
Abstract
A 90 m deep core from the Massaciuccoli lake basin (Tuscany, Italy) offers new material for improving the knowledge of the Holocene vegetation history in the coastal plains in Tuscany and Liguria bordering the Mar Ligure (Ligurian sea). In this paper, an assessment of the upper part of the core is presented. During the Holocene, the area was covered by woodlands whose composition changed in time. Numerous oscillations of the tree taxa were recorded which can probably be connected to sea level variations that had a strong influence on the depth of the water table. Human impact is evident particularly at ca. 4200–2500 b.p. when traces of opening of the woodland accompanied by noticeable pollen percentages of Vitis could suggest an early agricultural practice favouring this native plant. The new data have been integrated with previous palaeobotanical and archaeobotanical research. The occurrence of coastal wetlands has been shown. Throughout the Holocene, Mediterranean evergreen vegetation is generally poorly represented; it becomes widespread only in the late Holocene, suggesting that its establishment was probably favoured by the management practices in the coastal plains, particularly land reclamations, starting from the Roman period.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
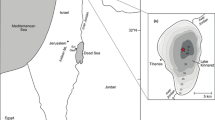
The coastal plains of Liguria and northern Tuscany, facing the Mar Ligure (Fig. 1), have been the setting of several corings and archaeological excavations, but their vegetation history is still far from being well known. Previous palynological investigations carried out in the Ligurian and Tuscan coastal areas (Ferrarini 1969; Arobba et al. 2001, 2004; Guido et al. 2004a,b), in the lower Arno river valley (Galletti Fancelli 1971, 1979; Paoli and Cellai Ciuffi 1980, 1983; Abbate Edlmann et al. 1991, 1993; Mori Secci 1991; Begliomini et al. 2003; Mariotti Lippi et al. 2006) and also in the Massaciuccoli basin (Marchetti 1935; Marchetti and Tongiorgi 1936; Tongiorgi 1936; Grassi et al. 2000; Menozzi et al. 2003) have supplied much information regarding vegetational and climatic changes in this area during the Quaternary.

Geographical location of the Massaciuccoli basin and the other sites mentioned in the paper
The Massaciuccoli lake basin (northwest Tuscany, Italy) has been considered for a long time the principal biostratigraphical archive for the study of the landscape evolution of that area. However, the complex history of the basin reduces the availability of suitable study sites. In fact, the area has been inhabited at least since the Middle and Upper Palaeo-lithic period (Cremonesi Grifoni and Tozzi 1994), subjected to repeated episodes of drainage starting from Roman times, and has also been influenced by quarrying activities. The silica sand extractions ceased only recently, and strongly affected the stratigraphy of the quarried areas (Mazzanti and Nencini 1994). This situation has limited the possibility of investigating the palaeoenvironment, since the stratigraphic sequences are damaged and the chronological sequence altered. For this reason, pollen analyses in the area must be preceded by historical study of the sampling site and geostratigraphic investigations of the sediments.
A 90 m deep core (MSC) was obtained by ENEA (Ente per le Nuove tecnologie, l’Energia e l’Ambiente –Italian National Agency for New Technologies, Energy and the Environment) in an area of the basin not directly disturbed by human activity. The sedimentology and chronology of this core have been presented in previous reports (Antonioli et al. 2000; Lambeck et al. 2004).
This paper is devoted to the Holocene vegetation history of the site as shown from pollen analyses of the upper portion of the MSC core. The study of the lower part of the core, which reaches a depth of 90 m with a date of 130,000 b.p. at ca. 70 m (Antonioli et al. 2000), is still in progress.
The study area
The Massaciuccoli lake basin is situated on the coastal plain between the mouths of the rivers Arno and Magra, at sea level (Fig. 1). It is characterised by complex dune systems along the sandy coasts, and by numerous inland wetlands, today much reduced by repeated drainage works for agricultural land use. The area, which was formed in the Upper Pleistocene, is the result of the interaction of geological and palaeogeographical factors, mainly the varying sediment charge of the Arno-Serchio fluvial system, which created the Versilia plain. The territory has been affected by numerous marine ingressions, and has passed through lagoon and fresh water marsh phases (Federici 1993).
The climate is essentially Mediterranean, influenced by the proximity of the sea which moderates the temperatures. In the Arno floodplain near Pisa, 900 mm mean annual precipitation and 15 °C mean annual temperature have been recorded. Winds, principally blowing from the south-west, are also an important factor, especially concerning relative humidity values that never fall below 60% (Rapetti and Vittorini 1994).
The current flora and vegetation of this plain have been the subject of numerous studies (Corti 1955; Montelucci 1964; Ferrarini 1977; Gellini et al. 1986; Tomei et al. 1986, 1995; Arrigoni 1990; Vagge and Biondi 1999; Garbari 2000; Garbari and Cecchi 2000). The coastal sandy zone is characterised by pioneer halophytes and psammophytes (plants growing on sand) such as Cakile maritima and Ammophila littoralis. In the wetland areas Phragmites australis, Cladium mariscus, Typha angustifolia and Carex elata are the most common species. Proceeding inland, Juniperus oxycedrus subsp. macrocarpa, and J. communis appear on the dunes, followed by Quercus ilex woods. In the interdunal areas the vegetation is formed by Quercus robur and Carpinus betulus. Where the soil becomes damper, Fraxinus oxycarpa grows, or alternatively Populus alba together with Alnus glutinosa and Ulmus minor.
