
Abstract
The Setesdal valley in South Norway runs north to south for 200 km, from alpine vegetation at 1200 m, passing the tree-line at around 1000 m, through Boreal forests, to Nemoral forest at sea level. The Holocene vegetation history and its altitudinal differentiation were reconstructed using pollen percentages and influx and plant macrofossil concentration records from four lakes along an altitudinal transect. During the early Holocene (c. 10500–8000 cal b.p.) Betula pubescens, Pinus sylvestris, Alnus, and Corylus expanded in the lowlands. Only Pinus and B. pubescens reached 1000 m asl (Lille Kjelavatn). Only B. pubescens reached Holebudalen (1144 m asl) at about the same time as it arrived in the lowlands. Between c. 8000–3000 cal b.p. mixed deciduous forest developed around Dalane (40 m asl) and to a lesser extent around Grostjørna (180 m asl), birch woodland with pine surrounded Lille Kjelavatn and birch woodland occurred at Holebudalen. From c. 3000 cal b.p. to present, the vegetation at Dalane hardly changed except for slight human impact and the immigration of Picea abies. At Grostjørna Pinus expanded. At Lille Kjelavatn Pinus disappeared and Betula became sparse as at the tree-line today. Betula retreated from Holebudalen thus leaving it above the tree-line in low-alpine vegetation. The strengths and weaknesses of pollen and plant macrofossil data were assessed for forest reconstructions. Where local pollen production is low, as near the tree-line, percentages of long-distance tree pollen can be misleadingly high. Pollen influxes of Betula and Pinus were much smaller near their altitudinal limits than at lower altitudes, although their macrofossils were equally abundant. The limited dispersal capacity of macrofossils documents the local presence of species and the character of the local vegetation, although macrofossils of some tree taxa are rarely found. Pollen and plant macrofossil evidence complement each other to provide a more complete reconstruction of Holocene tree-limits and tree-lines and hence climate changes, than either form of evidence alone.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The broad-scale Holocene movements of tree taxa through Europe were demonstrated using pollen data by Huntley and Birks (1983). However, smaller areas show complexity, particularly in mountainous landscapes. In the absence of human activity, the ecological factors affecting plant distribution are climatic (temperature, precipitation), edaphic and biotic. Broad-scale distributions of species are primarily influenced by regional climate (Dahl 1998). Once a species reaches climatic equilibrium, its geographical limits may be particularly sensitive to climate change. For example, at the tree-line, a small change in climate may cause large biotic changes (Kullman 1983, 2002a; MacDonald et al. 1993; Körner 1998). If past range-limit movements can be detected, environmental and climatic changes can be reconstructed (Jackson and Whitehead 1991; Barnekow 1999). The spread of tree species in the early Holocene is not necessarily limited by temperature, although obviously their minimum temperature requirements must be exceeded. The spread also depends on the biology of the species, such as their dispersal ability, longevity and time to first reproduction, their colonising ability on raw or mature soils, their competitive power against other species for light, shade tolerance, root competition and many other ecological factors (Iversen 1973; Andersen 1966; Berglund 1966; Birks 1986).
Pollen analysis has always been used to reconstruct past vegetation, and indirectly, past climate. However, the reconstruction of population density and spatial distribution of individual species by means of traditional pollen analysis is difficult (Hicks 1993, 1994). Most pollen reaches a lake by aerial and water transport from the catchment. Airborne pollen may be transported to the lake from both lower and higher altitudes. Pinus sylvestris pollen is particularly susceptible to long-distance dispersal (Hicks 1993; Birks et al. 1996). In contrast, records of plant macrofossils, such as needles, fruits and catkin and bud-scales, reflect the local presence of species more closely (HH Birks 1973, 2003; Dunwiddie 1987; Barnekow 1999; Birks and Birks 2000, 2003). In addition to smaller plant macrofossils, tree megafossils (i.e. trunks, stumps) have also been used to reconstruct former tree-lines through the Holocene (Moe 1979; Aas and Faarlund 1988, 1996, 1999; Kullman 1983, 1995; Kremenetski et al. 1998; Karlén 1998). However, megafossils represent chance preservation of individuals and their occurrence does not necessarily represent the tree species limit. The megafossil record is discontinuous in time and space (Birks et al. 1996).
Although plant macrofossils are useful in reconstructing local vegetation, several shortcomings have been demonstrated (Allen and Huntley 1999; Eide 2003). Macrofossils of several thermophilous deciduous trees are sparsely produced, poorly dispersed to lake sediments and may be poorly preserved and hard to identify. Thus it is difficult to estimate the abundance of such trees in the local forest except at a broad scale. In treeless vegetation, macrofossils of tree taxa at the tree-line may be wind-dispersed in small quantities above the tree-line (Kullman 2000; Molau and Larsson 2000; personal observation) and it may be unwise to interpret the occasional macrofossil of, for example, tree Betula or Pinus in terms of local presence. Evidence for the character of the local vegetation from the total macrofossil assemblage may help to decide if a tree taxon could have grown locally (Velle et al. 2005; Birks 2003). Ideally, a combination of pollen and macrofossil analysis provides a more reliable basis for the reconstruction of local forest development and tree-line changes (Watts 1979; Jackson 1989; Birks and Birks 2003).
The Setesdal valley crosses all but one (the south arctic) of the major vegetational ecotones in Norway (Moen 1998, Fig. 1). Since the vegetation zones are spatially compressed by altitude, migration delay is unlikely to be important and climate is thus assumed to be the major determinant of plant distribution within Setesdal. We studied an altitudinal transect of small lakes situated close to the present vegetational ecotonal boundaries, where a climatic change in the past that would have caused a change in vegetation that may be detectable from fossils preserved in the sediments. With this ecological setting, our aims were two-fold: first, to reconstruct the vegetation history in the Setesdal valley through the Holocene, with a focus on tree migrations and forest changes and hence the climatic fluctuations in southern Norway, and second, to compare and evaluate pollen and macrofossil analyses as tools for reconstructing forest and tree-line changes.

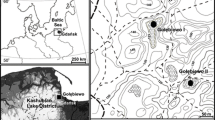
Map of southern Norway, showing the different vegetation zones (from Moen 1998), the Setesdal valley, (stippled line) and the lakes investigated. H = Haukeligrend
The sites
The Setesdal valley runs from 58°N to nearly 60°N for 200 km (Fig. 1) from the south coast and the temperate Nemoral deciduous forest region to the high-alpine zone on the Haukelifjell plateau. The pine tree-limit is at c. 900 m and the tree-line, formed by Betula pubescens s.l., is passed at about 1000 m. The range of mean annual temperature and precipitation along the Setesdal transect is 8°C to −2°C (Aune 1993) and 1500–3000 mm yr−1 (Førland 1993), respectively. As part of the Setesdal project, sediments from 11 small lakes have been pollen-analysed in all the vegetation zones (SM Peglar and HJB Birks unpublished). Because of difficulties encountered in ascertaining the local presence of taxa from the pollen record, macrofossils were subsequently analysed from 4 of the lakes (Fig. 1; Table 1).
The lake at Dalane (40 m asl) is in the Nemoral zone of mixed deciduous-coniferous forest. A small stream enters through a boggy area at its northwest end. A small farm to the north has no apparent influence on the lake. The basal marine sediments (Fig. 2a) show that a coastal bay was cut off from the sea by land uplift to form a freshwater lake. The bedrock is mainly acidic gneiss. The common trees are Pinus sylvestris, Populus tremula, Tilia cordata, Betula pendula, B. pubescens, and Quercus robur. The lake contains aquatic macrophytes (e.g. Potamogeton natans, Sparganium angustifolium, Callitriche spp., Nymphaea alba, Nuphar lutea) and is bordered by mesotrophic mires or cliffs. The steep cliffs on the north-western and south-eastern shores are cloaked by mixed forest, including Tilia cordata, Ulmus glabra and (rarely) Taxus baccata.


a. Selected pollen influx values (grains cm2 y−1) and macrofossil concentrations (number per 25 cm3 sediment) of trees and shrubs at Dalane (40 m asl), plotted against sediment depth. The estimated calibrated age of the sequence based on an age-depth model is shown on the left hand side. The pollen silhouettes enclose horizontal bars which indicate where the samples were counted. The macrofossils are presented as histograms unless very rare, when they are indicated by a ‘+’ outside the pollen curve. Type of macrofossil is given in brackets (A = anther, B = bark, C = capsule, F = fruit, L = leaf, N = needle, P = petiole, Sb = bud-scale, Sc = catkin scale, So = stomata, St = stem). Radiocarbon dates and calibrated ages are shown. The lithology is marine silt at the base, grading into fine detritus lake gyttja. b. % loss-on-ignition at 550°C, summary diagram of total terrestrial pollen and spores, together with selected pollen and spore percentages and macrofossil concentrations of ferns, herbs, sedges, grasses and Cristatella at Dalane plotted against sediment depth. Pollen percentages are presented as silhouette curves, with white silhouettes indicating ×10 values, of total terrestrial pollen and spores. Aquatic pollen and spore values are percentages of total terrestrial pollen and spores plus total aquatics. Type of macrofossil is given in brackets (Sp = sporangia, see caption to Fig. 2a for other symbols). ‘P’ (for productivity) indicates estimated high values of aquatic flora and fauna. All macrofossil values are given as number per 25 cm3 of sediments except taxa denoted by ‘*’ where abundance categories have been used
Grostjørna (180 m asl) lies in the Southern-Boreal vegetation zone dominated by conifers. The lake drains southwards and has an inflow from a tarn 120 m higher to the southeast. Acid granodiorite and hornblende gneiss dominate the bedrock. The vegetation is open Pinus sylvestris woodland with scattered Picea abies and Betula pubescens. Vaccinium myrtillus, Empetrum nigrum, and Deschampsia flexuosa dominate the field-layer. Macrophytes in the lake include Nuphar lutea, Nymphaea alba, and Utricularia vulgaris. The cliff at the northern end of the lake supports deciduous trees such as Quercus robur and Ulmus glabra. Bog vegetation borders the lake below the cliff and at the south shore.
Lille Kjelavatn (1000 m asl) is near the upper limit of the Northern-Boreal zone. It is 100 m above the present pine limit at 900 m and close to the birch tree-line. A small outflow drains northeast. Granite dominates the bedrock. The lake lies in a steep hollow and is surrounded by open birch forest, boggy peat shores, and steep scree slopes. The shrub layer is dominated by Salix glauca, with scattered Juniperus communis. Heaths, grasses and herbs of montane character form the field layer. Aquatic vegetation is sparse.
