insights into the subsistence strategies of hunter-gatherer groups in the temperate forests of southern Chile (35–55°S) are limited. Until now, only a few isolated sites have been located and thoroughly studied. It is in the valleys of the Calafquen region, in the southern Chilean Andes, where the earliest settlements have been recorded (Mera and Garcia 2005). The study of two rock shelters in this area has provided valuable information about hunter–gatherer groups and their ways of life in this ecosystem (Adán et al. 2004; Mera and Garcia 2005). Analysis of biological remains has shown that subsistence of these groups involved the direct procurement of animals from the wild, such as small and medium size mammals, birds from aquatic environments and freshwater mussels (Velásquez and Adán 2004). Hunting was probably assisted by the use of traps (Velásquez and Adán 2004). Collection of edible plants and other plant resources could have also been an important component of their subsistence strategy. In fact, numerous ethnobotanical, ethnohistorical and ethnographical studies have confirmed that peoples inhabiting this region had an extensive knowledge of the many plant resources available there. A review by Smith-Ramírez (1995) shows that 78–97% of the native flora occurring in these forests is still used by many indigenous communities; this included not only plants with nutritional value but also those used in rituals, popular medicine, tools, construction and fuel (Gonzalez and Valenzuela 1979; Bragg 1981; Aldunate and Villagrán 1992; Smith-Ramirez 1995; Villagrán 1998; Rapaport and Ladio 1999).
The analysis of biological remains from archaeological sites has become a regular and important aspect of many archaeological studies. Analysis of these ecofacts provides significant information about human activities and living conditions in the settlements, as well as past environmental conditions (Kenward and Hall 1997). Plant remains such as wood charcoals and charred seeds have also been helpful to define mobility strategies of hunter–gatherer groups (Bonzani 1997), identify plant resources available in the area (Rodriguez 2000, 2004), reconstruct past plant communities (Asouti 2003) and also indicate economic activities between groups (Weiss and Kislev 2004). Despite the importance of these analyses, archaeobotany has been poorly developed in Chile, and it is restricted to the northern regions (Planella and Tagle 1998; Quiroz 2001) and Patagonia (Solari 1991, 1993, 1994, 2003; Solari et al. 2002). The scarcity of archaeobotanical studies is especially evident in southern Chile where only data from two sites are available (Ramirez 1989; Dillehay and Rossen 1997; Rossen and Dillehay 1997; Quiroz and Belmar 1998). In this article we report the macro-botanical remains, wood charcoal and charred seeds, recovered from a rock shelter in the temperate forest of southern Chile (39°S). The main goal of our study was to assess the potential of plant macro-remains to indicate activities of collection, use and consumption of plant resources by hunter–gatherer groups inhabiting these forests.
Environmental setting
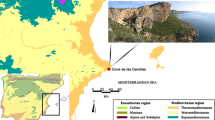
The Calafquén region, in southern Chile, is part of a major ecological area in which lakes, volcanoes and forests dominate the landscape (Thomasson 1963). Almost 65 archaeological sites have been recently discovered in this area, accounting for almost 10,000 years of human occupation (Pino et al. 2004). After an exploratory campaign to determine the potential of each of these sites, efforts were focused on the Marifilo 1 rock shelter, located in a valley of the Chilean Andes (39°S) at c. 300 masl, 1,260 m north of the shore of the Lago Calafquén and about 16 km from the Villarica Volcano (Fig. 1). This volcano has remained active for most of the late Pleistocene and Holocene, and at least 59 eruptions have been reported since 1558 (Petit-Brehuil and Lobato 1994). The modern vegetation surrounding the rock shelter is a combination of native forests, secondary scrub and man-made grasslands either assigned to crop or pasture (Catalan 1999). The primary forest is characterised by a blend of floristic elements from the lowland deciduous beech forests (Nothofagus obliqua), the northern sclerophyllous vegetation (matorral) and the Valdivian rainforest from southern Chile. At higher elevations Andean beech species (Nothofagus dombeyi, N. alpina and N. pumilio) and monkey-puzzle trees (Araucaria araucana) dominate the landscape. Secondary scrubs are formed by native and exotic shrubs, the native bamboo Chusquea quila, and several species of Myrtaceae, and, in pastures, numerous Poaceae.