All of the floristic and vegetational investigations carried out in the Massaciuccoli lake basin indicate the high bio-ecological significance of the vegetation which is the result of past distributional and climatic factors. The area includes the last conservation niches for many plant species that were more generally widespread in the past and have almost completely disappeared from Italy today. For this reason, part of this diversified natural heritage has recently (1979, 1995) been included in the “Parco Regionale di Migliarino-San Rossore-Massaciuccoli”.
Materials and methods
The core was obtained in the Massaciuccoli basin (43 °48’N, 10 °19’E) by ENEA in 1998; the subsampling was carried out by Prof. Bonadonna’s team at the Dipartimento di Scienze della Terra of the Università di Pisa (Table 1, Fig. 2).

Schematic representation of stratigraphy, chronological references, and pollen concentrations (APF) in the MSC core
In a previous research programme, pollen analyses were performed along all of the 90 m MSC core, with the aim of obtaining a general survey (Menozzi et al. 2003). In this paper, attention has been devoted to the upper 34 m of the core (Fig. 2), previously named MaSE II pro parte and MaSE-III, which correspond to the last 10,000 years up to Roman times (Table 2). The samples were collected, where possible, every 10 cm, with a total of 67 samples. The discontinuities were caused by the nature of the sediments which were sometimes sandy or gravelly (Table 1, Fig. 2).
Each sample was treated according to the standard methodologies (Fægri and Iversen 1989). Pollen analysis was carried out with a light microscope, operating at 400–1000×. The grains were identified making reference to the literature (Punt et al. 1976–1996; Fægri and Iversen 1989; Andersen 1979; Moore et al. 1991; Reille 1992–1998) and to reference collections. Pollen concentration (APF = Absolute Pollen Frequency) was calculated according to Accorsi and Rodolfi (1975); the values of all the examined samples are given in Fig. 2. Pollen percentages are calculated from the total land plant pollen sum (TLP). Only the samples with at least 250 identified grains are presented in the percentage diagram (Fig. 3), which was drawn using GpalWin (Goeury 1997).

Pollen diagram of the Holocene MSC core (selected taxa)
In the diagram, hygrophilous and hydrophilous herbs include Juncaceae, Filipendula cf. ulmaria, Anagallis cf. tenella, Lythrum, Glyceria type, Sparganium, Typha latifolia, Typha angustifolia type, Alisma, Butomus, Callitriche, Hydrocotyle, Myriophyllum and Potamogeton.
Results
In the Holocene sequence, the APF values are variable, but generally low, ranging from approximately 100 to nearly 400,000 grains/g (Fig. 2). The state of preservation of the pollen grains was not always good. However, in the samples described below, the percentage of the unidentifiable grains is quite low, averaging around 3%.
The pollen diagram (Fig. 3) has been divided into seven pollen zones (PAZ), which will be discussed from the earliest to the most recent.
MSC1 (33.85–33.45 m; 9590 ± 60 b.p. at 34 m)–This sequence is characterized by significant percentages of AP that are mainly represented by deciduous Quercus, Corylus and Alnus. Other elements of the mixed oak woodland, like Ulmus and Tilia, are noteworthy, although they are present in low percentages. The conifers, mostly Abies and Pinus, are scarcely represented. Q. ilex shows a slight increase, always with low values (<10%). Poaceae and Cyperaceae, the first with higher percentages, form the main part of the NAP component and follow a decreasing trend. Fern spores are recorded with low values.
MSC2 (33.05–32.20 m; 8970 ± 50 b.p. at 32.10 m) The onset of the second PAZ is marked by increasing values of Abies. Quercus is present at lower percentages than in MSC1; Corylus and Alnus decrease in the upper part of the zone. NAP percentages are always very low and represented by Poaceae and Cyperaceae. Fern spore percentages become more significant.
MSC3 (31.98–29.52 m; 8419 ± 48 b.p. at 29.52 m) The zone formed by sterile sandy levels (Table 1) is characterized by oscillating AP percentages. Abies follows an overall decreasing trend. Deciduous Quercus values are low, as are other elements of the mixed oak woodland; Alnus percentages oscillate. There are high values of Poaceae and a peak of fern spores in the lowest sample, and these then maintain relatively high values.
MSC4 (29.20–27.34 m) In the overlying sandy silts, the analysed samples show very low concentration values. Only one sample (28.60 m) has a sufficient APF and has significant values of Abies pollen and, to a lesser degree, deciduous Quercus.
MSC5 (8.30–4.60 m; 5471 ± 41 b.p. at 7.50 m) The AP percentages decrease at first, then increase. In the lower part (subzone a: 8.30–5.40 m), Abies, Pinus, Quercus, and Alnus are important. Pinus and Q. ilex type show an opposite trend to Alnus. NAP, mainly Poaceae, decreases after a slight increase. Wetland and freshwater plants (such as Cyperaceae and Potamogeton) are also recorded. Fern spores show a peak in the middle of this subzone.
The upper part (subzone b: 5.20–4.60 m) is characterized by high percentages of Corylus and Alnus, while all other tree taxa drop to very low values. Abies disappears in the uppermost level.
MSC6 (2.60–1.80 m; 3827 ± 40 b.p. at 2.60 m to 2716 ± 37 b.p. at 2.10 m) Throughout the zone, AP values are formed almost only by Alnus, Salix and Vitis, the latter with two peaks of high percentages. The occurrence of Castanea is noteworthy.
MSC7 (1.45 m–39 cm; 2206 ± 52 b.p. at 1 m) The first sample shows the lowest AP value of the whole diagram and a peak of Chenopodiaceae and Asteraceae. The following samples have increased AP due to the contribution of Q. ilex type and Erica. Elements of the deciduous woodlands seem to be more or less established. At 42 cm, hygrophilous and aquatic plants show a peak, while Q. ilex type and Pinus decrease.