The lake at Holebudalen (1144 m asl) lies on Precambrian acid granitic gneiss in the low-alpine vegetation zone. The lake receives water from a small lake, Reintjørna, at 1318 m and drains steeply to the valley approximately 150 m below the site. The lake is about 100 m above the present tree-line of Betula pubescens. Grass-heath and snow-bed communities dominate the catchment, characterised by Salix herbacea, together with ferns, grasses, sedges, and bryophytes. South-facing slopes support some tall-herb communities with shrub-willows. No aquatic macrophytes were observed.
Materials and methods
Sediments
Sediment cores were obtained in the summers of 1996 and 1997 using a 5 cm-diameter modified piston corer (Wright 1991) from a raft moored over open water at the deepest point, usually in the centre of the lake. The upper unconsolidated sediments were retrieved using a Perspex tube sampler (Wright 1980) and were sub-sampled immediately by vertical extrusion into 1 cm thick segments. Two series of overlapping cores were taken from all lakes. Cores and samples were stored in the dark at 4°C before sub-sampling in the laboratory. Sediment lithology was described following Troels-Smith (1955). Organic content was estimated by loss-on-ignition (%LOI): 1 cm3 samples were taken at 1–2 cm intervals, dried at 105°C, weighed, heated at 550°C for 6 h, and the percentage weight loss calculated.
Pollen and macrofossil analyses
Weighed tablets of known Eucalyptus pollen content were added to 0.5 cm3 subsamples from the primary series of cores before preparation to estimate concentrations and influxes of pollen, spores, and charcoal. Samples were treated by the standard Method B of Berglund and Ralska-Jasiewiczowa (1986) and suspended in silicone oil (2000 cs). Pollen was counted over whole or half slides along equally spaced traverses at a magnification of ×400, using ×1000 for critical examinations, until a minimum sum of 500 terrestrial pollen and spores was reached. Identifications were aided by keys (HJB Birks 1973; Moore et al. 1991) and an extensive pollen reference collection. Pollen-type names follow the conventions of HJB Birks (1973).
Macrofossil samples were taken from the second series of cores. These were correlated with the first series using the %LOI. Samples spanning 1 cm depth were usually taken at two or four cm intervals, depending on the amount of change between analyses. Sample volume was measured by water displacement. If necessary, humic sediment was deflocculated for a few hours in dilute sodium pyrophosphate (Na4P2O7·H2O) or KOH (10%). The sediment was rinsed with water through a 125 μm mesh. Plant macrofossils were picked systematically from the residue using a stereomicroscope at ×12 magnification. Magnifications of ×40–66 were used for identification. Detailed examinations were made with a microscope with magnification of up to ×1000 and optional overhead illumination. An extensive reference collection, keys and atlases (see Birks 2001) aided identification. Plant nomenclature follows Lid and Lid (1994).
Pollen and macrofossil diagrams
Pollen and spore taxa of trees and shrubs are presented as accumulation rates (influxes) in Figs. 2a, 3a, 4a, and 5a. This avoids the inevitable down-weighting of rare taxa by a few species (e.g. Betula and Pinus) with high pollen production, since, in contrast to percentages, influxes of any taxon are independent of all other taxa. Selected aquatic and terrestrial pollen and spore taxa representing herbs, sedges and grasses are presented as percentages of total pollen (Figs. 2b, 3b, 4b, and 5b), since these taxa generally do not produce enough pollen to complicate the interpretations. Calculations and diagrams were made using the programs TILIA and TILIA.GRAPH (Grimm 1990) and then modified in Palaeo Data Plotter (Juggins 2002). The pollen diagrams were divided into pollen assemblage zones using constrained optimal sum-of-squares divisive partitioning (Birks and Gordon 1985) within the program ZONE (Juggins 1991). The statistical significance of the zones was established by comparison with the broken-stick model (Bennett 1996).


a. Selected pollen influx values and macrofossil concentrations at Grostjørna, plotted against sediment depth. The lithology is silt and sand at the base grading into fine detritus lake gyttja. Symbols are as in caption to Fig. 2a. b. % loss-on-ignition at 550°C, summary diagram of total terrestrial pollen and spores, with selected pollen and spore percentages and macrofossil concentrations at Grostjørna, plotted against sediment depth. Symbols are as in captions to Fig. 2a and b


a. Selected pollen influx values and selected macrofossil concentrations at Lille Kjelavatn plotted against sediment depth. Apart from a minerogenic base, the sequence consists of uniform fine detritus lake gyttja. incl. = including; other symbols are as in caption to Fig. 2a. b. % loss-on-ignition at 550°C, summary diagram of total terrestrial pollen and spores, with selected pollen and spore percentages and macrofossil concentrations at Lille Kjelavatn plotted against sediment depth. Symbols are as in captions to Fig. 2a and b


a. Selected pollen influx values and macrofossil concentrations at Holebudalen plotted against sediment depth. The lower sediments are coarse detritus lake gyttja grading into fine detritus gyttja above 945 cm. Symbols are as in caption to Fig. 2a. b. % loss-on-ignition at 550°C, summary diagram of total terrestrial pollen and spores, with selected pollen and spore percentages and macrofossil concentrations at Holebudalen plotted against sediment depth. Symbols are as in captions to Fig. 2a and b
Macrofossil taxa are presented either as concentrations per 25 cm3 of sediment or on a 0–5 scale (absent to abundant) (e.g. charcoal, fern sporangia, Nymphaeaceae astrosclereids (‘star cells’), Sphagnum leaves).


Comparison of pollen percentages (filled curves), pollen influx (open curves) and plant macrofossil concentrations (filled histograms) for selected tree taxa plotted against calibrated years b.p. at the four sites. The x-axis for each proxy is kept equal within the species, but an exaggeration of ×10 has been added for influx curves with very low values
Dating and age-depth modelling
Accelerator mass spectrometry (AMS) radiocarbon dates were obtained from small bulk-sediment samples pre-treated at the Laboratory for Radiological Dating, University of Trondheim and measured at the Ångstrøm Laboratory in Uppsala. It would have been preferable to date terrestrial plant material, but the macrofossil studies were made after the pollen analyses were completed and dated. It was assumed that any hard-water error in the dates would be minimal as all the sites are on non-calcareous bedrock. However, especially in the earliest sediments, a hard-water error may be present, resulting in dates that are a few hundred years too old. The radiocarbon dates (Table 2) were calibrated as calendar years (cal b.p.) before present (a.d.. 1950) using CALIB 4.3 (Stuiver and Reimer 1993) method A and the INTCAL98 bi-decadal calibration data-set (Stuiver et al. 1998). When calibrated, the ages often have substantial 1σ ranges (Table 2) that vary due to the irregularity of the relationship between14C and calendar years (radiocarbon wiggles and plateaux). For the same reason, close 14C ages can have widely different calibrated ages (Table 2). Age-depth models for the four cores were developed using a weighted mixed-effect regression model within the framework of generalised additive modelling (Heegaard et al. 2005) to derive the simplest parsimonious age-depth model for each sequence (Seppä and Birks 2001). The uppermost sediment was assumed to be modern and was assigned an age of −47 or −48±5 years b.p. for modelling purposes. It is desirable to construct age-depth models to provide ages for samples that are not directly dated and to enable pollen influx calculations. However, the considerable uncertainties associated with the dates themselves (laboratory measurements, inherent sedimentary sources of error) and then with calibration to calendar years, combined with the relatively few dates (6–8) for each sequence and the inherent uncertainties in statistical modelling lead to a large imprecision in any age-depth model and in the age estimates for individual samples (Telford et al. 2004). This has to be borne in mind when assessing the likely ages of events in the sequences. The age models are shown to the left of the diagrams (Figs. 2a, 3a, 4a, and 5a).
Results
Table 2 lists the radiocarbon dates and their calibrated ages. Pollen influxes and macrofossil concentrations of selected trees and shrubs are presented in Figs. 2a, 3a, 4a, and 5a, for Dalane, Grostjørna, Lille Kjelavatn, and Holebudalen respectively. Pollen percentages and macrofossil concentrations of selected herbs, sedges, and macrophytes are presented in the same order of sites in Figs. 2b, 3b, 4b, and 5b. A comparison of pollen influx, percentages and macrofossil data for selected tree taxa is presented in Fig. 6.
Discussion
The vegetational and climate development through the Holocene are discussed for each site, in time sectors of early-, mid-, and late Holocene. Details of the pollen zones and sub-zones are not discussed, since these are not the focus of this paper. The pollen and macrofossil records are compared for each site and between sites for the major tree taxa through the Holocene in Setesdal.
Vegetational and climatic development
DALANE (40 m asl); Figs. 2a and b
Younger Dryas-Holocene transition; c. 11500–10500 cal yearsb.p.(pollen zones DP-1a-b). The lowest sediments are marine, containing dinoflagellate cysts (hystrichospheres) and foraminifera. The plant remains (Fig. 2a and b) indicate a Younger Dryas age, as they reflect montane dwarf-shrub heath (Salix, Betula nana, Empetrum, Dryas), grassland and snow beds (Ranunculus glacialis). The absence of any pollen stratigraphic changes (Fig. 2b) implies rapid deposition. It is possible that the earliest Holocene sediments (c. 11500–10500 cal b.p.) have been lost during isolation of the basin.
Early Holocene; c. 10500–8000 cal years b.p. (pollen zones DP-2 to DP-4).
In pollen zone DP-2 the sediments change to freshwater deposits (Fig. 2a), with an increase in %LOI and pollen influx (Fig. 2b). The relatively slow sedimentation and low organic content make radiocarbon dating difficult. The calibrated age scale below 946 cm (9750±85 b.p./10340±320 cal b.p.) in Fig. 2a is a linear extrapolation and is probably not realistic. The early-Holocene record at Dalane before c. 8000 cal b.p. is compressed, making the calculation of the sediment accumulation rate and the estimation of the timing of changes in the fossil record difficult. Thus the peaks in Betula and Pinus pollen influx at 945 and 939 cm may be artificially exaggerated. In addition, the dating of bulk sediment may include a reservoir effect, which could be checked by dating terrestrial plant material (e.g. Tørnqvist et al. 1992). However, the reservoir effect was expected to be small as the bedrock is non-calcareous.