Location of the Marifilo1 rock shelter in Chile
The Marifilo 1 rock shelter
Stratigraphic analysis showed an extended and repeated occupation of the Marifilo 1 rock shelter from the late Pleistocene (c. 10,190 b.p.) to the arrival of Europeans in the region (Mera and Garcia 2005). Seven strata were identified, four with a cultural origin interspersed with sterile layers representing significant volcanic events (Table 1). Some disturbance due to recent farming activities was observed in the superficial levels (0–10 cm). Six dates were obtained for the site using thermoluminescence (TL) and radiometric dating techniques. TL dating of a fragment of ceramic was used to date the superficial stratum. Four wood charcoal fragments (1 from layer 3, 1 from layer 4 each, and 2 from layer 6) and 1 fragment of human parietal bone (layer 4) were used to date the deeper levels using radiocarbon dating. Ten hearths of varying size and density were also uncovered, three in the Ceramic period, five in the mid-Holocene period and two in the early Holocene period. Animal remains were retrieved from the mid-Holocene, including bone debris of Pseudalopex griseus (Chilla fox), Pudu pudu (Chilean dwarf deer), mustelids, aquatic birds, and shells of freshwater mussels. Two instruments carved from the bones of Chilean dwarf deer were also found in this period (Velásquez and Adán 2004). Stone artefacts were uncommon in the site (Mera and Becerra 2001). A human burial was also discovered from the mid-Holocene associated with one bone and one stone artefact. A summary description of each strata, dates and main archaeological features is shown in Table 1.
Materials and methods
The rock shelter was excavated during three summer campaigns using a grid of 8 × 1 m2 excavation units. Reported here is the analysis of plant remains recovered from three excavation squares situated in the centre of the rock shelter, where most of the daily activities were expected to have taken place. Sediment was extracted from each quadrant in 10 cm deep layers following the natural stratigraphy of the site, where each cultural stratum was clearly delimited by a sterile layer of volcanic origin. Comprehensive retrieval of archaeological material was facilitated by use of wet sieving and flotation. Previous studies by Solari and Lehnebach (2004) and Solari et al. (2004) showed that a wider range of seed sizes may be recovered when combining these two techniques. Wet sieving allowed us to retrieve items of ≥3 mm in diameter, while flotation, including light and heavy fractions, recovered items over 0.5 mm in diameter. Wood charcoals and seeds were both present in the sediment preserved by charring. Wood charcoal and seeds recovered by wet sieving during excavation were stored till examination. A sub-sample equivalent to 12.5% of each level was collected and bagged to be floated after completion of excavation. A total of 43 sub-samples were floated (c. 272 l); 12 from the Ceramic (70 l), 21 from the mid-Holocene (133 l) and 10 from the early Holocene (69 l).
Seeds were studied under a dissecting stereomicroscope (Nikon SMZ645). Seed size, external features and internal anatomy were used for identification, assisted by comparison with modern plant specimens from the Laboratorio de Arqueobotánica, Instituto de Ciencias Sociales (Universidad Austral de Chile) and published identification keys (Martin and Barkley 1961; Mol 1983; Matthei 1995; Buxó 1997; Planella and Tagle 1998). Wood charcoal was identified through examination of transversal and longitudinal sections under a reflected light microscope (Olympus BX60) and comparison to modern specimens and published descriptions (Rancussi et al. 1987; Solari 1993; Wagemann 1949). Terminology followed standard practice (Adams 2004) and botanical nomenclature followed Marticorena and Quezada (1985).
Taxon ubiquity (Popper 1988) and the Shannon-Weaver Index (Pearsall 2000) were used to analyse the data. The Shannon–Weaver index or diversity index (DI) was applied only to the seed data and was calculated using the following formula:
Where N X is the number of seeds of taxon “X” in the phase; and N T is the total number of seeds in the phase. A diversity index close to 1 (the maximum) shows that a large number of taxa are evenly distributed, therefore diversity is high. A lower index, on the contrary (close to zero), results when the number of taxa present in the phase is low, or when one or a few taxa account for most of the population (Pearsall 2000).
Results
Seed remains
A total of 579 charred and uncharred seeds were retrieved from the three site phases (early Holocene, mid-Holocene and Ceramic). Nearly 95% of them were positively identified or assigned to a genus, family or division. The remaining 5% corresponded to four unknown seed types (8 specimens) and unidentifiable fragments (19 specimens). All uncharred seeds were excluded and considered modern in origin, being only used to infer the degree of disturbance and/or contamination of the period. Removal of the uncharred seeds left a total count of 472 charred seeds: Ceramic period n = 93; mid-Holocene period n = 153; early Holocene period n = 18; and 208 seeds accumulated in one mid-Holocene hearth. These macro-remains represent 19 plant taxa and one epiphytic fungus of the genus Rossellinia (Xylariaceae).