Discussion
The oldest samples (MSC1; Fig. 3), representing the beginning of the Holocene, show a tree cover mainly of deciduous Quercus, Corylus, Tilia and Ulmus. Alnus is also well re-presented, while conifers, particularly Abies, are scarce. The Poaceae values show the contemporary occurrence of open areas.
Earlier studies (Marchetti 1935; Marchetti and Tongiorgi 1936; Tongiorgi 1936; Alessio et al. 1965; Menozzi et al. 2003) suggest that a cold continental climate, which determined the development of woodlands dominated by Abies and Pinus sylvestris/mugo was established in the Late Pleistocene. Significant percentages of Abies and Pinus mugo/sylvestris pollen were still found at the onset of the Holocene in Pisa (Trinci 2003). However, mesophilous and Mediterranean taxa also occurred in the area, showing the existence of at least some temperate climate habitats, which might have played the role of refuge areas. The finding of Menyanthes trifoliata seeds (Marchetti and Tongiorgi 1936) shows the presence of wetlands, also indicated by freshwater plant pollen at the base of the Holocene (Trinci 2003).
Around 9000 b.p., the MSC core (MSC2) has high values of Abies pollen, suggesting the spreading of this conifer when the oak woodland was reduced and the open areas mostly disappeared. The subsequent oscillations of the AP values (MSC3) denote a rather variable landscape in which the tree cover alternately expanded and retreated, each time modifying its composition. This suggests a period of environmental instability, probably related to fluctuations of the water table. This is above all indicated by the oscillations of Alnus and Poaceae. A peak of both Abies and deciduous Quercus (MSC4) precedes the occurrence of a thick sand layer caused by a marine ingression (Figs. 2 and 3) in the Massaciuccoli basin.
At higher altitudes, the presence of significant percentages (>10%) of Abies are reported, starting from 11,500–10,500 b.p. in the northern Apennines (Terhürne-Berson et al. 2004). Particularly in the Ligurian Apennines, Abies is recorded from 10,600 b.p. together with Quercus robur type and Ulmus (Branch 2004). Starting from 10,000 b.p., the presence of significant percentages of both Abies and deciduous trees like Quercus, Corylus, Ulmus and Tilia were also recorded in the Appennino Parmense (Prato Spilla, Watson et al. 1994), and in the Tuscan-Aemilian Appennines (Lago Padule, Lowe et al. 1994a). In the Ligurian woodlands, Abies became the dominant element between 8600 and 6700 b.p. (Branch 2004); in the Tuscan-Aemilian Appennines, Abies shows high values after 9000 b.p., together with a drop of the Quercus-Tilia-Ulmus curve (Lago del Greppo and Lago Baccioli, Bertolani Marchetti et al. 1994).
In central Italy around 10,000–8000 b.p., Abies is sporadically recorded with low percentages, while Quercus and other broadleaved trees are dominant in the spectra (Lago di Vico, Magri and Sadori 1999; Stracciacappa, Giardini 2006). In the same period in southern Italy, noticeable percentages of Abies are recorded, even if broadleaved trees were prevalent (Lago Grande di Monticchio, Allen et al. 2002; Gulf of Salerno, Russo Ermolli and di Pasquale 2002). However, interpreting the relationship between the northern and southern Italian mountain plants is a difficult task, because of the different and peculiar vegetation histories of the two geographical areas.
Between about 7500 and 5500 b.p. the MSC pollen sequence is interrupted by a thick sand layer from a marine ingression that took place between 10,400 and 6600 b.p. (Figs. 2 and 3; Antonioli et al. 2000; Table 1). This gap in the vegetation history is partially filled by Ligurian coastal data (Cornara et al. 1996; Montanari et al. 1998; Brandolini et al. 2002), from cores of continental sediments in the city of Genova (Liguria), at the mouth of the streams Bisagno and Polcevera (Fig. 1). These analyses show that between 8500 and 5500 b.p., the coastal vegetation was characterised by woodlands in which Abies was abundant, together with Pinus, deciduous Quercus, Ulmus, Tilia, Alnus, etc. A substantially similar scenario results from the analysis carried out at the mouth of the river Centa (Fig. 1; Arobba et al. 2001). In France, Abies spread between 7200 and 5200 b.p. in the western Alps and Provence (Digerfeldt et al. 1997). A study in the Alpes Maritimes in southeast France places the onset of significant percentages of Abies at the beginning of the Atlantic period (Nichol-Pichard and Dubar 1998).
Starting from 5500 b.p. (MSC5), the Massaciuccoli sequence indicates a landscape composed of coniferous and broadleaved woodlands, with occasional open areas. Later, high percentages of Corylus could indicate the occurrence of woodland clearance, presumably a signal of human activity. More inland areas had a thin tree cover, dominated by deciduous Quercus, Ulmus, Fraxinus and Acer, as is shown by the archaeobotanical investigations at Pontedera dated to the Neolithic period (Fig. 1; Abbate Edlmann et al. 1991, 1993) and by analyses carried out on cores from the Bientina marsh, the Padule di Fucecchio and Lago Sibolla (Fig. 1; Paoli and Cellai Ciuffi 1980, 1983, 1984). The slight development of the woodland may be the consequence of damp soils or may result from woodland clearance for agricultural purposes, or both. Indeed, cultivating practices are revealed by numerous plant macrofossil remains from Neolithic and Bronze Age archaeological sites in the Pisa area (D’Amato Avanzi 1953; Nisbet 1987; Mori Secci 1991). Northwards, at the same time (about 5000 b.p.), pollen analysis carried out in Liguria near Chiavari shows the presence of a Mediterranean maquis mainly composed of Erica cf. arborea (Fig. 1; Guido et al. 2004a). Although the anthropogenic indicators are not present in high values, this kind of vegetation may be a consequence of long-term human action.