The basal pioneer assemblage developed into open Salix, Betula nana and Juniperus scrub with Empetrum, grasses, ferns, and damp-soil herbs. The large increase in the proportion of tree and shrub pollen through zone DP-2 in the summary pollen diagram (Fig. 2b) is probably a percentage artefact due to the low absolute amounts of both local and regional pollen deposited. This is supported by the macrofossil record (Fig. 2a) where only B. nana was found at the beginning of the Holocene. The Betula pollen maximum would seem to reflect local B. nana growth together with some long-distance tree-birch pollen (van Dinter and Birks 1996). Birch-forest seems not to have been established at the very beginning of Holocene, as was proposed by Aas and Faarlund (1988), but at the start of zone DP-3. This is tentatively estimated at c. 10000 cal b.p.; a firm conclusion should not be drawn because of the dating difficulties and model uncertainties. The peaks in Pinus pollen in zone DP-3 coincide with the start of the macrofossil record of bud scales, but the macrofossil numbers only expand substantially after the pollen maxima, suggesting that the latter might be partly a result of long-distance transport and/or partly an artefact of the slow and imprecisely calculated sedimentation rate (Figs. 2a and 6). One interesting aspect is that the macrofossil evidence indicates the arrival of pine soon after tree-birch (Fig. 2a) in contrast to the proposal by Hafsten (1987) that pine expanded some 400 years later than tree-birch. However, the slow sedimentation rate may exaggerate this impression. In the lake, the expansion of Isoetes echinospora, I. lacustris, Nymphaea, and the bryozoan Cristatella mucedo (Fig. 2b) indicates summer temperatures above c. 10°C.
Corylus pollen influx and percentages increase abruptly in zone DP-3, indicating the local arrival and expansion of Corylus, although its macrofossils (bud-scales) are not recorded until zone DP-4. These bud-scales preserve poorly and are hard to identify (Tomlinson 1985). In addition, the parent plants may have been growing too far from the lake for macrofossil remains to reach the sediments, whereas the pollen is widely wind-dispersed. Corylus arrived at a similar time elsewhere in Norway (Danielsen 1970; Hafsten et al. 1979; Kaland 1984). The gradual increases in deciduous-tree pollen (Fig. 2a) suggest that these taxa were spreading towards southern Norway (Fig. 2a). In the lake there is an increase in Isoetes spp., Nymphaeaceae and Potamogeton, which provided a habitat for Cristatella (Fig. 2b).
In zone DP-3 Populus tremula pollen increased. Although the influx curve rises in DP-2 and is high until the end of DP-4, Populus may actually not have been very abundant, considering the low influx values (Fig. 2a). Its macrofossils do not have a continuous record until DP-4, suggesting a low frequency in DP-3 if it was growing locally. Small stands may have occupied the nearby cliffs, perhaps together with Corylus, before Ulmus and other more competitive (i.e. shade tolerant) species arrived. According to Danielsen (1970) and Hafsten (1987), Ulmus arrived in southern Norway about 9450 cal b.p., compared to 9700 cal b.p. at this site. However, our date may be too old (see above) and has an uncertainty of about ±300 years. There is no macrofossil evidence for local growth until c. 300 years later (DP-4). This may be because Ulmus fruits and bud-scales, like Corylus bud-scales, do not preserve well. Consequently, their absence further up the macrofossil record (Fig. 2a) when the pollen values are much lower, may not mean that Ulmus was absent. Today Corylus and Ulmus have tetratherms (mean June-September temperature) of 12.5°C and 11.4°C (Helland 1912) respectively, suggesting an increase in summer temperature around 9000–9500 cal b.p. This would have led to improved soil fertility. In addition, nitrogen fixation by Alnus roots would also increase soil nitrogen content through the decomposition of Alnus leaves.
The expansion of Alnus pollen at the beginning of zone DP-4 (c. 9800 cal b.p.) is relatively early compared with some areas (e.g. Hafsten 1987; Huntley and Birks 1983) but not all areas (Bennett and Birks 1990) in north-west Europe. Local variation in the timing of the arrival and expansion of alder appears to be characteristic of its Holocene behaviour (Birks 1989; Bennett and Birks 1990). In addition, our date may include a reservoir effect. The large pollen influx from c. 9700 cal b.p. (Fig. 2a) suggests local occurrence and is soon followed by macrofossil evidence. It was not possible to separate A. glutinosa from A. incana. It has been suggested that A. glutinosa spread from the south and A. incana from the east (Tallantire 1974; Kaland 1984), so the Dalane records of Alnus fruits might be A. glutinosa. If so, the mean tetratherm temperature was at least 12.4°C (Helland 1912).
During zones DP-2–DP-4 the closed forest was dominated by Betula and Pinus, with Corylus and Ulmus. Alnus was common in wet areas and Populus and cf. Sorbus aucuparia grew in places with less shade, such as on the cliffs. Salix bushes occurred near the lake, and aquatic macrophytes flourished.
Mid Holocene; c. 8000–3000 cal years b.p. (pollen zone DP-5 to first half of DP-6.
During zone DP-5, the birch-pine forest was colonised increasingly by deciduous trees (Populus, Quercus, Alnus, Tilia, Fraxinus) that competitively reduced the pine and birch. The Nemoral vegetation type developed, which still persists today, especially on the south-east facing cliff above the lake. The pollen influx values of Tilia cordata rise sharply c. 8000 cal b.p. accompanied by macrofossils (Fig. 2a). This indicates the arrival of T. cordata in Norway 700–800 years earlier than previously suggested by Hafsten (1987); however see the comments on radiocarbon dating above. Fraxinus excelsior reached northern Europe around 6800 cal b.p. (Huntley and Birks 1983) and was the last warmth-demanding tree species to arrive in Norway. The delayed arrival was probably due to its shade intolerance and hence its need for forest openings (Birks 1989) which may have been rare in the closed Tilia, Ulmus, and Quercus forests. The low influx values and the absence of macrofossil evidence (only one macrofossil was found, just before 1000 cal b.p.) make it difficult to say when Fraxinus became locally established at Dalane, but it has evidently never been common. Quercus was uncommon at the beginning of this zone, but shows gradually increasing pollen influx values. Although its pollen record started at around the same time as that of Ulmus (10500 cal b.p.), it did not become an important component of the forest until c. 7000 cal b.p. (see also Danielsen 1970; Hafsten 1987) which coincides with the first record of its macrofossils. Taxus baccata was also present in zone DP-5, but was not common. It is often associated with calcareous soils, but if competition is not severe it can flourish on acidic soils, as it does in Setesdal today.
In the first half of DP-6, Corylus and Quercus increased and Pinus, Ulmus, and Fraxinus decreased. Tilia remained common. Around 4000 cal b.p. (early zone DP-6) a peak in Taxus pollen influx implies that Taxus expanded near the lake, perhaps on to the cliffs. It subsequently decreased but remained locally present. Mires developed around the lake, with large amounts of Juncus and, later, the presence of Drosera, and Myrica gale (Fig. 2b). This mire development may explain the disappearance of Alnus macrofossils around 2500 years cal b.p. (Fig. 2a), even though its pollen values remain high, since the trees may have been situated further from the lake and their fruits and bud-scales filtered out before they could reach the water (Wainman and Mathewes 1990). Three pollen grains of Picea recorded between 4000 and 3000 cal b.p. cannot be taken as evidence of local spruce occurrence (Geisecke and Bennett 2004; cf. Segerström and Stedingk 2003).
Late Holocene; c. 3000 cal years b.p.-present (upper half of pollen zone DP-6 and whole of DP-7).
Mesotrophic mire development continued (Fig. 2b) with Carex, Poaceae, Drosera and Juncus spp. increasing. During DP-6, the record of Isoetes lacustris ceased, although I. echinospora persisted and occurs in the lake today. Its megaspores were not recorded.
In DP-7 the proportion of herbs, pteridophytes, and grasses increased relative to that of trees and shrubs (Fig. 2b), suggesting some forest clearance for cultivation, as indicated by Hordeum-type pollen (Fig. 2b) and/or grazing. Light-demanding trees such as Sorbus aucuparia, Fraxinus and Acer became more apparent. Anomalously, Populus pollen virtually disappeared but its macrofossils were still recorded. Low continuous influx of Picea pollen started c. 1500 cal b.p., suggesting that Picea had immigrated to the region. However, macrofossils (Fig. 2a) demonstrating its local presence at Dalane are not recorded until c. 700 cal b.p.
GROSTJØRNA (180 m asl); Figs. 3a and b
Early Holocene; c. 11500-c. 8000 cal years b.p. (pollen zones GR-1 to GR-4a).
Zones GR-1 and GR-2 contain characteristic late-glacial taxa such as Artemisia, Rumex spp., and Oxyria. The basal minerogenic sediments may represent the end of the Younger Dryas. The extrapolated age scale places the start of the Holocene (11500 cal b.p.) at the zone GR-1/GR-2 boundary, where Ericaceae and Salix pollen increased, quickly followed by ferns and Betula in a typical early Holocene succession. Later in GR-2, %LOI and species diversity increased (Fig. 3a and b). The vegetation was probably a mosaic with occasional Betula spp. and Populus tremula, mixed with Salix, heaths, and ferns.
Macrofossils show that tree-birch and poplar were present by c. 10800 cal b.p. (Fig. 3a). The arrival of Populus accords with Huntley and Birks (1983), who estimated that it arrived in southernmost Norway near 11150 cal (c. 10000 14C) b.p. Pinus arrived about 1000 years later (9900 cal b.p.). This is slightly earlier than at Dalane, where there are dating problems in the lowermost sediments. Alternatively, the bulk sediment dates at Grostjørna may be affected by a reservoir age. Although the underlying rocks are acidic, dates on terrestrial plant material would be desirable. However, it may also be that the vegetation did not develop gradually and in an ordered way, but rather in steps, where chance (‘historical accident’, Carrión 2002) was important for where and when species settled. Perhaps the coastal situation of Dalane was less favourable for seedling establishment than the more sheltered Grostjørna, where the mean July temperature is higher than at Dalane today (Table 1). Pinus and Betula have high pollen percentages and influx values in zones GR-2 and GR-3 but their macrofossils are sparse or absent. They were growing regionally but the macrofossil evidence suggests either they were not close to the site or that macrofossils were not transported into deep water (c. 13.5 m) at the coring site (HH Birks 1973).