The abundance and ubiquity of species recovered in each period are given in Fig. 2. Seeds of the endemic evergreen tree Aextoxicum punctatum (Aextoxicaceae) were the most numerous (379 seeds) and ubiquitous types at the site (44% of the samples). This species was found in the three cultural periods but its abundance was higher in the recent periods: 52.4% in the mid-Holocene period and 66.7% in the Ceramic period (Fig. 2). A large accumulation of A. punctatum seeds (n = 208) was also recovered from one of the hearths in the mid-Holocene period on top of the child burial. These seeds were charred, with both their shape and internal anatomy incredibly well preserved (Fig. 3). The second most common seed in the site was assigned to the herbaceous family Convolvulaceae (14 seeds, 23% of the samples). These were present in all three periods but were more abundant in the Ceramic period (Fig. 2). The Poaceae family was also well represented; specimens of Bromus cf. mango/unioloides (16 caryopses), Poaceae 1 (7 caryopses) and Poaceae 2 (13 caryopses) were recovered from 11% of the samples. Seeds from several other tree and herbaceous taxa were poorly represented (<7%) and only appeared in particular contexts. For example Relbunium/Galium sp. was present only during the early Holocene and Pisum sativum during the Ceramic period.

Abundance and ubiquity of charred seeds and fungal remains found at Marifilo 1 separated by occupation phase. Seeds of Aextoxicum punctatum (n = 208) found in a hearth of the mid-Holocene period were not included in this table

SEM micrograph of charred Aextoxicum punctatum cotyledons found in Marifilo 1 (scale bar = 1 mm)
Composition of the plant assemblage varied between the three cultural periods. The early and the mid-Holocene period had the lowest diversity indices; DI 0.51 and 0.27, respectively. The Ceramic period, on the contrary, showed the highest diversity index (DI 0.97) indicating a more diverse plant assemblage (18 taxa) and a most even distribution of the remains within the period.
Wood charcoal analysis
About 700 fragments of wood charcoal were recovered but only c. 600 were successfully identified or classified. The small size of the remaining specimens (≤2 mm) made handling and identification difficult. Fragments were recovered from all three periods (Ceramic = 221; mid-Holocene = 207; early Holocene = 73) and from six hearths (n = 103 fragments). Hereafter, fragments scattered over the site will be referred to as dispersed charcoal and those recovered from hearths as accumulated charcoal. Dispersed charcoal contained 28 different native plants, including 26 dicotyledons, 1 monocotyledon and 1 gymnosperm. Tree and shrubby taxa such as Aristotelia chilensis (38.2%), Myrtaceae (35.3%), Peumus boldus (32.4%) and Aextoxicum punctatum (29.4%) were the most abundant at the site. These were followed by Proteaceae and Nothofagus deciduous type (26.5%), Laurelia (23.5%), cf. Persea lingue (20.6%) and one woody monocotyledon species (20.6%). The remaining taxa were poorly represented and their abundance scores were below 10%. Only five small shrubby taxa were identified; Berberis sp, Buddleja globosa, Colletia sp, Ericaceae and Ovidia pillo-pillo.
With only eight species and 73 fragments the early Holocene samples contained the lowest diversity and abundance of dispersed charcoal fragments. The mid-Holocene and the Ceramic periods contained much higher diversity with 22 and 20 plant taxa, respectively. During the early Holocene, the heliophylous tree P. lingue (66.7%) and the shrub A. chilensis (44.4%) dominated the assemblage (Fig. 4). Later, during the mid-Holocene, the sclerophyllus tree P. boldus (35.3%), a plant in the Myrtaceae family and one woody monocotyledon (29.4% each) and a Nothofagus species (23.5%) were the most abundant at the site. During the Ceramic period the plant assemblage was dominated by four tree taxa; Aextoxicum punctatum, Nothofagus deciduous, Proteaceae and A. chilensis. A considerable number of bark fragments and unidentifiable woody fragments were also recovered from the Ceramic phase.

Abundance and ubiquity of charcoal remains found at Marifilo 1 separated by occupation phase
Species composition and abundance for the 6 hearths studied are showed in Fig. 5. Taxon diversity and fragment abundance were low in the deeper hearths, with only five taxa recorded in Hearth 1, three taxa in Hearth 2 and two taxa in Hearth 3. All fragments, except those of Myrtaceae in Hearth 2, were assigned to tree or major shrubby taxa. Superficial hearths, on the contrary, contained a greater number of charcoal fragments and higher taxonomic diversity (Fig. 5). Remains of the native trees P. boldus, A. punctatum, Gevuina avellana, Laurelia and Proteaceae were the most abundant here. Fragments of minor shrubs (such as Berberis and Compositae) were also recovered from these hearths.