At Massaciuccoli, high percentages of Vitis together with low AP values were found between approximately 4200 and 2700 b.p. (MSC6). These two contemporary features could suggest an early agricultural practice, at least favouring this native plant. The analyses show the presence of Salix and Alnus in coincidence with both of the Vitis peaks (Fig. 3) and hint at the occurrence of swamp woods in which the grapevine grew wild in the form of a climber. Therefore, it could be hazardous to suggest that the grapevine was actually being cultivated, in the absence of significant percentages of anthropogenic indicators and/or archaeological evidence; a less ambitious interpretation could be that some form of management practice in the area, such as woodland clearance, indirectly favoured the spread of Vitis, and then its cultivation or management followed. At Pontedera, not far from Pisa, stored wild Vitis seeds were found in a Neolithic (5500 b.p.) archaeological site (Mori Secci 1991). Pollen analysis of deposits referred to the upper Holocene (Grassi et al. 2000) shows a strong development of wetland vegetation and the spread of human-induced habitats in the Massaciuccoli lake.
Investigations of coastal-marine sediments in Liguria have recorded the appearance and spread of fruit trees such as Olea, Juglans and Vitis, since at least 2500 b.p. (Guido et al. 2004c).
A decrease in the tree cover is recorded between 3800 and 3500 b.p. in central Italy (Lago di Mezzano, Sadori et al. 2004) and is interpreted as having been triggered by a climatic change towards drier conditions, successively intensified by human action; at the same time low percentages of Vitis occur. Vitis pollen probably related to agricultural practices was detected at Lago di Vico only after ca. 2630 b.p. (Magri and Sadori 1999). In northern Italy, there is a reduction of deciduous woodlands and the presence of Vitis micro- and macroremains around 2000 and 1100 B.C. (Lago Lucone, Valsecchi et al. 2006) and at the onset of the Bronze Age (Terramara di Montale, Mercuri et al. 2006).
Pollen analysis of a valley mire in southwest Spain showed two Vitis events at approximately 4500 and 3500 b.p. The first event, accompanied by Salix, was interpreted as a spontaneous expansion, while the second event, accompanied by disturbance indicators, was interpreted as being a signal of cultivation (Stevenson and Moore 1988). In Greece an increase in Vitis pollen shortly after 3500 b.p. is interpreted as a spread of the swamp woods where Vitis was growing (Crete, Bottema and Sarpaki 2003). The authors remark on the high pollen productivity and dispersal of wild vines and the under-representation of the cultivated forms. An intermediate pollen productivity is supposed to have occurred during the transition from the dioecious wild form to the hermaphrodite cultivated grapevine. A slight increase in Vitis, cautiously read as a signal of an agricultural phase, contemporary with the Celto-Ligurian civilization, was detected in southern France around 2000 b.p. (lower Rhone valley, Andrieu-Ponel 2000).
At Massaciuccoli, Vitis disappears together with a drop in the AP percentage mainly due to a large quantity (45%) of Chenopodiaceae. Since determination of Chenopodiaceae pollen at a lower taxonomic level is not always possible, interpreting this event unequivocally is hard. However, it is important to keep in mind that the site is located in an area adjacent to the coast that represents a natural habitat of many of the Chenopodiaceae. Consequently, their peak could be considered as a signal of an enlargement of brackish environments in the Massaciuccoli lake basin, also according to the fluctuation of the shoreline (Lambeck et al. 2004; Fazzini and Maffei 2000).
Research data show that a cooler climatic phase began during the Etruscan epoch (Lamb 1995), causing the spread of Fagus in the area, at lower altitudes than the present ones. Unfortunately the dating of this event is not clear. The spread of Fagus was hypothesized by Negri (1927) and confirmed by investigations carried out on the Apuane coastal plains (Ferrarini 1969) and in the Pisa cores (Trinci 2003). A similar condition is also suggested by the analyses in the most ancient sediments, of uncertain date, at the archaeological site of Pisa San Rossore, where the subsequent Fagus decline coincides with the expansion of deciduous Quercus (Begliomini et al. 2003; Mariotti Lippi et al. 2006). At the same time in the Apennines, decreasing pollen percentages of Abies were recorded, until its disappearance in the late Middle Ages (Lowe et al. 1994b; Guido et al. 2004c).
At Pisa San Rossore, the integration between pollen and sedimentological data has allowed some inferences on the dynamics of the warm Roman period and indicates that the area was affected by century-scale hydroclimatic crises triggering catastrophic floods (Mariotti Lippi et al. 2006).
Around 2400 b.p., the coastal plain of eastern Liguria in the surroundings of Rapallo was characterized by swampy woodlands dominated by Alnus, Cyperaceae, Juncaceae and abundant aquatic plants (Fig. 1; Guido et al. 2004a). Abies was widespread in the surrounding hills, its pollen reaching 30% of the total. This is probably the sign of an unexploited coastal wetland, at the mouth of a small stream. Similar wetlands were widespread in western Liguria, as observed at the mouth of the river Centa (Fig. 1; Arobba et al. 2004).
The top part of the sequence (MSC7) represents approximately the last 2000 years and is characterised by an increase in Mediterranean evergreen taxa suggesting a drying up of the area. This may be attributed to the human attempts to make new land available for agricultural purposes by draining wetlands. However, this situation must not have lasted for a long time, since hygro-hydrophilous trees and especially herbs become more abundant, reaching their highest values.