Early in pollen zone GR-3a, a Juniperus pollen peak indicates its local abundance in the woodland (Fig. 3a). Simultaneously, Calluna vulgaris became established and remained common for the rest of Holocene, apparently unaffected by increasing shade (Fig. 3b). The pollen influxes of thermophilous deciduous tree taxa, especially Corylus, increase in zone GR-3 (Fig. 3a), suggesting their northward expansion. However, the only macrofossils, Betula and Salix, are sparse, suggesting that Grostjørna may have been an ineffective macrofossil trap during this period. In pollen zone GR-4a the forest became denser, with abundant Pinus and Betula trees (macrofossil evidence) and some Alnus and Populus. To our knowledge this is the earliest Holocene record of Alnus (9900 cal b.p., Fig. 3a) in Norway. Why it became established at Grostjørna prior to Dalane may be for the same reasons discussed above for pine, and birch, poplar (Bennett and, Birks 1990), or because of a dating anomaly. The occurrence of B. pendula macrofossils, and the fact that the Alnus is probably A. glutinosa (Tallantire 1974) suggests an increase in summer temperature. This is also suggested by the pollen records of Ulmus, Fraxinus, and Corylus in zones GR-3 and G-4 (tetratherms of 11.2, 12.4, and 12.5°C, respectively, Helland 1912), which probably grew locally judging from the relatively high influx values (Fig. 3a), although the species are not represented by macrofossils. Tilia pollen also started to increase in zone GR-4. Aquatic diversity and productivity gradually increased (Fig. 3b) with the arrival and expansion of Nymphaeaceae, Isoetes echinospora, I. lacustris, and Cristatella. The occurrence of Juncus spp. (Fig. 3b) suggests mire development around the lake.
Mid Holocene; c. 8000–3000 cal years b.p. (pollen zones GR-4b to lower half of GR-4c).
GR-4b stands out as the zone with the most terrestrial macrofossils (Fig. 3a). Deciduous forest must have been well established around the lake. Populus tremula, Fraxinus, cf. Sorbus, and perhaps Taxus baccata were probably on the cliffs. Alnus and Salix occurred in damp places and Pinus, Betula, Tilia, Corylus, Quercus, and Ulmus in drier habitats. Overall this suggests a further increase in temperature and soil maturation.
Corylus and Ulmus are not represented by macrofossils until c. 8500 and c. 7000 cal b.p. respectively, which is much later than suggested by Kaland and Krzywinski (1978) for their local arrival. As at Dalane, the macrofossils are recorded after the initial maximum pollen influx peaks that are considerably earlier, although the influx of both is lower than at Dalane (Fig. 6). Corylus pollen influx (Fig. 3a) suggests local occurrence from at least 10500 cal b.p. As at Dalane, Quercus trees were uncommon, not becoming important until c. 7000 cal b.p. (Fig. 3a), in agreement with the evidence of Danielsen (1970) and Hafsten (1987).
During zone GR-4b Juncus seeds increased considerably, indicating, together with the presence of Scirpus lacustris type and Carex pollen, an expansion of mesotrophic mires and swamps around the lake (Fig. 3b). In GR-4c there is a small decrease in the proportion of tree and shrub pollen and a marked decrease of Betula spp. and Pinus macrofossils, but not their pollen percentages or influx. Quercus pollen increased, which could be due to opening up of the forest. Records of Plantago spp. and, at about 2800 cal b.p., cereal pollen, suggest human activity in the area. Although macroscopic charcoal is most frequent in zone GR-4b, suggesting local burning, the slight increase in microscopic charcoal (Fig. 3b) in zone GR-4c might result from nearby human settlement. Nymphaeaceae increased in zone GR-4c (Fig. 3b) but other aquatics were unchanged.
Tilia, Ulmus, Fraxinus and Sorbus macrofossils are absent but pollen is present. If these species were growing locally on the cliff, the mire development below may have trapped the plant remains and prevented them from reaching the lake (Wainman and Mathewes 1990). In addition, fruit production of these taxa is relatively low and the non-woody Ulmus and Fraxinus fruits are not well preserved. Similarly their bud-scales are fragile and difficult to identify.
Late Holocene; 3000 cal b.p. – present day (pollen-zones upper half of GR-4c to GR-5).
In GR-4c the forest was dominated by Betula, Pinus, Alnus, and Quercus with scattered Corylus and Populus. The warmth-demanding trees were probably growing on the cliff, as suggested above, with the mire below trapping plant material. Picea pollen is first recorded at about 1600 cal b.p. (Fig. 3a) at the same time as extensive spruce-dominated forest had developed in Sweden (Huntley and Birks 1983, Geisecke and Bennett 2004). However, the amounts of pollen are very small, so a local occurrence at Grostjørna is not likely until perhaps 500 cal b.p. A reduction in forest diversity and density occurred in zone GR-5, when Fraxinus disappeared and Populus, Ulmus, Tilia and Quercus were substantially reduced. Juniperus and Calluna expanded in the openings, together with Myrica in mires. Betula gradually declined, leaving Pinus as the dominant tree, as it is today. Hordeum-type and Secale pollen indicate cultivation near the site. Floating-leaved macrophytes were still present but Isoetes spp. were reduced (Fig. 3b).
LILLE KJELAVATN (1000 m asl); Figs. 4aand b
Early Holocene; c. 9000–8000 cal b.p. (pollen zones LK-1 and LK-2a).
This sequence starts after the arrival of pine and birch. Their macrofossils are found in the basal sediments at around 9000 cal b.p. (Fig. 4a). Records of pine stumps (9300 cal b.p.) 1100 m a.s.l. on the Hardangervidda plateau (Moe 1979) and pine stumps of 9650 cal b.p. age on the eastern Hardangervidda (Aas and Faarlund 1988), also suggest that pine was growing at high altitudes by this time. Birch megafossils, in contrast, have never been dated to as old as this in southern Norway, perhaps due to poor preservation. However, with birch growing at Grostjørna and Dalane at 11000 and 10000 cal b.p. respectively and with 8900 year-old (cal b.p.) birch stumps at an altitude of 1288 m in southern Jotunheimen (Aas and Faarlund 1988), it is likely that birch was growing around Lille Kjelavatn at least by 10000 cal b.p. The Picea pollen grain recorded at c. 9600 cal b.p. could be the result of long-distance transportation from small populations in central Sweden (e.g. Kullman 2000; Giesecke and Bennett 2004).
Pollen and macrofossil assemblages (Fig. 4a and b) reflect Pinus - Betula woodland with cf. Sorbus and Juniperus and a field layer of ferns (Dryopteris-type), grasses, sedges and herbs. Low pollen influx of Ulmus, Tilia, Quercus, Corylus, and Alnus (Figs. 4a and 6) probably derives from the presence of these trees in the valleys. A slightly higher (c. 200 grains cm−2 yr−1) influx of Corylus and Alnus pollen suggests these taxa may be closer to Lille Kjelevatn, although they are both large pollen producers. Higher influx values than these (as at Dalane and Grostjørna) and perhaps also macrofossils would be expected if they were growing locally (Fig. 6). Isoetes echinospora and Potamogeton were already established in the lake.
In pollen zone LK-2a, a decrease in Juniperus and Empetrum may suggest that the forest became denser. Slightly increased thermophilous tree-pollen from the lowlands suggests the spread of trees, perhaps in response to climatic warming.
Mid Holocene; c. 8000–3000 cal b.p. (pollen zones LK-2b to upper part of LK-3).
In sub-zone LK-2b, a small increase in Empetrum nigrum, Calluna vulgaris, and Vaccinium spp. pollen and macrofossils, with increasing fern spores (though a gradual decrease in sporangia) reflects the acid soil conditions that favoured ericoids in dry areas and tall ferns in mesic areas within the birch and pine forest.
At the beginning of LK-3 Pinus and Betula dominated with some cf. Sorbus aucuparia. The increase in Juniperus and Empetrum macrofossils may suggest more open areas, but could also reflect a replacement of the ferns as podsol soils developed. Pine macrofossils decreased from 5400 cal b.p. and became almost completely absent at c. 4000 cal b.p. (Fig. 4a). Megafossils from this altitude in southern Norway have not been dated after c. 5300 cal b.p. (Moe 1979), also suggesting the lowering of the pine limit and the tree-line. The local disappearance of pine is associated stratigraphically with an increased inwash of sclerotia of the soil fungus Coenococcum and of Selaginella selaginoides megaspores and a decrease in %LOI, suggesting soil instability. Climatic cooling and increased precipitation may have caused the catchment changes, perhaps exacerbated by grazing and felling, reducing pine regeneration near to its climatic limit. The persistent occurrence of Plantago lanceolata pollen (Fig. 4b) indicates exploitation for grazing. The gradual decrease in the low influxes of long-distance pollen of the thermophilous trees including Alnus (Figs. 4a and 6) may reflect cooling at lower altitudes. On the whole, there appears to have been a general cooling of climate, approaching today's Northern-Boreal/ low-alpine conditions at Lille Kjelavatn. The higher lake productivity (Fig. 4b), indicated by increased Isoetes and Cristatella abundance, may have resulted from an increased input of mineral material and nutrients from the catchment.
Late Holocene; 3000 cal b.p. – present (pollen zones upper part of LK–3 to LK-5).
The continued decrease in tree-pollen indicates more open Betula woodland, mixed with Juniperus, dwarf shrubs (Empetrum, Calluna vulgaris, Ericaceae-type, Vaccinium-type) and herbs. The rise in Selaginella selaginoides microspores suggests an expansion of damp grassland and some soil instability. Picea pollen percentages reach c. 2% at about 2000 cal b.p., about four times those at Dalane and Grostjørna. However, its influx is much lower than at these sites, demonstrating the effect of increased percentages of long-distance transported pollen in sites with low local pollen production. The increase at Lille Kjelavatn occurs earlier than in southern Setesdal, supporting the inference of Huntley and Birks (1983), Hafsten et al. (1979) and Geiseke and Bennett (2004) that Picea spread from the east. Pollen evidence from upper Grungedal (H.J.B. Birks and S.M. Peglar unpubl.) and other valleys east of Haukeli (Fig. 1) shows its local presence here at about this time; this is probably the source of Picea pollen at Lille Kjelavatn (Birks et al. 2005).
By c. 1800 cal b.p., the vegetation resembled the modern vegetation, with sparse tree-Betula and extensive ericaceous-Juniperus dwarf-shrub heath with Gentianella campestris-type and several sub-alpine/alpine species (e.g. Sagina and Selaginella selaginoides) growing in damp and more base-rich flushed areas on the surrounding slopes.