Percentage of taxa found in wood charcoal assemblages from hearths 1 to 6 at Marifilo 1
Discussion
Seed remains
Charred seeds recovered from the rock shelter represented a number of native trees and both wild and cultivated herbaceous plants. Ethnographic and ecological information was used to group the seeds into three categories based on the assumed source (Table 2): those collected as food; seeds introduced with food items (that is, only the flesh of the fruit or the tuber is consumed or utilised and the rest discarded); and seeds introduced with non-food items (branches bearing fruits used in fires), see Minnis (1981) and Pearsall (2000) for more on this classification.
Collected food seeds included pulses and grains. Five specimens of Pisum sativum (pea) were recovered from the superficial levels; however, their archaeological origin is uncertain since this legume is currently cultivated in the immediate surroundings of the site and may have been introduced by disturbance. Among the grains, Bromus mango/unioloides was the most abundant and, unlike P. sativum, it was also present in the earlier periods. Historical records indicate that Bromus mango and B. unioloides were collected, used to make bread and, at some stage, cultivated by the Mapuche people in the south of Chile (Reiche 1901; Aldunate and Villagrán 1992). The presence of charred caryopses of Poaceae types 1 and 2 in this group may have a similar origin. The number of edible grains in the south of Chile is very limited and the only other taxa mentioned in the historical records belong to the genus Chusquea (Chilean Bamboo). If the caryopses of Poaceae types 1 and 2 belong to Chusquea species, collection of grains from the wild could be supported. The historical record reports that these grains were collected and used to make bread by the pre-Hispanic populations in the south of Chile (Aldunate and Villagrán 1992). Unfortunately, plant material or detailed taxonomic descriptions of Chusquea caryopses were not available for comparison and this idea remains speculative.
A more diverse range of seeds have been classified as those introduced to the site with food items. This category includes specimens of the herbaceous family Convolvulaceae and shrubby/tree species such as Aristotelia chilensis, Gevuina avellana, Podocarpaceae and Peumus boldus. Seeds of Convolvulaceae were the second most abundant in the site and were present in all three cultural periods. Despite the low number of species in this family in Chile, identification of the remains to the genus or species level was impossible due to the great morphological convergence in seed morphology. In southern Chile, however, only species of Ipomoea and Calystegia are collected for food (tuberous stem) or medicinal purpose (leaves), respectively (Reiche 1901; Aldunate and Villagrán 1992). It is likely that plants were collected over summer when fruiting, and the aerial and fruiting parts were discarded and burned. The shrub Aristotelia chilensis produces an edible black-purplish berry during summer. Only the flesh of these berries is consumed and the seeds are usually discarded. Nowadays, and also historically, they are collected by the local communities either for immediate consumption, production of alcoholic beverages or to be dried and stored for consumption over winter (Reiche 1901; Aldunate and Villagrán 1992). Conversely, in Gevuina avellana it is the cotyledons inside the nutshell that are edible. The trees fruit during the summer and early autumn. The nuts are boiled and split open to eat the cotyledons which can also be roasted or they may also be stored and consumed over winter. The next species in this group belongs to the Podocarpaceae family, which has five species within three genera in Chile (Podocarpus, Prumnopytis and Saxegothaea; see Hoffmann 1998). Only the aril (fleshy structure covering the woody seed) of Prumnopytis andina is edible and it is locally known as the “uva de la cordillera” (Andean grape). Peumus boldus, the last tree species in this category, produces a small fleshy and juicy, yellow-greenish drupe that is available during summer (Hoffman 1998); only the flesh is eaten and the stone is discarded. These stones are rich in oils and burn very easily (personal observation).
The group of seeds introduced with non-food items includes native herbaceous and tree species of no evident use in the ethnobotanical or historical record. Their abundance is low, only one or two fragments. The native tree Aextoxicum punctatum is a remarkable exception within this group. Large accumulations of stones of this species were found in all three cultural periods. In fact, over 70% of the charred seeds recovered in the site belong to A. punctatum. A large accumulation of these seeds was also found in one hearth of the mid-Holocene period associated with a human burial (n = 208, 100%). The great abundance and continuous presence of this species in the site was unexpected. These drupes are not consumed by people and the only uses reported for this species in the literature are carpentry, charcoal and firewood. Aextoxicum punctatum is common in wet areas of southern of Chile and some scattered individuals are currently found near the site. Female individuals form a purple drupe that ripens during autumn; this is either dispersed by gravity, birds or mice. Several possible explanations account for the great abundance of these seeds in the rock shelter. Dispersal by birds and gravity could explain the accumulation of A. punctatum in the site, but not their repeated carbonization. Accumulation by rodents was also discarded because mice usually eat this fruit in situ (Murua 1995) and, again, that vector does not explain their carbonization. Pino et al. (2004) suggested these fruits were probably used to rekindle the fire; however, laboratory observations indicate they have poor combustibility (Solari et al. 2004). Other possible explanations are the use of these fruits to cover fire embers or the use of fruiting branches as firewood. If the accumulation of this species has cultural origin, its presence in the site could be a good indicator of seasonal activities and/or occupation of the site during the summer-autumn period when it is fruiting.