Conclusion
The general outline emerging from the Massaciuccoli core suggests that at the beginning of the Holocene the area was mainly covered by deciduous woodlands, in which Quercus, Corylus and Alnus were dominant. Then Abies spread and reached high values between about 9000 and 7500 b.p. After this, the pollen record is interrupted by a thick sandy layer resulting from a marine ingression (Antonioli et al. 2000).
At about 5500 b.p. deciduous woodlands, which were reduced in the most recent levels, again covered the area. Numerous vegetational oscillations were recorded, although there was not evidence of drastic changes. These oscillations can probably be connected to the sea level variations that would have had a strong influence on the local water table, rather than directly to global climatic changes. Furthermore, the MSC pollen record does not show evidence of the arid phases revealed by other south-western European sequences (Jalut et al. 2000). Particularly, no clear arid phase is detectable in the late Holocene, around 4500–4000 b.p., as recently observed in Mediterranean islands (Perez-Obiol and Sadori 2006). The Massaciuccoli basin is, indeed, a poorly drained area with low sensitivity to climate changes; these did not determine all the plant extinctions observed elsewhere, probably because of the mitigation effect of the sea and the shelter of the mountains, which in this area run parallel to the coastline, at a very short distance from it. This peculiarity is confirmed by its refuge role for relict plants, such as the elements of the Sphagno-Droseretum rotundifoliae Tomei et al. (1997).
The occurrence of high percentages of Vitis pollen for an apparently long period (starting from ca 4200 b.p.) is interpreted as a possible clue of an early management of the wild plant.
The data suggests that during the Holocene, along this part of the coast, thermophilous evergreen plants are scarcely represented. The spread of Mediterranean evergreen vegetation in the study area was probably often favoured by exploitation of deciduous woodlands, land reclamation, or other human activities. This is also suggested by the recent history of the area indicating that each time human management decreases, swamp conditions become newly established (Barsanti 2006). Human impact as an important factor in the Late Holocene spreading of the sclerophyllous vegetation in moister northern Mediterranean regions was also considered by Reille and Pons (1992).
All the investigations carried out along the coast of the Mar Ligure have shown the past presence of coastal wetlands which have allowed the preservation of macro- and microremains thanks to which we attempt to read the vegetation history of the area today. Alnus swamps must have been widespread in the damp environments behind the dunes along the coastal plains, and especially near river mouths. These environments have practically disappeared today, and even along the major water-courses they are strongly reduced. The reduction of these wetlands seems to have advances parallel with the beginning of farming in the area, when the people were attempting to drain portions of the swamps for cultivation by the construction of drainage canals. This process became much more pronounced in historical times, after the Roman period, and especially after the Middle Ages, up to the last century (Ceccarelli Lemut et al. 1994; Cori and Lombardi 1994).
References
Abbate Edlmann ML, Bargelli S, Giachi G (1991) I carboni nell’insediamento preistorico del Podere Casanuova. In: Aranguren BM, Ducci S, Perazzi P (eds) Il villaggio neolitico di Podere Casanuova (Pontedera, Pisa). Rivista di Scienze preistoriche 43:225–228
Abbate Edlmann ML, Bargelli S, Giachi G (1993) Indagine sulla vegetazione nei dintorni dell’insediamento preistorico del podere Casanova in Pontedera (Pisa), attraverso il riconoscimento del legno e dei reperti antracologici. Acta Naturalia de “L’Ateneo Parmense” 29:73–87
Accorsi CA, Rodolfi G (1975) Primi risultati sullo studio di un suolo calcimorfo delle Alpi Apuane in relazione ad analisi palinologiche e microbiologiche. Boll Soc Ital Scienze Suolo 9:35–51
Alessio M, Bella F, Bachechi F, Cortesi C (1965) University of Rome Carbon-14 Dates III. Radiocarbon 8:213–222
Allen JRM, Watts WA, McGee E, Huntley B (2002) Holocene environmental variability – the record from Lago Grande di Monticchio, Italy. Quat Internat 88:69–80
Andersen ST (1979) Identification of wild grass and cereal pollen. Danm geol Unders, Årbog 1978:69–92
Andrieu-Ponel V, Ponel P, Bruneton H, Leveau P, de Beaulieu J-L (2000) Palaeoenvironments and cultural landscapes of the last 2000 years reconstructed from pollen and Coleopteran records in the Lower Rhone Valley, southern France. The Holocene 10:341–355