HOLEBUDALEN (1144 m asl); Figs. 5a and b
Early Holocene; c. 10800–8000 cal years b.p. (pollen zone HP-1)
No Pinus stomata were found on the pollen slides, but a bark fragment (Fig. 5a) was recorded at about 10000 cal b.p., indicating the possible local occurrence of pine at or near Holebudalen. Pine stumps at almost 1200 m elevation on the Hardangervidda and dated to c. 9650 cal b.p. (Aas and Faarlund 1988) suggest that this is not unexpected. However, pine was probably more common in the valley just below the site (e.g. round Lille Kjelevatn) and occasional bark fragments could have been blown to the site in winter over a smooth snow landscape. Local presence of pine is usually documented by abundant macrofossils (Fig. 6). In spite of the relatively low influx values for Betula and Salix (Fig. 5a), the occurrence of macrofossils suggests the presence of open birch woodland from c. 10800 cal b.p., with an understorey of willow and juniper bushes, dwarf shrubs, ferns, and herbs (Fig. 5b).
Pollen of Narthecium ossifragum and Humulus lupulus, both with altitude limits today 250–300 m below Holebudalen, together with the occasional pine macrofossils, suggests a warmer climate than today. This may be partly explained by early Holocene land uplift. At Finse (c. 80 km north) uplift is about 70 m since c. 10000 cal b.p. (Dahl and Nesje 1996), which is a reasonable estimate for uplift at Holebudalen. This implies the altitudinal limits of Pinus and Narthecium were 190–220 m higher than today. Applying a 0.6°C lapse rate per 100 m elevation results in tetratherm values 1–1.5°C higher than present during this period which accords with the presence of birch woodland round the site.
Mid Holocene; 8000-c. 2400 cal b.p. (pollen zones HP-2 and HP-3).
Pinus and Juniperus pollen influx gradually declined through zone HP-2, whereas influx from deciduous trees, such as Corylus, Tilia and Ulmus, slightly increased (Figs. 5a and 6), suggesting that Nemoral forest was more extensive in the lowlands. Both pollen percentages and influx of Alnus increased, particularly towards the top of the zone, where records of clumps of immature pollen may suggest the local growth of A. incana (Moe 1998). Similar influx values at Lille Kjelavatn have no associated evidence for local presence. However, the influx values are about one tenth of those at Dalane and Grostjørna (Fig. 6), where most of the Alnus pollen was probably from A. glutinosa. Open birch woodland around the lake probably had an understorey of juniper and willows, with grasses, tall-herbs and ferns. During this time Isoetes lacustris flourished in the lake. Cristatella was rare, probably being near its climatic limit.
Pollen zone HP-3 shows a large increase in ferns, higher dwarf-shrub pollen values than in HP-2, a gradual increase in herbs and Poaceae and a decrease in tree and shrub pollen (Fig. 5a and b). Terrestrial macrofossils and indicators of soil inwash (e.g. Coenococcum sclerotia) are sparse suggesting a stable catchment. This stability may reflect reduced snow-meltwater input from Reintjørna above Holebudalen and/or reduced solifluction on the surrounding slopes. Glacial evidence from the Hardangerjøkulen glacier some 80 km to the north indicates ice retreat between 6000 and 3800 cal b.p. (Dahl and Nesje 1996). Pollen percentages could suggest that Betula, Pinus, and Alnus were growing around the site during the first half of the zone, but the influx values are too low to suggest local presence, so the percentages are probably distorted by the lack of any high pollen producers growing locally. This inference is supported by the sparse macrofossil evidence, which suggests a few birch trees growing by the lake and the absence of Alnus. At the end of the zone, tree pollen and macrofossils decreased. Empetrum and Salix macrofossils increased (Fig. 4a, and b). One pine needle fragment and a few bark fragments were probably transported long-distance, funnelled up by the wind from the deep steep valley below, since there is no corroborating evidence for other lower-altitude species growing at Holebudalen at this time.
Increased inwash indicators at the end of the zone, the decline in tree-pollen influx and the absence of Betula macrofossils (Fig. 5a) are evidence for loss of woodland. The birch tree-line gradually dropped below the altitude of Holebudalen (1144 m) around 2500 cal b.p. Dahl and Nesje (1996) estimated that the pine tree-limit fell to its present altitude (c. 900 m) by approximately 2600 cal B.P
Late Holocene; c. 2400 cal years b.p. – present (pollen zones HP-4 and HP-5).
In pollen zone HP-4 tree-pollen values (Betula, Pinus and Alnus) continued to decrease, whereas fern spores increased as did many shrub (e.g. Salix, B. nana), dwarf shrub (particularly Empetrum), and herb taxa (Fig. 5a and b), indicating that the tree-line was migrating further from Holebudalen. After c. 2500 cal b.p. grassland and heath vegetation, characteristic of low-alpine areas, expanded around the lake. Soil stability decreased, as shown by an increased inwash of Coenococcum sclerotia and mycorrhizal root nodules (Fig. 5b) and the overall increased influx of macrofossils. Isoetes lacustris did not respond to increased minerals and nutrients but rapidly declined and disappeared (Fig. 5), suggesting that its thermal limit had been passed. However, its recorded maximum altitude today is about 1220 m. Local factors or within-lake processes may have caused its disappearance, possibly cooling by an increased inflow of snow meltwater from the Reintjørna catchment above. There is no evidence for tree-birch advance with climate warming over the last century around Holebudalen, in contrast to central Sweden (Kullman 2002a), although a few stunted seedlings were observed near the lake in 1997.
Tree and shrub pollen continued to decrease in zone HP-5, with a concomitant increase in herbs, dwarf shrubs, and pteridophytes (Fig. 5a and b). Pollen and macrofossils of taxa associated with snow-bed communities today (e.g. Salix herbacea) increased, suggesting a continued trend towards the development of the low-alpine vegetation of today. Picea pollen influx and percentages increased c. 1200 cal b.p., but the values are extremely low (Figs. 5a and 6). As Picea slowly spread westwards, its continuous pollen record started at Holebudalen about 800 years after its continuous record at Lille Kjelavatn. There is no subsequent increase, suggesting that Picea had reached its natural climatic limit.
Holocene tree migrations and climate changes in Setesdal
Our data reflect a rapid warming at the start of the Holocene and the arrival of pioneer taxa, birch (c. 10500 cal b.p.), and pine (c. 10000 cal b.p.) in the southern lowlands, although these unexpectedly late dates have large uncertainties. Birch spread through southern Setesdal and also from the eastern lowlands, rapidly reaching the south Norwegian mountains. It was already present at c. 10800 cal b.p. at 1144 m at Holebudalen but pine never expanded there (Fig. 6), suggesting that the tetratherm never exceeded 12°C. However, both birch and pine were present in the earliest sediments at Lille Kjelavatn (1000 m) at c. 9000 cal b.p. Birch trees were probably absent in the late-glacial in southern Norway (e.g. Birks 2003), but were present during the Younger Dryas in southern Sweden (e.g. Liedberg Jönsson 1988), Denmark (e.g. Jensen 1985) and the central Swedish mountains (Kullman 2002b). Pinus arrived in southern Setesdal about 400 years after birch. It was absent in south Norway during the late-glacial (Birks et al. 2005; cf. Kullman 2002b), the nearest late-glacial localities of the species probably being in Denmark (Bornholm), S. Sweden (Birks et al. 2005) and the central Swedish mountains (Kullman 2002b). It spread rapidly across northern Europe at the start of the Holocene (Huntley and Birks 1983), and reached altitudes around 1000 m (accounting for land uplift) by c. 9650 cal b.p. (Aas and Faarlund 1988).
The establishment of Ulmus and Corylus in southern Setesdal indicates that tetratherm temperatures exceeded c. 12.5°C by c. 10000 cal b.p. Their regional abundance in the lower valleys is reflected by rises in long-distance pollen influx at Holebudalen (Fig. 6). Alnus (probably A. glutinosa) arrived very early in south Setesdal, about 10500 cal b.p. at Grostjørna and 500 years later at Dalane, illustrating the spatially and temporally patchy early establishment and expansion of alder in Setesdal. A similar patchiness is described from the British Isles by Bennett and Birks (1990). Winters and springs must have been sufficiently mild for it to reproduce (McVean 1953). Although A. incana inhabits the middle valley today above the limit of A. glutinosa, we found the fossils difficult to separate, even the fruits (Tallantire 1974). However, no Alnus macrofossils were found at the mountain sites and the low pollen influx suggests pollen was blown from the valley below. If A. incana never reached these altitudes, the mean maximum summer temperatures were less than 17.2°C (Moe and Odland 1992). None of the sites shows evidence of, or response to, early Holocene climate oscillations.
Tilia reached Dalane about 8000 cal b.p. and persisted until today. It reached Grostjørna c. 1000 years later (Fig. 6). It was probably delayed by lack of suitable habitats rather than climate. Its macrofossil record ceased after c. 1500 years at Grostjørna and its pollen percentages and influx decreased, suggesting human activity may have affected it, although the important role of changes in a lake and its catchment in influencing deposition of fossils should not be ignored.
Human impact started to reduce forest cover in south Setesdal around 2000 cal b.p. The present mean July temperatures (Table 1) in south Setesdal are above the temperature requirements of the deciduous trees present during the Holocene, so any small (1–2°C) cooling would not have affected tree populations as they were not near their climatic limits. However, the situation was different at high altitudes. The near disappearance of birch trees at Holebudalen around 5300 cal b.p. and a clear thinning of pine trees at Lille Kjelavatn around 5000 cal b.p. (Fig. 6) correlate with glacier expansion phases at Hardangerjøkulen c. 6300–5200 b.p. (c. 7200–6000 cal b.p.) and Jostedalsbreen c. 6000–5500 b.p. (c. 6800–6300 cal b.p.) (Nesje et al. 1994). The development of low-alpine vegetation at Holebudalen and the decrease in birch trees at Lille Kjelavatn around 2500 cal b.p. (Fig. 7) can also, to some degree, be correlated with glacier expansion phases in southern Norway (Nesje et al. 1994). These two sites were near vegetation ecotones during the late Holocene and record the vegetation responses to climatic changes.