Finally, it is important to point out the absence or little representation of species of dietary importance such as Araucaria araucana (pehuen, monkey puzzle), Chenopodium quinoa (quinoa), Gevuina avellana (Chilean hazelnut), Ugni molinae (murta, Chilean guava) and Solanum species (potatoes). Collection of these taxa has been well documented in ethnobotanical and historical records (Reiche 1901; Aldunate and Villagrán 1992) and many of them are commonly found in the vicinity of the shelter. It is possible that the processes involved in their preparation before consumption or the way they were consumed may have hindered their preservation.
Wood charcoal
Wood charcoal, unlike charred seeds, provided a greater diversity of taxa (28 taxa) including dicotyledons, monocotyledons and a gymnosperm. None of the identified taxa have a specific use according to the ethnographic literature and using direct analogy were probably used for a variety of functions, including firewood, charcoal, construction and cooking tools (Reiche 1901; Aldunate and Villagrán 1992; Hoffmann 1998). Such opportunistic use of wood has been observed in Patagonian settlements by Solari (1993). For example, her analyses of wood charcoal from hearths have showed the use of Drimys winteri as firewood in these sites, despite the astringent smoke it produces when burning. This tree is primarily used in rituals or collected due to its medicinal properties (Aldunate and Villagrán 1992).
Many of the tree or shrubby taxa recovered in the rock shelter did not appear in the seed record suggesting these two disciplines complement each other in providing information about plant use (see discussion in Solari et al. 2004). For instance, Berberis sp. and Myrtaceae were found only as wood charcoal fragments. These two taxa produce edible berries during summer and early autumn which are collected and immediately consumed (Reiche 1901; Aldunate and Villagrán 1992). Complementarity between these two records is also demonstrated when comparing the number of seed remains of Aristotelia chilensis (one fragment) with the numerous charcoal fragments recovered (over 30). These results have indirectly confirmed the presence of many edible plants in the area, which did not appear in the seed record but may nevertheless have been used.
Analysis of dispersed charcoal fragments also allowed reconstruction of the vegetation surrounding the rock shelter during its past occupation phases and, to a limited extent, vegetation succession after disturbance episodes such as volcanic eruptions. Many of the taxa found in the rock shelter are elements of at least three forest types; the evergreen forest (Nothofagus, Laurelia and Aextoxicum), the deciduous forest (Persea) and the sclerophyllous forests (Peumus). This mixture of taxa is commonly found in ecotonal forests and is still observed in isolated patches close to the site. Fragments of a monocotyledon (5.9%) found in all three cultural periods may come from the genus Chusquea. The Chilean bamboo is the only woody monocotyledon occurring in these forests and generally colonises large areas after disturbance episodes. Pollen studies by Heusser (1984) also support the uninterrupted occurrence of the Chilean bamboo in this area. Other early successional shrubs found in the site were Aristotelia chilensis, Colletya sp., Ericaceae, Berberis sp. and Proteaceae. In general an increase in evergreen and late successional taxa along with a rise in species diversity was observed from the mid-Holocene to Ceramic periods. The tree assemblage during the early and mid-Holocene period is dominated by sclerophyllous and heliophyllous taxa such as Persea lingue and Proteaceae. Similar changes in the vegetation assemblage have been evidenced by pollen records obtained in the area (Heusser 1984).
Unlike dispersed charcoal fragments, remains concentrated in hearths could be directly associated to human activities in the site. The hearth sequence followed the same trend of species diversity observed in the dispersed charcoal samples, with increasing diversity in the later periods. Hearths from the early Holocene period generally had fewer taxa than the later periods (Fig. 5). This may indicate sporadic and brief occupation of the rock shelter during the early Holocene, as suggested previously by Mera and Garcia (2005); however, this trend may be also caused by differential preservation conditions or loss of remains from the older deposits (see below).
Fragments recovered from the hearths were mainly from trees and only one minor shrub (Berberis sp.) was found in them. This shrub is usually found in open and disturbed areas. It is interesting to note the presence of charcoal fragments of Asteraceae in Hearth 2 (Fig. 5b). There are only two woody Asteraceae genera in this area that may withstand a fire and produced charcoal remains, namely Dasyphilum diacanthoides (a tree) and a range of Baccharis species (shrubs). If these remains belong to Baccharis species, wood collection from open and disturbed areas away from the forest could be further supported as these species are rarely found growing under the forest canopy.