Antonioli F, Girotti O, Improta S, Nisi MF, Puglisi C, Verrubbi V (2000) Nuovi dati sulla trasgressione marina olocenica nella pianura versiliese. In: Barchesi P, Angelelli A, Forni S (eds) Atti del Convegno “Le Pianure – Conoscenza e salvaguardia”, Ferrara, 8–11 Novembre 1999, Ferrara, pp 214–218
Arobba D, Caramiello R, Firpo M (2001) Studio paleobotanico e sedimentologico di un sondaggio alla foce del Fiume Centa (Albenga, Liguria occidentale). Allionia 38:147–158
Arobba D, Caramiello R, Firpo M (2004) Contributi paleobotanici alla storia dell’evoluzione di una pianura costiera: il caso di Albenga. In: de Marinis RC, Spadea G (eds) I Liguri. Skira, Ginevra-Milano, pp 76–78
Arrigoni PV (1990) Flora e vegetazione della Macchia lucchese di Viareggio (Toscana). Webbia 44:1–62
Barsanti D (2006) L’assetto del territorio. In: Bertuccelli Migliorini A V, Caccia S (eds) Mirabilia maris: visioni cartografiche e resoconti di viaggio Edizioni ETS, Pisa, pp 51–63
Begliomini V, Benvenuti M, Mariotti Lippi M, Pallecchi P, Sagri M (2003) Il contesto paleoambientale dell’antico porto di Pisa. In: Bruni S (ed) Il porto urbano di Pisa Antica. La fase ertrusca, il contesto e il relitto ellenistico. Silvana ed, Cinisello Balsamo (MI), pp 103–107
Bertolani Marchetti D, Dallai D, Mori Secci M, Trevisan Grandi G (1994) Palynological evidence and forest events in the upper Tuscan/Emilian Apennines in the context of the whole Apennines Holocene history. Fitosociologia 26:145–164
Bottema S, Sarpaki A (2003) Environmental change in Crete: a 9000-year record of Holocene vegetation history and the effect of the Santorini eruption. The Holocene 13:733–749
Branch N (2004) Late Würm Lateglacial and Holocene environmental history of the Ligurian Apennines, Italy. In: Balzaretti R, Pearce M, Watkins S (eds) Ligurian landscapes: studies in archaeology, geography and history. Accordia specialist studies on Italy 10, Accordia Research Institute, University of London, pp 7–69
Brandolini P, Calderoni G, Guido MA, Montanari C, Nicchia P, Ramella A, Terranova R (2002) Emergencias geomorfológico-paleoambientales de los depósitos cuaternarios en la llanura fluvial costera del torrente Polcevera – Liguria, norte de Italia. Revista de Geologia Aplicada a la Ingenieria y al Ambiente 17:29–38
Ceccarelli Lemut ML, Mazzanti R, Morelli P (1994) Il contributo delle fonti storiche alla conoscenza della geomorfologia. In: Mazzanti R (ed) La pianura di Pisa e I rilievi contermini. Società Geografica Italiana, Roma, pp 401–429
Cori B, Lombardi D (1994) La pianura di Pisa nell’età contemporanea. Aspetti geo-economici. In: Mazzanti R (ed) La pianura di Pisa e I rilievi contermini. Società Geografica Italiana, Roma, pp 465–491
Cornara L, Guido MA, Montanari C, Placereani S (1996) Analisi polliniche. In: Melli P (ed) “La città ritrovata”. Tormena Ed, Genova pp 373–375
Corti R (1955) Ricerche sulla vegetazione dell’Etruria.X. Aspetti geobotanici della selva costiera. La selva pisana a S. Rossore e l’importanza di questa formazione relitta per la storia della vegetazione mediterranea. Nuovo Giorn Bot Ital 62:75–262
Cremonesi Grifoni R, Tozzi C (1994) Gli insediamenti dal Paleolitico ell’età del Bronzo. In: Mazzanti R (ed) La pianura di Pisa e I rilievi contermini. Società Geografica Italiana, Roma, pp 153–182
D’Amato Avanzi MG (1953) Il grano della popolazione con civiltà tipo Lagozza della grotta di Agnano (Pisa). L’agricoltura italiana 53 (VIII n.s.):1–9
Digerfeldt G, de Beaulieu J-L, Guiot J, Mouthon J (1997) Reconstruction and paleoclimatic interpretation of Holocene lake-level changes in Lac de Saint-Léger, Haute-Provence, southeast France. Palaeogeogr Palaeoclim Palaeoecol 136:231–258
Fægri K, Iversen J (1989) In: Fægri K, Kaland P, Krzywinski K (eds) Textbook of pollen analysis, 4th edn. Wiley, Chichester
Fazzini P, Maffei M (2000) The disappearence of the city of Luni. J Cultur Heritage 1:247–260
Federici PR (1993) The Versilian transgression of the Versilian area in the light of drillings and radiometric data. Mem Soc Geol It 49:217–225
Ferrarini E (1969) Reperti fossili di faggeta nella pianura costiera apuana – Analisi polliniche In: Francini Corti E, Reperti fossili. Analisi polliniche e dei legni subfossili. Quaderni della Ricerca scientifica, 60, Scienze sussidiarie dell’archeologia. CNR, Roma, pp 65–66
Ferrarini E (1977) Studi sulla vegetazione litoranea di Massa (Toscana). Mem Accad Lunig Sc “G. Capellini” 41:3–44
Galletti Fancelli ML (1971) Ricerche sulla pianura pisana- I analisi polliniche di sedimenti quaternari lacustri della zona di Pontedera (Pisa). Atti Soc Tosc Sci Nat, Mem, Serie A 78:118–134
Galletti Fancelli ML (1979) Ricerche sulla subsidenza della pianura pisana. Analisi polliniche di sedimenti quaternari della pianura costiera tra Pisa e Livorno. Boll Soc Geol It 98:197–245
Garbari F (2000) La flora di S. Rossore (Pisa) aggiornata al 1999. Atti Soc Tosc Sc Nat, Mem, Serie B 107:11–42