A summary of the main vegetational changes at the four sites Dalane, Grostjørna, Lille Kjelavatn, and Holebudalen, plotted against calibrated years b.p. The curves for deciduous trees are the sum of the pollen influx values for Corylus avellana, Ulmus, Tilia cordata and Quercus. At Holebudalen and Lille Kjelavatn, the deciduous influx curves were multiplied by ×5 to make them more visible. Filled circles represent macrofossil abundance. Vertical lines denote major changes
Picea spread to Setesdal in the late Holocene. It was present locally (macrofossils) at Dalane at c. 700 cal b.p., although its pollen values began to rise several centuries earlier. At Grostjørna no macrofossils were found, although its pollen influx became continuous (Fig. 3a) at the same time as its macrofossil occurrence at Dalane (Fig. 6). Today occasional trees are growing on the cliff at Grostjørna, but perhaps they are so few that the probability of finding macrofossils is very small. Native Picea is absent in the central Setesdal valley today and the pollen records (SM Peglar and HJB Birks unpubl.) indicate that it was spreading from the south. In contrast, continuous pollen influx of Picea started earlier at the high sites (c. 2000 cal b.p. at Lille Kjelavatn, c. 1200 cal b.p. at Holebudalen), reflecting its spread from the east (see above, Birks et al. 2005). Both these sites are well above its present altitudinal limit, although scattered spruce trees in the upper pine forests may attest its continuing spread towards the upper limit of pine today.
Comparison between pollen and macrofossils
Pollen and macrofossils differ in their reliability for the inference of the local presence of trees, both within and between species, and between sites (mainly due to catchment characteristics and lake-basin morphometry). We attempt to evaluate their strengths and weaknesses using the pollen and macrofossil records of the eight most common tree-pollen taxa (Betula, Pinus, Alnus, Corylus, Ulmus, Tilia, Quercus, and Picea). Pollen percentages and influx values and plant macrofossil concentrations for each taxon and locality are compared directly in Fig. 6. Fig 7 gives an overall synthesis of the data.
Betula
Most Betula macrofossils can be identified to species level, but it is difficult to differentiate consistently between the pollen of tree-Betula and Betula nana. At Dalane, macrofossils suggest that the first peak in Betula pollen influx around 10200 cal b.p. was from B. nana, with tree-birch arriving shortly after (Fig. 2a). The relatively low macrofossil concentrations (Figs. 2a and 6) suggest that birch trees were never abundant. At Grostjørna pollen influx values and tree-macrofossil expansions coincide (Fig. 6). There is no evidence for the local growth of Betula nana so the high Betula pollen percentages before the rise in influx at c. 10800 cal b.p. (Fig. 6) may be an effect of low local pollen production exaggerating the percentages of long-distance dispersed Betula pollen. The macrofossil record through the Holocene shows a relatively constant background concentration with a well-marked period of greater abundance from c. 9000–6500 cal b.p., which is not visible in either the pollen percentage or influx curves (Fig. 6).
At Lille Kjelavatn Betula pollen percentages are almost constant at c. 25%, only 5% lower than at Grostjørna and slightly more than at Dalane. Pollen influx and macrofossils correspond quite well (Fig. 6), suggesting a decrease from open birch forest from c. 9000 cal b.p. to the present-day conditions of solitary trees. The influx values are very much lower than in the lowland sites, but the concentration of macrofossils at Lille Kjelavatn is higher than at Dalane, suggesting that Betula pubescens pollen production is much reduced near its altitudinal limit. Without the macrofossil evidence, it would have been difficult to determine whether there were birch trees growing locally during the Holocene or not. The difficulty of interpreting a regional or local source of Betula pollen has also been demonstrated by Birks (2003). One cannot say that a certain level of Betula pollen influx represents the local growth of Betula without knowing more about the effects of altitude and climate on its biology.
At Holebudalen Betula pollen percentages are almost constant at c. 15–20% and influx is around 500 grains cm−2 yr−1 (Fig. 6), providing little information about when and how much birch was growing locally, except for a decline in the late Holocene. Macrofossils are necessary to elucidate the history of birch at this high-altitude site. Highest macrofossil concentrations during the early Holocene suggest the occurrence of open birch woodland, which then thins out and disappears around 2500 cal b.p. (Figs. 5 and 6). As birch fruits are effectively wind dispersed, especially over winter snow in a treeless landscape (HH Birks unpubl.), it is difficult to say whether single tree-birch macrofossils derive from local presence or remote growth in the valley below.
All four sites show the insensitivity of pollen percentages as a record of local birch-tree presence and abundance. Apart from the period of early Holocene abundance at Dalane, the pollen influx records also do not reliably indicate the abundance of trees. Influxes are high (c. 2000–3000 grains cm−2 y−1) at both Dalane and Grostjørna, although the concentration of macrofossils is much greater at Grostjørna, even during the period of highest influx (up to 6000 grains cm−2 yr-1) in the early Holocene at Dalane. This difference may reflect differences in the efficiency of the lakes as pollen and macrofossil traps; Grostjørna has an overall greater concentration of macrofossils especially in the mid Holocene. The basin characteristics influencing pollen and macrofossil sedimentation are virtually unknown, and the same factors probably influence the taphonomy of the fossils in different ways.
Pinus sylvestris
Due to its high pollen production and efficient dispersal, it is difficult to determine what values of pollen percentages and influx permit the presence of pine to be inferred with confidence (Hicks and Hyvärinen 1999; Birks et al. 1996). At Dalane, pollen percentages and influx both peak around 10000 cal b.p. (Fig. 6), whereas the macrofossils peak slightly later, indicating the time of local expansion of pine. The decrease in macrofossils is reflected in the pollen influx, whereas the pollen percentages are insensitive. At Grostjørna both pollen and macrofossils indicate a continuous pine-forest during the last 10000 years. However, the macrofossils give a more varied and detailed picture, first with an increase of pine from around 10000 cal b.p., then a pine-maximum between c. 9000–6000 cal b.p., followed by a lower level until the present.
At Lille Kjelavatn, pollen percentages are as high or higher than at Grostjørna throughout the sequence. However, the influx is c. 800 grains cm−2 yr−1 after 6500 cal b.p., which is less than half that at Grostjørna (mean c. 2000 grains cm−2 yr−1). Macrofossils, however, provide more precise information, suggesting the early Holocene development of pine-forest with a maximum density from 9000 to 5000 cal b.p. The decrease in pollen influx, whilst the macrofossils suggest a stable local population (Fig. 6), may be due to reduced pollen production as temperatures were gradually decreasing. The point at which pine disappears from the area cannot be determined from the influx values alone, but the lack of macrofossils after c. 4000 cal b.p. strongly indicates when pine most likely disappeared. This sequence clearly shows the importance of studying macrofossils at high altitudes, where it is difficult to distinguish between low local production and long-distance transported pollen. This is also the case at Holebudalen, where the few macrofossil records suggest when single pine trees may have been present nearby. However, small fragments of pine must be interpreted with care, and the same applies to the absence of macrofossils. The pine pollen influx is higher than at Lille Kjelavatn (c. 600–1000 grains cm−2 yr−1; Figs. 5a and 6) illustrating the difficulty of interpreting influx values in terms of local presence of trees, especially pine (Hicks and Hyvärinen 1999).
Alnus spp.
Macrofossils were found only at the lowland sites (Fig. 6), suggesting that Alnus never grew at 1000 m elevation. Except for the percentage and influx peaks between 9500 and 8500 cal b.p., the pollen and macrofossils correspond well at Dalane and Grostjørna. The low pollen percentage and influx at Lille Kjelavatn and Holebudalen indicate a long-distance source of Alnus pollen, although clumps of immature pollen were found in the mid Holocene at Holebudalen. Such clumps are not unambiguous evidence of local alder presence (but see Moe 1998). Kullman (1998) recorded Alnus glutinosa cones at 830m a.s.l. in central Sweden dated to 7310 b.p. (c. 8000 cal b.p.).
Corylus avellana
At Dalane and Grostjørna macrofossils indicate the local presence of Corylus during the second peak in the pollen percentage and influx curves. Most of the other pollen peaks are reflected in the macrofossils at Dalane. The discontinuous macrofossil record of Corylus may be explained by the distance from the lake at which it was growing or by the poor dispersal, preservation, and difficult identification of its bud-scales. At Lille Kjelavatn and Holebudalen there were no macrofossils (Fig. 6) and the relatively low pollen influxes represent long-distance pollen dispersal. Percentages at Holebudalen are as high as at Grostjørna but obviously should not be interpreted at face value.
Ulmus glabra
Apparently, the population density of U. glabra needs to be fairly high for its local presence to be registered in the sediments as macrofossils (Fig. 6). The reason probably lies in the poor preservation of the bud-scales and in their poor dispersal to the lake. Nevertheless, the records of Ulmus bud-scales at Dalane around 9000 cal b.p. coincide with peaks of pollen percentages and influx, confirming its local presence. A relatively rapid decrease in influx values from the maximum is followed by a gradual decrease in percentages and influx. At Grostjørna only one macrofossil of Ulmus was found, just after the pollen influx peaks, confirming local growth. As at Dalane, Ulmus decreased from c. 5000 cal b.p. Pollen percentages at Lille Kjelavatn and Holebudalen were at least as high as in the lowlands, again demonstrating the percentage exaggeration effect of abundant pollen production in the lowlands. The low pollen influx values and absence of macrofossils give no evidence for local growth of Ulmus near these lakes.
Tilia cordata
At Dalane pollen and macrofossils correlate well, both for the time of arrival and the relative abundances (Fig. 6). At Grostjørna macrofossils are present where the pollen records show a large double peak, but the pollen record begins much earlier with low values. At Lille Kjelavatn and Holebudalen there were no macrofossils and the low pollen influx values suggest no local growth of Tilia, but a detectable and consistent record of long-distance transported pollen is notable for such a relatively heavy and insect-dispersed pollen type. Kullman (1998) recorded a T. cordata inflorescence at 526 m a.s.l. in central Sweden, dated to 6980 b.p. (c. 7800 cal b.p.).
Quercus spp.
It is difficult to decide when Quercus arrived at Dalane from the pollen percentage and influx curves (Fig. 6). However, the macrofossils clearly indicate its latest time of arrival and this coincides with increases in the pollen curves. At Grostjørna the pollen influx peak around 4000 cal b.p. indicates that Quercus arrived earlier than indicated by the single macrofossil record at c. 2500 cal b.p. In general, in forested areas, occurrence of macrofossils of upland trees in lake sediment may be more or less accidental. At Lille Kjelavatn and Holebudalen pollen influx and percentages are very low (Fig. 6) and macrofossils are absent. Consequently, the influx curves only reflect the expansion of Quercus in the lowlands.