Identification of charcoal fragments was not always simple, many fragments were small, fragile and their anatomy was distorted. This situation was unusual and has never been observed in earlier charcoal studies in southern Chile and Patagonia (Solari 1991, 1993; Gaete et al. 2004). A possible explanation is the effect successive volcanic events may have on the fragment anatomy producing intrusions or crystallisation.
Diversity of the plant assemblage
The Ceramic period contained a more diverse plant assemblage (DI 0.97) than the mid-Holocene (DI 0.27) and early Holocene (DI 0.51). These differences could be attributable to habitation patterns, natural processes and, to a limited extent, sampling effort. The scarcity of remains in the earlier levels may be correlated with short occupation periods of the shelter or occupation by small groups, or both. This has been previously suggested for this site based on the limited number of ecofacts and biological remains recovered there (Mera and Garcia 2005). Seasonal occupation of the shelter, and therefore mobility of these groups, may be a plausible explanation. This could be further supported if we confirm that the presence of A. punctatum seeds has seasonal implications and cultural origin.
Seed durability and preservation conditions could also have affected the diversity of the plant assemblage (Wright 2003). Seed and wood remains from different species may differentially withstand the effects of fire and weathering. These factors are particularly important in southern Chile where preservation of botanical remains is only possible if carbonisation has occurred. For instance, preservation of A. punctatum seeds was probably favoured by the hard endocarp and its low combustibility. Conversely, most of the edible plants in these forests have fleshy fruits and small seeds that may not withstand carbonization and, consequently, be undetectable in the archaeological record. Unfortunately, taphonomic studies of plant remains in Chile are lacking and we are unable to predict the durability of plant remains or successfully identify the variables affecting their presence or absence in the site. The effect of volcanic events on the presence of biological remains is also worth considering here. Volcanism may not only alter the preservation of biological remains but also the vegetation in the surrounding area and, consequently, may reduce or modify the resources available in the area.
The effect sampling effort may have on the number of taxa recovered in each period should also be acknowledged, but this may only explain the low diversity index observed in the early Holocene period (DI 0.51). The volume and number of samples examined for this phase was the smallest (69 l, 10 samples) and it is likely that a larger sample size could have represented the botanical assemblage better. However, it should be noticed that these numbers are not that different from the Ceramic period (70 l, 12 samples), which showed the highest diversity index (DI 0.91). The low DI obtained in the mid-Holocene period however cannot be attributed to a limited sampling effort. This period had the lowest diversity index (DI = 0.27) but the largest volume of sediment (133 l) and number of samples (21 samples) examined of all three periods. Here the low DI was probably caused by the great abundance of A. punctatum seeds (c. 90%).
Conclusions
Macro-botanical remains found in the rock shelter represent a variety of native trees, shrubs and herbs usually found in the temperate forests of southern Chile. The seed record demonstrated the use of native grains and, to some extent, the collection of fruits and nuts available in these forests over summer and autumn. Furthermore, the analysis of dispersed charcoal fragments and those accumulated in the hearths were especially useful to identify plant resources available in the area and uncover other taxa that did not appear in the seed record. They were also useful to infer successional events and describe the vegetation surrounding the rock shelter during its occupation. Collection of resources from other habitats and, to some extent, the seasonal habitation of the shelter during the early and mid-Holocene period could also be suggested.
The study of the plant remains from this site has proved to be informative and provided the first insights into the use and collection of plant products in this region of Chile; however, further archaeobotanical evidence is still required to fully understand the subsistence strategies that evolved in these ecosystems. A new methodological approach to detect a greater range of botanical evidence will be indispensable. The study of starch grains and phytoliths or other remains in vessels and grinding stones will help to broaden the understanding of plant use. A range of taphonomic studies will be also required to understand plant remain durability and the variables affecting their preservation.
