
Abstract
The Magellan plunderfish Harpagifer bispinis is a small and benthic sub-Antarctic notothenioid commonly found in shallow waters of the Beagle Channel, whose biology is poorly known. Based on a seasonal sampling conducted throughout a year, we provide original data on some biological parameters, such as reproductive traits and age structure. The sampled population of Magellan plunderfish was sex balanced, showing no dimorphism in fish size and isometric body growth. Macroscopic and histological analyses of gonads enabled us to infer that this species is a total spawner, with a single annual spawning season during the austral winter. Total fecundity was considerably low (mean of 574 eggs per female), even compared with other harpagiferids. Based on daily increment counts made in juveniles, larval hatching took place between late September and early October, likely enhancing their survival during the warmer months. The Von Bertalanffy growth parameters, estimated from the length-at-age data pairs for the whole population, were L∞ = 80.7 (± 0.82), K = 0.25 (± 0.01), and t0 = − 2.31. The estimated growth performance (3.21) and life span (7 years) of H. bispinis were both comparable to those reported for their most closely related species, H. antarcticus. The life-history traits of Magellan plunderfish are discussed from an ecological and evolutionary perspective, through a comparative approach with their Antarctic counterparts.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Harpagiferidae is a monophyletic family (Derome et al. 2002; Near and Cheng 2008) that includes a single genus (Harpagifer) and between 10 and 12 species according to different authors (Duhamel et al. 2014; Eastman and Eakin 2016; Eschmeyer et al. 2019). Harpagiferids are the smallest fish among notothenioids ranging between 5.7 and 11 cm of total length (Eastman 2019), and are distributed in shallow waters between 0 and 320 m depth (Eastman 2017). They are ecologically and morphologically similar species (Eastman 2005) that inhabit marine benthic/epibenthic environments at both sides of the Antarctic Polar Front (APF). However, most of the species are distributed around the sub-Antarctic islands, whereas a single species inhabits the inshore waters off the Antarctic Peninsula (Harpagifer antarcticus Nybelin, 1947) and another species lives at the southern tip of South America (Harpagifer bispinis Forster, 1801).
From an evolutionary point of view, Harpagifer species provided new evidence of faunal connections between the Antarctic and South America through the Scotia Arc during the Pleistocene (between 1.7 and 0.8 Ma) (Hüne et al. 2015). These authors proposed that the northward displacement of the APF occurring during the middle Pleistocene, along with the long larval life span of H. antarcticus (Kock and Kellermann 1991; White and Burren 1992), and the retention of antifreeze glycoproteins facilitated their dispersion from polar latitudes. The next relocation of the APF to its current position probably finished the dispersion process and resulted in the definitive separation of H. bispinis from their closest ancestor H. antarcticus (Hüne et al. 2015).
The Magellan plunderfish H. bispinis inhabits the sublittoral zones and intertidal pools in southern South America (Hureau 1990) between the 46° 40′ S in the Pacific Ocean (Zama and Cardenas 1984) and 47° 45′ S in the Atlantic Ocean (Gostonyi pers. comm.). In these habitats, they can be found in the subtidal zone, within the kelp forest hidden on the floor beside the fronds and near the holdfast of Macrocystis pyrifera (Cruz-Jiménez 2019). Furthermore, Bruno et al. (2018) demonstrated the importance of the giant kelp forests for the early stages of this species in the Beagle Channel. Also, H. bispinis is commonly found under rocks in the low and medium levels of the intertidal zone, in tide pools. Living in these tides pools was interpreted as a huge ecophysiological challenge, as this habitat is usually frozen during winter and is exposed to a wide range of salinity (from 0 to 38 ‰) by the flow of small freshwater input during ice melting (Calvo, unpublished data). The capacity of H. bispinis to cope with these ranges of salinity and temperature are related to specific adaptation such as aglomerularism in the kidney (Perez et al. 2003) and the retention of antifreeze glycoproteins (DeVries pers. comm.).