Picea abies
Increased influx values about 1500 cal b.p. at Dalane (Fig. 6) may indicate the arrival of spruce in the area, even though macrofossils are not present until c. 700 cal b.p. This may be a result of limited macrofossil dispersal in forested areas when trees are not growing next to the lake or the inflowing streams. At Grostjørna there is a good correlation between pollen percentages and influx, but the values are low and the absence of macrofossils makes it difficult to say when Picea arrived locally. It is rare near the site today. The pollen percentages at Lille Kjelavatn are much higher than at any other site, due to the exaggerated representation of long-distance pollen from spruce populations spreading up the valley from the east. Even at Holebudalen the percentages are as high as at Dalane where spruce was locally present. However, the influx values are extremely low at both sites.
The vegetation history of the Setesdal valley is summarised in the time-space diagram (Fig. 7). The largely climatic differentiation of the vegetation zones along the altitudinal gradient has always been present through the Holocene. In the early Holocene Betula was the first tree, closely followed by Pinus. Both reached altitudes around 1000 m very rapidly, perhaps via the eastern lowlands. Deciduous trees entered southern Setesdal early in the Holocene, reaching their greatest abundance at Dalane where they dominated the forests throughout the Holocene; the site today is in the Nemoral vegetation zone (Moen 1998). Deciduous trees were never so abundant at Grostjørna, with their greatest expansion c. 9000–5500 cal b.p. and the forests were dominated by Pinus and Betula throughout the Holocene. Today the site is in the Southern-Boreal zone of Moen (1998) and the surrounding forests are dominated by Pinus with deciduous trees in locally favourable habitats, such as south-facing cliffs. At 1000 m open birch- and pine-forest occurred around Lille Kjelavatn until climate cooling resulted in the loss of pine c. 4000 cal b.p. Betula gradually decreased and it is at its climatic limit there today. 150 m higher, birch was the only tree present in any abundance in the early Holocene up to c. 5000 cal b.p., after which the woodland became open and Northern-Boreal in character. By about 2500 cal b.p., the tree-line had descended below Holebudalen and it became surrounded by low-alpine vegetation, a situation that has continued until today.
Evidence for human activity is relatively slight at all the sites. In the lowlands grazing probably occurred since c. 5000 cal b.p., but appreciable cereal cultivation is only registered after c. 1500 cal b.p. Southern Setesdal is unsuitable for large-scale farming and today it is still largely forested. The mountains were used for summer grazing for at least 5000 years (Plantago lanceolata pollen) and this activity, combined with tree felling, may have been a factor in the reduction of high altitude forests, in addition to climate cooling.
Conclusions
The main trends in the vegetation development at the four sites throughout the Holocene are summarised in Fig. 7. At all sites except Lille Kjelavatn (where the sequence only covers 9000 years), pioneer vegetation on mineral soils became denser and humus content increased. A temperature maximum during the first half of the Holocene was followed by a cooling from around 5000 cal b.p. that is especially well demonstrated through the changes in the tree-lines and tree-limits at Holebudalen and Lille Kjelavatn.
At all sites the pioneers Betula and Pinus arrived first. At Dalane and Grostjørna, temperature increase allowed the establishment of Alnus and Corylus. The progressive increase of temperature and maturation of soil through time facilitated the growth of broad-leaved trees such as Ulmus and Tilia, which are also better competitors for abiotic factors such as light. The establishment of Quercus was not restricted by temperature, since it has similar temperature demands to Tilia. Most likely, gradual soil acidification and consequently reduced competition allowed Quercus to become established locally. The pollen records of Picea demonstrate its spread from the east into upper Setesdal and its late and rather ineffective spread into southern Setesdal. It is absent naturally from the middle of the valley today.
Macrofossil concentration and pollen influx for trees generally correspond well at the lowland sites, Dalane and Grostjørna, except for Corylus, Fraxinus and, Ulmus, where the macrofossil evidence is unreliable, due to poor representation and/or preservation. The pollen influx and percentage curves show a striking lack of variation through the Holocene for most of the tree taxa. The macrofossil record is more precise for well-represented taxa, showing local arrivals and disappearances and changes in population densities. At Lille Kjelavatn and Holebudalen the pollen record of Betula would be impossible to interpret without the macrofossil record. In addition, it is possible to separate dwarf- and tree-birch macrofossils. Pinus is well represented by pollen and the record of stomata is valuable in forested sites. Pollen percentages are misleading at the high-altitude sites with low local pollen productivity. The interpretation of the history of pine at Lille Kjelavatn (1000 m) depends entirely on the macrofossil record. Alnus pollen and macrofossil records generally correspond well in the lowlands. However, Alnus pollen is present at sites that were always above the altitudinal limit of Alnus spp.
The pollen and macrofossil data provide both contrasting and complementary views of vegetational change in the Setesdal valley during the Holocene. The contrasts arise from the fact that the two proxies record the vegetation composition at different spatial scales. In addition, different production, dispersal and preservation of pollen and macrofossils, and different site characteristics influencing taphonomy differentiate the records. Because macrofossils are derived from areas near the lake, they can reflect vegetation patterns at the scale of the altitudinal gradient. In contrast, much of the pollen collected in the lake sediments comes from greater distances and hence it integrates the forest composition over a broad area encompassing a wide range of elevations. In order to track tree movements during the Holocene over an altitudinal gradient covering a relatively small spatial distance, macrofossil evidence of local tree presence is essential (see also Jackson and Whitehead 1991; Barnekow 1999; Wick and Tinner 1997; Tinner and Theurillat 2003). The inherent problems with sediment dating and with both pollen analysis and macrofossil analyses are highlighted in the Setesdal study. General climatic conclusions can be drawn, but a denser concentration of well-dated sediment sequences is probably needed to test hypotheses of migration lags and rates of vegetational response to climate change.
References
Aas B, Faarlund T (1988) Postglacial forest limits in central south Norwegian mountains. Radiocarbon dating of subfossil pine and birch stumps. Norsk Geografisk Tidsskrift 42:25–61
Aas B, Faarlund T (1996) The present and past Holocene subalpine birch belt in Norway. Paläoklimaforchung 20:19–42
Aas B, Faarlund T (1999) Macrofossils versus pollen as evidence of the Holocene forest development in Fennoscandia. AmS-Rapport Stavanger 12B:307–346
Allen JRM, Huntley B (1999) Estimating past floristic diversity in montane regions from macrofossil assemblages. Journal of Biogeography 26:55–73
Andersen ST (1966) Interglacial vegetation succession and lake development in Denmark. The Palaeobotanist 15:117–127
Aune B (1993) Nasjonalatlas for Norge, kartblad 3.1.6. (National Atlas of Norway, sheet 3.1.6) Statens kartverk
Barnekow L (1999) Holocene tree-line dynamics and inferred climatic changes in the Abisko area, northern Sweden, based on macrofossil and pollen records. The Holocene 9:253–265
Bennett KD (1996) Determination of the number of zones in biostratigraphical sequences. New Phytologist 132:155–170
Bennett KD, Birks HJB (1990) Postglacial history of alder (Alnus glutinosa (L.) Gaertn.) in the British Isles. Journal of Quaternary Science 5:123–133
Berglund BE (1966) Late-Quaternary vegetation in Eastern Blekinge, south-eastern Sweden. II. Post-Glacial time. Opera Botanica 12:1–190
Berglund BE, Ralska-Jasiewiczowa M (1986) Pollen analysis and pollen diagrams. In: Berglund BE (ed) Handbook of Holocene Palaeoecology and Palaeohydrology. Wiley, Chichester, pp 455–484
Birks HH (1973) Modern macrofossil assemblages in lake sediments in Minnesota. In: Birks HJB, West RG (eds) Quaternary plant ecology. Blackwell Scientific Publications, Oxford, pp 173–190
Birks HH (2001) Plant macrofossils. In: Smol JP, Birks HJB, Last WM (eds) Tracking environmental change using lake sediments, vol 3. Kluwer, Dordrecht, pp 49–74
Birks HH (2003) The importance of plant macrofossils in the reconstruction of Late-Glacial vegetation and climate: examples from Scotland, western Norway, and Minnesota, USA. Quaternary Science Reviews 22:453–473
Birks HH, Birks HJB (2000) Future uses of pollen analysis must include plant macro-fossils. Journal of Biogeography 27:31–35
Birks HH, Birks HJB (2003) Reconstructing Holocene climates from pollen and plant macrofossils. In: Mackay A, Battarbee R, Birks J, Oldfeld F (eds) Global change in the Holocene. Arnold, London, pp 342–357
Birks HH, Larsen E, Birks HJB (2005) Did tree-Betula, Pinus, and Picea survive the last glaciation along the west coast of Norway? A review of the evidence, in light of Kullman (2002). Journal of Biogeography 32:1461–1471
Birks HH, Vorren KD, Birks HJB (1996) Holocene treelines, dendrochronology and palaeoclimate. Paläoklimaforschung 20:1–18
Birks HJB (1973) Past and Present Vegetation of the Isle of Skye: a Palaeoecological Study. Cambridge University Press, London
Birks HJB (1986) Late-Quaternary biotic changes in terrestrial and lacustrine environments, with particular reference to north-west Europe. In: Berglund BE (ed) Handbook of holocene palaeoecology and palaeohydrology. Wiley, Chichester, pp 3–65
Birks HJB (1989) Holocene isochrone maps and patterns of tree-spreading in the British Isles. Journal of Biogeography 16:503–540
Birks HJB, Gordon AD (1985) Numerical methods in Quaternary pollen analysis. Academic Press, London