References
Adán LI, Mera R, Becerra M, Godoy M (2004) Ocupación arcaica en territorios boscosos y lacustres de la región precordillerana andina (IX & X regiones): el sitio “Marifilo 1” de la localidad de Pucura. Chungará 36:1121–1136
Adams KR (2004) Archaeobotanical analysis: principles and methods. URL: http://www.crowcanyon.org/plantmethods. Version 4 August 2006
Aldunate C, Villagrán C (1992) Botánica indígena de Chile, Museo Chileno de Arte Precolombino. Editorial Andres Bello, Santiago
Asouti E (2003) Woodland vegetation and fuel exploitation at the prehistoric campsite of Pinarbasi, south-central Anatolia, Turkey: the evidence from the wood charcoal macro-remains. J Archaeol Sci 30:1185–1201
Bonzani RM (1997) Plant diversity in the archaeological record: a means toward defining hunter-gatherer mobility strategies. J Archaeol Sci 24:1129–1139
Bragg K (1981) La etnobotánica y ecología humana de una comunidad indígena de Chile. The Thomas J. Watson Foundation, Rhode Island
Buxó R (1997) Arqueología de las plantas. Editorial Crítica, Barcelona
Catalán R (1999) Caracterización de la vegetación de la cuenca del lago Calafquén y sus potencialidades para la ocupación humana”. Informe Proyecto Fondecyt 1970105
Dillehay T, Rossen J (1997) Integrity and distribution of archaeobotanical collection. In: Dillehey T (eds) Monteverde: a late Pleistocene settlement in Chile: the archaeological context and interpretation, Vol. 2. Smithsonian Institution Press, Washington DC, pp 351–381
Gaete N, Navarro X, Constantinescu F, Mera C, Selles D, Solari ME, Vargas ML, Oliva D, Durán L (2004) Una mirada al modo de vida canoero del mar interior de Piedra Azul. Chungará 36:333–346
González H, Valenzuela R (1979) Recolección y consumo del piñon. Actas del VIII Congreso de Arqueología Chilena. Ediciones El Kultrun, Santiago
Heusser CJ (1984) Late-Glacial-Holocene climate of the Lake District of Chile. Quat Res 22:77–90
Hoffmann A (1998) Flora silvestre de Chile; zona araucana. Ediciones Fundación Claudio Gay, Santiago
Kenward H, Hall A (1997) Enhancing bioarchaeological interpretation using indicator groups: stable manure as a paradigm. J Archaeol Sci 24:663–673
Marticorena C, Quezada M (1985) Catálogo de la flora vascular de Chile. Gayana Botánica 42:1–157
Martin AC, Barkley WD (1961) Seed identification manual. University of California Press, Los Angeles
Matthei O (1995) Manual de las malezas que crecen en Chile. Alfabeta Impresores, Concepción
Mera C, García C (2005) Alero Marifilo-1. Ocupación holoceno temprana en la costa del lago Calafquén (X Región, Chile). In: Civalero MT, Fernández PM, Guraieb AG (eds) Contra viento y marea, Arqueología de la Patagonia, Instituto Nacional de Antropología y Pensamiento Latinoamericano. Sociedad Argentina de Antropología, Buenos Aires, pp 249–262
Mera C, Becerra R (2001) Análisis del material lítico de los sitios de la costa del Calafquén Museos 25:7–12
Minnis PE (1981) Seeds in archaeological sites: sources and some interpretative problems. Am Antiquity 46:143–152
Mol E (1983) Manual para la identificación de disemínulos de plantas herbáceas no cultivadas del Jardín Botánico de la Universidad Austral de Chile, Bachellor Thesis, Universidad Austral de Chile, Valdivia
Murua R (1995) Comunidades de mamíferos del bosque templado de Chile. In: Armesto JJ, Villagrán C, Arroyo MK (eds) Ecología de los bosques nativos de Chile. Editorial Universitaria, Santiago, pp 113–133
Pearsall DM (2000) Paleoethnobotany: a handbook of procedures, 2nd edn. Academic, San Diego
Petit-Brehuil M, Lobato J (1994) Análisis comparativo de la cronologia eruptiva histórica de los volcanes Villarrica y Llaima (38–39 S). Actas VII Congreso Geologico Chileno 1:366–370
Pino M, Adán LI, Seguel O, Mera R, Brummer R (2004) Entre lagos, bosques y volcanes: geoarqueología del Valle Marifilo. distrito de Los Lagos Araucanos, Valdivia
Planella MT, Tagle B (1998) El sitio agro-alfarero temprano de la Granja: un aporte desde la perspectiva arqueobotánica. Publicación Ocasional del Museo Nacional de Historia Natural de Chile N°52, Santiago
Popper VS (1988) Selecting quantitative measurements in paleoethnobotany. In: Hastorf CA, Popper VS (eds) Current paleoethnobotany. University of Chicago Press, Chicago, pp 53–71
Quiroz L (2001) Arqueobotánica del sitio intermedio tardío parcelación El Ingenio (Viña Errázuriz, V Región): desafío metodológico en un sitio arado. Bol Soc Chil de Arqueol 32:57–65