Biological information on H. bispinis is still very scarce. From a morphological point of view, this species is characterized by having a big head with two prominent preopercular and opercular spines and a mouth with villiform teeth (Lloris and Rucabado 1991). This is a sluggish, bottom-dweller species with the lowest value of buoyancy among sub-Antarctic notothenioids (Fernández et al. 2012) that can change their scaleless-skin colour depending on the type of substrate. It was described as a “sit-and-wait” predator with a mean trophic level of 3.7 (Riccialdelli et al. 2017), primarily consuming benthic prey such as crustacean gammarids, tanaids and isopods (Hüne and Vega 2016).
Some generalization on reproductive traits and early life stages of harpagiferids has been made, although based almost exclusively on H. antarcticus (Everson 1968; Tomo and Cordero Funes 1972; Daniels 1978, 1983; Burren 1988; Kellerman 1990; White and Burren 1992; La Mesa et al. 2017). For example, it was demonstrated that H. antarcticus exhibits nesting behaviour, consistently with its relatively low fecundity. Furthermore, it is characterized by a wide spawning period, demersal eggs, small size at hatch and an extended larval pelagic phase. In a recent study carried out in the South Shetland Islands, the hatching of small and fast growing larvae of H. antarcticus occurred in spring–summer and is probably triggered by wind-induced turbulence in nearshore subtidal environments (La Mesa et al. 2017). Conversely, little is known about spawning season, fecundity, hatching time and size at maturity in Harpagifer species living in sub-Antarctic waters. Until now, the oogenetic process of Harpagifer spinosus was described from the Crozet Islands (Van der Molen and Matallanas 2004) and different larval stages were described for H. kerguelensis (Koubbi et al. 1991) and H. georgianus (North 1991).
Considering the wide distribution of harpagiferids from the Antarctic Peninsula to sub-Antarctic Islands to Patagonian coastal waters, it would be interesting to assess how environmental factors in different latitudes contribute to shape the life strategies of these species. In particular, our hypothesis is that the establishment of closely related species in non-Antarctic coastal waters, such as H. bispinis, supposes a rearrangement of their life-history traits to cope with seawater temperature increases of up to 11 °C, as is the case in the Beagle Channel during summer. To address this issue, the aim of this study was to analyse the life-history strategies of H. bispinis including its reproductive cycle, age structure and growth rate, in order to gain further insight into the biological adaptations evolved in non-Antarctic notothenioids.
Materials and methods
Study area
The Beagle Channel (BC) is a tectonic valley repeatedly carved by glacial cycles and flooded by the sea during the last transgression (Rabassa 2008; Bujalesky 2011). The BC is a unique subpolar marine ecosystem situated at the tip of South America with water flowing eastwards and connecting the Pacific and the Atlantic Oceans (Isla et al. 1999; Gordillo et al. 2008). In this area, water temperature ranges between 4.2 and 4.3 °C in August and between 8.9 and 9.8 °C in January, while salinity ranges from 26.7‰ in November to 31.3‰ during July (Balestrini et al. 1998). However, extreme values up to 0 °C during winter and 11 °C in summer as well as a salinity range between 0 and 38‰ can be found in the intertidal zone. The tidal regime is almost microtidal, with typical amplitudes between 1 and 2 m (D’Onofrio 1989).
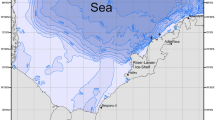
Fish samples were collected in an intertidal zone with rocky substratum near the southernmost natural protected area in Argentina, Tierra del Fuego National Park (Fig. 1). This protected area supports high marine diversity, representing the spawning and nursery site for many invertebrate species (Rae and Calvo 1995; Adami and Gordillo 1999; Diez et al. 2009). The BC fish fauna encompasses almost 50 different species (Lloris and Rucabado 1991). Fish assemblages in the kelp forest of the BC are dominated in diversity and biomass by notothenioids both as adults (Vanella et al. 2007) and at larvae stages (Bruno et al. 2018). In the intertidal zone, the most common species are Patagonotothen tessellata (Richardson, 1845), P. cornucola (Richardson, 1844), P. sima (Richardson, 1845) and H. bispinis.