Garbari F, Cecchi O (2000) Solidago litoralis Savi (Asteraceae): typification, taxonomy and distribution of a plant endemic of the N-W Tyrrhenian seashores of Tuscany. Webbia 54:183–192
Gellini R, Pedrotti F, Venanzoni R (1986) Le associazioni forestali ripariali e palustri della selva di S. Rossore (Pisa) Docum Phytosociol 10:27–41
Giardini M (2006) Late Quaternary vegetation history at Stracciacappa (Rome, central Italy). Veget Hist Archaeobot DOI 10.1007/s00334-006-0037-y
Goeury C (1997) Gestion, traitement et représentation des données de la paléoécologie. XVéme Symposium de l’APLF, Lyon, p 31
Grassi R, Mariotti Lippi M, Zanchetta G, Bianciardi T, Bonadonna FP (2000) Studio preliminare di due sondaggi superficiali eseguiti nella piana della Bassa Versilia (Toscana Nord-Occidentale). In: Barchesi P, Angelelli A, Forni S (eds) Atti del Convegno “Le Pianure – Conoscenza e salvaguardia” Ferrara 8–11 Novembre 1999. Ferrara, p 219
Guido MA, Mariotti Lippi M, Menozzi BI, Placerani S, Montanari C (2004a) Ambienti costieri nella riviera ligure di Levante tra le età del Bronzo e del Ferro: aree di Rapallo e di Chiavari. In: de Marinis RC, Spadea G (eds) I Liguri. Skira Ginevra-Milano, pp 78–81
Guido MA, Mariotti Lippi M, Menozzi BI, Placerani S, Montanari C (2004b) Il paesaggio vegetale della costa toscana settentrionale negli ultimi tre millenni a.C. In: de Marinis RC, Spadea G (eds) I Liguri. Skira, Ginevra-Milano, pp 84–85
Guido MA, Mariotti Lippi M, Menozzi BI, Placerani S, Montanari C (2004c) Il paesaggio vegetale montano della Liguria centro-occidentale nell’età del Ferro: area del monte Beigua (Savona). In: de Marinis RC, Spadea G (eds) I Liguri. Skira, Ginevra-Milano, pp 91–95
Jalut G, Esteban Amat A, Bonnet L, Gauquelin T, Fontugne M (2000) Holocene climatic changes in the Western Mediterranean, from south-east France to south-east Spain. Palaeogeogr Palaeoclim Palaeoecol 160:255–290
Lamb HH (1995) Climate, history and modern world. 2nd edn. Routledge, London
Lambeck K, Antonioli F, Purcell A, Silenzi S (2004) Sea-level change along the Italian coast for the past 10,000 yr. Quatern Sci Rev 23:1567–1598
Lowe JJ, Branch N, Watson C (1994a) The chronology of human disturbance of the vegetation of the Northern Apennines during the Holocene. Monografie di “Natura Bresciana” 20:169–187
Lowe JJ, Davite C, Moreno D, Maggi R (1994b) Holocene pollen stratigraphy and human interference on the woodlands of the northern Apennines, Italy. The Holocene 4:153–164
Magri D, Sadori L (1999) Late Pleistocene and Holocene pollen stratigraphy at Lago di Vico, central Italy. Veget Hist Archaeobot 8:247–260
Marchetti M (1935) Una torba glaciale del Lago di Massaciuccoli. Atti Soc Tosc Sc Nat, Proc Verb 43–44:143–150
Marchetti M, Tongiorgi E (1936) Ricerche sulla vegetazione dell’Etruria Marittima. VII. Una torba glaciale del Lago di Massaciuccoli (Versilia). Nuovo Giorn Bot Ital 4:872–884
Mariotti Lippi M, Bellini C, Trinci C, Benvenuti M, Pallecchi P, Sagri M (2006) Pollen analysis of the ship site of Pisa San Rossore, Tuscany, Italy: the implications for catastrophic hydrological events and climatic change during the late Holocene. Veget Hist Archaeobot. DOI 10.1007/s00334-006-0070-x
Mazzanti R, Nencini C (1994) La morfologia. In: Mazzanti R (ed) La pianura pisana e i rilievi contermini. Società Geografica Italiana, Roma, pp 89–102
Menozzi BI, Fichera A, Guido MA, Mariotti Lippi M, Montanari C, Zanchetta G, Bonadonna FP, Garbari F (2003) Lineamenti paleoambientali del bacino del Lago di Massaciuccoli (Toscana Nord-occidentale, Italia). Atti Soc Tosc Sci Nat, Serie B, 109 (2002):177–187
Mercuri AM, Accorsi CA, Bandini Mazzanti M, Bosi G, Cardarelli A, Labate D, Marchesini M, Trevisan Grandi G (2006) Economy and environment of Bronze Age settlements – Terramaras – on the Po Plain (Northern Italy): first results from the archaeobotanical research at the Terramara di Montale. Veget Hist Archaeobot 16:43–60
Montanari C, Guido MA, Cornara L, Placereani S (1998) Tracce polliniche di boschi neolitici di abete bianco in Val Bisagno (area urbana di Genova). Biogeographia 19:133–143
Montelucci G (1964) Ricerche sulla vegetazione dell’Etruria. XIII. Materiali per la flora e la vegetazione di Viareggio. Webbia 19:73–347
Moore PD, Webb JA, Collison ME (1991) Pollen analysis, 2nd edn. Blackwell, Oxford
Mori Secci M (1991) Analisi dei semi e dei frutti rinvenuti negli scavi di Podere Casanuova. In: Aranguren et al. (eds) Il villaggio neolitico di Podere Casanuova (Pontedera, Pisa). Rivista di Scienze preistoriche 43:229–234
Negri G (1927) Come si possa ricostruire la fisionomia della vegetazione della Toscana durante il periodo etrusco. Studi Etruschi 1:3–13
Nicol-Pichard S, Dubar M (1998) Reconstruction of the Late-glacial and Holocene environments in south-east France based on the study of a 66-m long core from Biot, Alpes Maritimes. Veget Hist Archaeobot 7:11–15