Quiroz L, Belmar C (1998) Evidencias de restos vegetales en el contexto arqueológico de Chan-Chan 18 (Valdivia): su implicancia interpretativa en la economía de grupos cazadores recolectores. IV Jornadas de Arqueología de la Patagonia, Río Gallegos
Ramirez C (1989) Macro-botanical remains. In: Dillhey T (ed) Monteverde: a late Pleistocene settlement in Chile: palaeoenvironment and site context, Vol 1. Smithsonian Institution Press, Washington DC, pp 147–170
Rancussi M, Nishida M, Nishida H (1987) Xylotomy of important Chilean woods. In: Nishida M (ed) Contributions to the botany in the Andes II. Academia Scientific Book, Tokyo, pp 68–158
Rapaport E, Ladio A (1999) Los bosques andino-patagónicos como fuentes de alimento. Bosque 20:55–64
Reiche K (1901) Los productos vejetales indíjenas de Chile. Imprenta Cervantes, Santiago
Rodriguez MF (2000) Woody plant species used during the Archaic Period in the Southern Argentine Puna. Archaeobotany of Quebrada Seca 3. J Archaeol Sci 27:341–361
Rodriguez MF (2004) Woody plant resources in the southern Argentine Puna: Punta de la Peña 9 archaeological site. J Archaeol Sci 31:1361–1372
Rossen J, Dillehay T (1997) Modelling ancient procurement and use at Monte Verde. In: Dillehay T (ed) Monte Verde: a late Pleistocene settlement in Chile: the archaeological context and interpretation, Vol 2. Smithsonian Institution Press, Washington DC, pp 331–350
Smith-Ramírez C (1995) Algunos usos indígenas-tradicionales de la flora del bosque templado. In: Armesto JJ, Villagrán C, Arroyo MK (eds) Ecologia de los bosques nativos de Chile. Editorial Universitaria, Santiago, pp 389–404
Solari ME (1991) Estudio antracológico del sitio Punta Baja 1 (Mar de Otway). Anales del Instituto de la Patagania. Series Ciencias Sociales y Humanidades 19:115–120
Solari ME (1993) L’homme et le bois en Patagonie et Terre de Feu au cours des six derniers millénaires: recherches anthracologiques au Chili et en Argentine, doctoral thesis, Université de Montpellier II, Montpellier
Solari ME (1994) Estudio antracológico del archipiélago del Cabo de Hornos y Seno Grandi. Anales del Instituto de la Patagonia. Series Ciencias Sociales y Humanidades 22:137–148
Solari ME (2003) Analisis antracológico del Locus 1. Magallania 31:327–335
Solari ME, Bernard V, Legupil D, Richard H, Roiron P, Schoenauer P (2002) Palaeo-environmental approach of Ponsonby archaeological site (Riesco Island, Chilean Patagonia). BAR Internat Ser 1063:179–185
Solari ME, Lehnebach CA (2004) Pensando la antracología para el centro-sur de Chile: sitios arqueológicos y bosque en el Lago Calafquén. Chungará 36:373–380
Solari ME, Lehnebach CA, Adán LI (2004) Metodologías compartidas para el estudio de carbones y semillas de los Períodos Arcaico y Formativo del alero “Marifilo 1” (Lago Calafquen, 39°S). Boletin del Museo Nacional de Historia Natural 53:125–137
Thomasson K. (1963) Araucarian lakes. Acta Phytogeographica Sve 47:1–139
Velásquez H, Adán LI (2004) Evidencias arqueofaunísticas del sitio alero Marifilo-1: Adaptación a los bosques templados de los sistemas lacustres cordilleranos del centro sur de Chile. Boletín de la Sociedad Chilena de Arqueología 33/34:27–35
Villagrán C (1998) Etnobotánica indígena de los bosques templados de Chile: sistema de clasificación de un recurso multiple. Revista Chilena de Historia Natural 71:245–268
Wagemann W (1949) Maderas chilenas: contribución a su anatomía e identificación. Lilloa 16:263–375
Weiss E, Kislev ME (2004) Plant remains as indicators for economic activity: a case study from Iron Age Ashkelon. J Archaeol Sci 13:1–13
Wright P (2003) Preservation or destruction of plant remains by carbonization. J Archaeol Sci 30:577–583
Acknowledgments
The authors want to thank: the Marifilo Family and Pucura community for their friendship and help during this study; F. Poblete for help in the laboratory; the archaeologists who worked in this site, and the two anonymous reviewers for helpful comments on the manuscript. This study was funded by the grants FONDECYT 1040326 and 1010200, and DID-UACH 200154.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by A. Fairbairn.
Rights and permissions
About this article
Cite this article
Lehnebach, C.A., Solari, M.E., Adán, L.I. et al. Plant macro-remains from a rock shelter in the temperate forests of southern Chile. Veget Hist Archaeobot 17, 403–413 (2008). https://doi.org/10.1007/s00334-007-0123-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-007-0123-9