Map of South America (upper left) and fish sampling site in the Beagle Channel (main panel)
Field sampling and laboratory activities
Fish were collected by hand during low tides both during the day and night at the same beach during spring (27 November 2018), summer (18 February 2019), autumn (30 April 2019) and winter (18 and 29 July 2019) to consider seasonal variability. Fish were transported to the laboratory in aerated 100 L tanks with water at environmental temperature. In the laboratory, fish were euthanized with tricaine methanesulfonate (MS-222, Sigma- Aldrich, St. Louis, MO, USA). The total length (TL, mm) was measured and the total body mass (TW, g) and gonad mass (GW, g) were recorded with an accuracy of 0.01 g. The sagittal otoliths were removed and stored dry. Gonads were extracted and immersed in Bouin solution for 12 h and then in ethanol 70% for histological analyses. Sex was assessed macroscopically by the gross gonad morphology.
For ageing purposes, left or right otoliths were randomly selected from a few fish, and after being sectioned, their ring pattern was observed. This pattern was compared with the result from whole otolith reading. As no difference was found, otolith pairs from each individual were put whole in a petri dish and analysed under reflected light by means of a stereomicroscope (Leica M205C) at ×25 magnification. To enhance the contrast between opaque and translucent zones, otoliths were soaked in ethanol before reading. Individual age was estimated by counting annuli, which are formed by contiguous opaque and translucent zones assumed to be laid down annually. In a few numbers of unsexed fishes smaller than 40 mm, sagittal otoliths were embedded in resin (Crystalbond 509), ground on abrasive paper and polished with a lapping film. The sagittal sections were observed at ×400–630 magnification using a light microscope (LEICA DM400B) linked to a software video analysis system (LAS, Leica Application Suite). Individual age (days) was estimated by microincrement counts, assuming they were laid down on a daily basis (White 1991).
Histological analyses of gonads were performed in males and females collected in all seasons investigated. Following a standard protocol, gonads were dehydrated through increasing concentrations of ethanol and embedded in paraffin. Transverse sections of 6–8 µm thickness were put on slides and stained with Groat’s haematoxylin–eosin. Gonad sections were examined using a light microscope (Leica DM4000B) at ×5–100 magnification. According to Brown-Peterson et al. (2011), gonad development of adult males and females was assigned to four different phases based on cell morphology, as developing, spawning capable, regressing and regenerating.
Data analysis
The length–frequency distributions of sexes were compared by the Kolmogorov–Smirnov two-sample test. The sex ratio was estimated for the whole sampling period and the difference from 1:1 was analysed by the Chi-square test (Zar 1999). The length–weight relationships of fish were calculated for each sex, fitting data to the equation log (TW) = log a + b (TL), where a and b are the regression parameters being estimated by the least-squares method. The slopes of regressions (b) of both sexes were tested for allometry (H0: b = 3), applying a t test to the equation t = (b – 3) SE−1, where SE is the standard error of b, and the difference between them tested using an F test (Zar 1999). All tests were performed with a significance level of 0.05.
The gonadosomatic index (GSI), as the percentage of GW to TW, was calculated for each specimen. To assess the reproductive investment during the gonad cycle, the mean value (± standard deviation SD) of GSI was calculated for each sampling season and for each phase of gonad development separately from males and females. The ovaries of spawning capable females were examined and the total number of yolked oocytes was counted to estimate individual total fecundity (Ftot). The relative fecundity (Frel) was then calculated as the number of yolked oocytes per gram of total body mass.
For assessing ageing precision between the two readers, the average percent error (APE) and the mean coefficient of variation (CV) (Campana 2001) were applied. A bias plot was computed to measure for systematic difference between readings (Campana et al. 1995). The somatic growth rate of fish population was modelled by fitting the von Bertalanffy growth function (VBGF) to the individual age-length data pairs. The VBGF parameters (L∞, K, t0) estimated for males and females were then compared using the likelihood ratio test (Kimura 1980). The growth performance index (Φ′ = 2 logL∞ + logK) was calculated to compare with other notothenioids.
Results
Demographic characteristics
The sampled population of H. bispinis consisted of 99 females ranging between 38 and 76 mm TL and between 0.79 and 7.01 g TW and of 84 males ranging between 40 and 80 mm TL and between 1.19 and 8.28 g TW. A few specimens (n = 14) were unsexed juveniles between 38 and 43 mm TL and between 0.71 and 1.08 g. No statistical difference was found between the length–frequency distributions of males and females (Kolmogorov–Smirnov test, D = 0.07, p > 0.05) (Fig. 2). The departure of sex ratio from the expected proportion 1:1 was not statistically significant (Chi-square test, X1 = 1.23, p = 0.27). As no difference was found between sexes (Fisher test, F1,178 = 0.95, p = 0.33), the relationship between TL and TW was calculated for the whole population (Fig. 3), resulting in the following function: TW = 0.0000237 TL2.88. The population exhibited an isometric growth since the slope (b) was non-significantly different from 3 (Student test, t180 = 1.47, p = 0.14).