Nisbet R (1987) L’utilizzazione delle piante al Riparo del Lauro durante l’età del Bronzo. In: Cocchi Genik D. (ed) “Il riparo del Lauro di Candalla nel Quadro del Bronzo medio iniziale dell’Italia centro-occidentale”, pp 175–183
Paoli P, Cellai Ciuffi G (1980) Ricerche palinologiche nel Padule di Fucecchio. In: Min. Agr. e Foreste. Progetto Pilota per la salvaguardia e valorizzazione del padule di Fucecchio. Giorgi & Gambi, Firenze, pp 265–280
Paoli P, Cellai Ciuffi G (1983) Contributo palinologico allo studio dei sedimenti stratigrafici del laghetto di Sibolla. In: Min Agr e Foreste. Progetti di studi per la salvaguardia del laghetto di Sibolla. Menegazzo, Lucca, pp 81–88
Paoli P, Cellai Ciuffi G (1984) Lineamenti vegetazionali e paleobotanici del territorio di Montelupo Fiorentino. Quaderni del Museo della ceramica e del territorio 1:19–25
Perez-Obiol R, Sadori L (2006) Similarities and dissimilarities, synchronisms and diachronisms in the Holocene vegetation history of the Balearic Islands and Sicily. Veget Hist Archaeobot. DOI 10.1007/s00334-006-0038-x
Punt W et al (1976–1996) The Northwest European Pollen Flora (NEPF) Vol I (1976), Vol II (1980), Vol III (1981), Vol IV (1984) Vol V (1988), Vol VI (1991), Vol VII (1996), Elsevier, Amsterdam
Rapetti F, Vittorini S (1994) I caratteri del clima. In: Mazzanti R (ed) La pianura pisana e i rilievi contermini. Società Geografica Italiana, Roma, pp 103–132
Reille M (1992–98) Pollen et spores d’Europe et d’Afrique du Nord. Laboratoire de botanique historique et palynologie, Marseille
Reille M, Pons A (1992) The ecological significance of sclerophyllous oak forests in the western part of the Mediterranean basin: a note on pollen analytical data. Vegetatio 99–100:13–17
Russo Ermolli E, di Pasquale G (2002) Vegetation dynamics of south-western Italy in the last 28 kyr inferred from pollen analysis of a Tyrrhenian Sea core. Veget Hist Archaeobot 11:211–219
Sadori L, Giraudi C, Petitti P, Ramrath A (2004) Human impact at Lago di Mezzano (central Italy) during the Bronze Age: a multidisciplinary approach. Quat Internat 113:5–17
Stevenson AC, Moore PD (1988) Studies in the vegetational history of S. W. Spain. IV. Palynological investigations of a valley mire at El Acebron, Huelva. J Biogeogr 15:339–361
Terhürne-Berson R, Litt T, Cheddadi R (2004) The spread of Abies throughout Europe since the last glacial period: combined macrofossil and pollen data. Veget Hist Archaeobot 13:257–268
Tomei PE, Amadei L, Garbari F (1986) Données distributives de quelques angiospermes rares de la région méditerranéenne d’Italie. Atti Soc Tosc Sc Nat, Mem, Serie B 92:207–240
Tomei PE, Guazzi E, Barsanti A (1995) Contributo alla conoscenza floristica delle paludi e del Lago di Massaciuccoli. In: Tomei PE, Guazzi E (eds) Il Bacino del Massaciuccoli, Collana di indagini tecniche e scientifiche per una migliore conoscenza del Lago di Massaciuccoli e del suo territorio, Pisa, pp 43–78
Tomei PE, Guazzi E, Barsanti A (1997) La carta della vegetazione delle paludi e del lago d Massaciuccoli. In: Lago di Massaciuccoli-13 ricerche finalizzate al risanamento. Ente Parco Regioale Migliarino, San Rossore, Massaciuccoli, pp 275–288
Tongiorgi E (1936) La variazioni climatiche testimoniate dallo studio paleobotanico della serie Fiandriana nella pianura della Bassa Versilia presso il Lago di Massaciuccoli. Nuovo Giorn Bot Ital 43:762–764
Trinci C (2003) Indagini palinologiche su sedimenti olocenici della pianura di Pisa nel sito del porto fluviale romano di S. Rossore. Master’s degree thesis. Università di Firenze
Vagge I, Biondi E (1999) La vegetazione delle coste sabbiose del Tirreno settentrionale italiano. Fitosociologia 36:61–95
Valsecchi V, Tinner W, Finsinger W, Ammann B (2006) Human impact during the Bronze Age on the vegetation at Lago Lucone (northern Italy). Veget Hist Archaeobot 15:99–113
Watson C, Branch N, Lowe JJ (1994) The vegetational history of the northern Apennines during the Holocene. Monografie di “Natura Bresciana” 20:153–168
Acknowledgements
The authors wish to thank Prof. F.P. Bonadonna and Prof. F. Garbari (University of Pisa) for access to the studied material and scientific support. Prof. L. Sadori (University “La Sapienza” Roma, Italy), Prof. R. Pérez-Obiol (University of Barcelona, Spain), and Dr. F. Bittmann (Lower Saxony Institute for Historical Coastal Research, Wilhelmshaven, Germany) for the improvements to the text. S. Placereani and T. Gonnelli for the technical assistance and Dr. A. Fichera for aid in pollen analysis.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Mariotti Lippi, M., Guido, M., Menozzi, B.I. et al. The Massaciuccoli Holocene pollen sequence and the vegetation history of the coastal plains by the Mar Ligure (Tuscany and Liguria, Italy). Veget Hist Archaeobot 16, 267–277 (2007). https://doi.org/10.1007/s00334-006-0090-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-006-0090-6