Carrión JS (2002) Patterns and processes of Late Quaternary environmental change in a montane region of southwestern Europe. Quaternary Science Reviews 21:2047–2066
Dahl E (1998) The Phytogeography of Northern Europe (British Isles, Fennoscandia and adjacent areas). Cambridge University Press, Cambridge
Dahl SO, Nesje A (1996) A new approach to calculating Holocene winter precipitation by combining glacier equilibrium-line altitudes and pine-tree limits: a case study from Hardangerjøkulen, central Norway. The Holocene 6:381–398
Danielsen A (1970) Pollen-analytical Late Quaternary studies in the Ra district of Østfold, southeast Norway. Årbok Univ. Bergen 1970, Matematisk-Naturvitenskapelig Serie 14
Dinter M van, Birks HH (1996) Distinguishing fossil Betula nana and B. pubescens using their wingless fruits: Implications for the late-glacial vegetational history of western Norway. Vegetation History and Archaeobotany 5:229–240
Dunwiddie PW (1987) Macrofossil and pollen representation of coniferous trees in modern sediments from Washington. Ecology 68:1–11
Eide W (2003) Plant macrofossils as a terrestrial climate archive for the last 11000 years in south and central Norway. Doctoral Thesis University of Bergen
Førland E (1993) Nasjonalatlas for Norge, kartblad 3.1.1 (National Atlas for Norway, sheet 3.1.1). Statens kartverk
Geisecke T, Bennett KD (2004) The Holocene spread of Picea abies (L.) Karst. in Fennoscandia and adjacent areas. Journal of Biogeography 31:1523–1548
Grimm EC (1990) TILIA and TILIA.GRAPH. PC spreadsheet and graphics software for pollen data. INQUA, Working Group on Data-handling Methods Newsletter 4:5–7
Hafsten U (1987) Vegetasjon, klima og landskapsutvikling i Trøndelag etter siste istid (Vegetation, climate, and landscape development in Trøndelag since the last glaciation). Norsk Geografisk Tidsskrift 41:144–163
Hafsten U, Henningsmoen KE, Hoeg HI (1979) Innvandring av gran til Norge (Immigration of spruce into Norway). Norsk Geografisk Tidsskrift 41:101–120
Heegaard E, Birks HJB, Telford RJ (2005) Relationships between calibrated ages and depth in stratigraphical sequences: an estimation procedure by mixed-effect regression. The Holocene 15:612–618
Helland A (1912) Trægrendser og sommervarmen (Tree-lines and summer temperatures). Grøndahl og Søns Boktrykkeri, Kristiania
Hicks S (1993) The use of recent pollen rain records in investigating natural and anthropogenic changes in the polar tree limit in Northern Fennoscandia. Paläoklimaforschung 9:5–18
Hicks S (1994) Present and past pollen records of Lapland forests. Review of Palaeobotany and Palynology 82:17–35
Hicks S, Hyvärinen H (1999) Pollen influx values measured in different sedimentary environments and their palaeoecological implications. Grana 38:228–242
Huntley B, Birks HJB (1983) An Atlas of Past and Present Pollen Maps for Europe: 0–13000 years ago. Cambridge University Press, Cambridge
Iversen J (1973) The development of Denmark's nature since the last glacial. Danmarks Geologiske Undersøgelse V, 7-C
Jackson ST (1989) Postglacial vegetational changes along an elevational gradient in the Adirondack Mountains (New York). New York State Museum Bulletin 465
Jackson ST, Whitehead DR (1991) Holocene vegetation patterns in the Adirondack Mountains. Ecology 72:641–653
Jensen HA (1985) Catalogue of late- and post-glacial macrofossils of Spermatophyta from Denmark, Schleswig Holstein, Scania, Halland, and Blekinge dated 13 000 b.p. to 1536 a.d.. Danmarks Geologiske Undersøgelse, Series A, 6
Juggins S (1991) Zone - version 1.2. Unpublished computer program: http://www.campus.ncl.ac.uk/staff/Stephen.Juggins/software/softhome.htm. University of Newcastle, UK
Juggins S (2002) Palaeo Data Plotter version 1.0 ©Steve Juggins, University of Newcastle, UK
Kaland PE (1984) Holocene shore displacement and shorelines in Hordaland, western Norway. Boreas 13:203–242
Kaland PE, Krzywinski K (1978) Hasselens innvandring etter siste istid og den eldre kystbefolkning (The immigration of hazel since the last glaciation and the old coast people). Arkeo: arkeologiske meddelelser fra Historisk Museum, Universitetet i Bergen 1:11–14
Karlén W (1998) Climate variations and the enhanced Greenhouse Effect. Ambio 27:270–274
Körner C (1998) A re-assesment of high elevation treeline positions and their explanation. Oecologia 115:445–459
Kremenetski CV, Sulerzhitsky LD, Hantemirov R (1998) Holocene history of the northern range limits of some trees and shrubs in Russia. Arctic and Alpine Research 30:317–333
Kullman L (1983) Past and present tree-lines of different species in the Händolan valley, central Sweden. In: Morisset P, Payette S (eds) Treeline ecology. Collection Nordicana 47, pp 24–45
Kullman L (1995) Holocene tree-limits and climate history from the Scandes Mountains, Sweden. Ecology 76:2490–2502
Kullman L (1998) The occurrence of thermophilous trees in the Scandes Mountains during the early Holocene: evidence for a diverse tree flora from macroscopic remains. Journal of Ecology 86:421–428
Kullman L (2000) The geoecological history of Picea abies in northern Sweden and adjacent parts of Norway. A contrarian hypothesis of postglacial tree immigration patterns. Geo-Öko 21:141–172
Kullman L (2002a) Rapid recent range-margin rise of tree and shrub species in the Swedish Scandes. Journal of Ecology 90:68–77
Kullman L (2002b) Boreal tree taxa in the central Scandes during the Late-Glacial: implications for Late-Quaternary forest history. Journal of Biogeography 29:111–127
Lid J, Lid DT (1994) Norsk flora. 6.utg. (Norwegian Flora, 6 ed) Det norske Samlaget, Oslo
Liedberg Jönsson B (1988) The Late Weichselian macrofossil flora in western Skåne, southern Sweden. LUNDQA Thesis 24
McVean DM (1953) Biological Flora of the British Isles. Alnus glutinosa (L.) Gaertn. Journal of Ecology 43:447–466
MacDonald GM, Edwards TWD, Moser KA, Pienitz R, Smol JP (1993) Rapid response of treeline vegetation and lakes to past climate warming. Nature 361:243–46
Moe D (1979) Tregrense-fluktuasjoner på Hardangervidda etter siste istid (Tree-line fluctuations on Hardangervidda since the last glaciation). In: Nydal R, Westin S, Hafsten U, Gulliksen S (eds) Fortiden i søkelyset. Datering med14C metoden gjennom 25 år. NTH, Trondheim, pp 199–208
Moe D (1998) Pollen production of Alnus incana at its south Norwegian altitudinal ecotone. Preliminary observations. Grana 37:35–39
Moe D, Odland A (1992) A palaeo- and recent-ecological discussion of Alnus incana in Norway. Acta Botanica Fennica 144:35–49
Moen A (1998) Nasjonalatlas for Noreg: Vegetasjon. (National Atlas for Norway: Vegetation). Statens Kartverk, Hønefoss
Molau U, Larsson E-L (2000) Seed rain and seed bank along an alpine altitudinal gradient in Swedish Lapland. Candian Journal of Botany 78:728–747
Moore PD, Webb JA, Collinson ME (1991) Pollen analysis, 2nd edn. Blackwell Scientific Publications, Oxford
Nesje A, Dahl SO, Løvlie R, Sulebak JR (1994) Holocene glacier activity at the southwestern part of Hardangerjøkulen, central-southern Norway: evidence from lacustrine sediments. The Holocene 4:377–382
Segerström U, von Stedingk H (2003) Early Holocene spruce, Picea abies (L.) Karst., in west central Sweden as revealed by pollen analysis. The Holocene 13:97–906
Seppä H, Birks HJB (2001) July mean temperature and annual precipitation trends during the Holocene in the Fennoscandian tree-line area: pollen-based climate reconstructions. The Holocene 11:27–539
Stuiver M, Reimer PJ (1993) Extended14C data base and revised CALIB 3.214C age calibration program. Radiocarbon 35:15–230
Stuiver M, Reimer PJ, Bard E, Beck JW, Burr GS, Hughen KA, Kromer B, McCormac G, van der Plicht J, Spurk M (1998) INTCAL98 radiocarbon age calibration, 24,000-0 cal b.p. Radiocarbon 40:527–539
Tallantire PA (1974) The palaeohistory of the grey alder (Alnus incana (L.) Moench.) and black alder (A. glutinosa (L.) Gaertn.) in Fennoscandia. New Phytologist 73:529–546
Telford RJ, Heegaard E, Birks HJB (2004) All age-depth models are wrong: but how badly? Quaternary Science Reviews 23:1–5
Tinner W, Theurillat J-M (2003) Uppermost limit, extent, and fluctuations of the timberline and treeline ecocline in the Swiss Central Alps during the past 11,500 years. Arctic, Antarctic, and Alpine Research 35:158–169
Tomlinson C (1985) An aid to identification of fossil buds, bud-scales and catkin-bracts of British trees and shrubs. Circaea 3:45–130
Troels-Smith J (1955) Karakterisering av løse jordarter (Characterisation of unconsolidated sediments). Danmarks Geologiske Undersøgelse, IV 3:1–73
Törnqvist TE, de Jong AFM, Oosterbaan WA, van der Borg K (1992) Accurate dating of organic deposits by AMS14C measurement of macrofossils. Radiocarbon 34:566–577
Velle G, Larsen J, Eide W, Peglar SM, Birks HJB (2005) Holocene environmental history and climate of Råtåsjoen, a low-alpine lake in south-central Norway. Journal of Paleolimnology 33:129–153
Wainman N, Mathewes RW (1990) Forest history of the last 12 000 years based on plant macrofossil analysis of sediment from the Marion Lake, southwestern British Columbia. Canadian Journal of Botany 65:2179–2187
Watts WA (1979) Late-Quaternary vegetation of central Appalachia and the New Jersey coastal plain. Ecological Monographs 49:427–269
Wick L, Tinner W (1997) Vegetation changes and timberline fluctuations in the central Alps as indicators of Holocene climatic oscillations. Arctic and Alpine Research 29:445–458
Wright HE (1980) Cores of soft lake sediments Boreas 9:107–114
Wright HE (1991) Coring tips. Journal of Paleolimnology 6:37–49
Acknowledgements
We thank L. Allott and T.E.H. Allott, C. Dalton, A. Furnes, P.E. Noon and E. Shilland for help with the fieldwork, E. Heegaard for deriving the age-depth models and A. Odland for the modern climate values. We appreciate comments on an earlier draft of this manuscript by J. Larsen and reviews by B. Aaby and L. Kullman. W. Eide was supported by VISTA (Climate archives 6520), N.H. Bigelow by NSF-NATO Postdoctoral fellowship DGE-9633851, S.M. Peglar by NFR (grant no. 110486/720) and H.H. Birks by NFR (NORPAST and NORPEC projects).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Eide, W., Birks, H.H., Bigelow, N.H. et al. Holocene forest development along the Setesdal valley, southern Norway, reconstructed from macrofossil and pollen evidence. Veget Hist Archaeobot 15, 65–85 (2006). https://doi.org/10.1007/s00334-005-0025-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-005-0025-7