Length–frequency distribution of Harpagifer bispinis males and females collected in the Beagle Channel

Length–mass relationship for the population (sexes pooled) of Harpagifer bispinis in the Beagle Channel
Reproductive traits
As a measure of the reproductive investment and the gonad development by sex, the GSI ranged from 0.3 to 6.8% in males and from 0.4 to 42.9% in females. Although with difference in amplitude, the GSI trend according to histological development phases was the same in males and females. GSI increased from regenerating through developing to spawning capable phases, with a sharp decrease after spawning (regressing phase) (Fig. 4). Accordingly, the seasonal trend of GSI showed a minimum in November and then increased progressively from April to July when H. bispinis was ready to spawn (Fig. 5). Gonad development was rather synchronous, with almost all specimens at the same phase in the same period (Fig. 6).

Mean values of gonadosomatic index (GSI) calculated for each histological phase of gonad development in males and females of Harpagifer bispinis. Vertical bars represent the standard deviation

Mean values of gonadosomatic index (GSI) calculated for males and females of Harpagifer bispinis per sampling period. Vertical bars represent the standard deviation

Pie charts showing the proportion of fish in each stage of gonad development calculated for females (left panel) and males (right panel) of Harpagifer bispinis during the whole sampling period
In spring (November), males and females were in the regressing (cessation of spawning) and regenerating phases (sexually mature but reproductively inactive) of gonad development, respectively. Males had small and flaccid testes, with a proliferation of spermatogonia in the peripheral lobules and a small amount of residual spermatozoa in the enlarged lumina of inner lobules (Fig. 7a). Females were characterized by small ovaries, with ovarian follicles composed of primary growth oocytes (chromatin nucleolar and perinucleolar stages, mean size ± SD, 51.7 ± 9.7 µm and 92.6 ± 17.8 µm, respectively) and cortical alveoli at an early stage (260.9 ± 22.3 µm), a thick ovarian wall and a few scattered degenerating postovulatory follicles (POF) (Fig. 7b).

Gonad histological sections of Harpagifer bispinis collected in the Beagle Channel. a Regressing male; b regenerating female; c regenerating male; d early developing female; e developing male; f developing female; g spawning capable male; h, i spawning capable female; l regressing female. Ao atretic oocyte, Ca cortical alveoli oocyte, Gc granulosa cell layer, Ow ovarian wall, Pg primary growth oocyte, Pof postovulatory follicle, Sc spermatocyte, Sg spermatogonia, St spermatid, Sz spermatozoa, Vtg vitellogenic oocyte, yg yolk granules, Zr zona radiata
In summer (February), males were in the regenerating phase, whereas females were in early developing subphase of gonad development. Males had small and compact testes, with an active proliferation of spermatogonia throughout the lobules and lumina completely occluded (Fig. 7c). Gonads of females increased in size, and the ovarian follicles mainly consisted of cortical alveoli oocytes (386.4 ± 29.1 µm) with cytoplasm completely filled by empty vesicles, as well as a few scattered primary growth oocytes (Fig. 7d).
In autumn (April), males and females were both in the developing phase (i.e. gonads beginning to develop, but still not ready to spawn) of gonad development. In males, testes were enlarged with lobules mainly containing cysts of both spermatocytes and spermatids, as well as spermatogonia in the periphery (Fig. 7e). In females, ovaries mainly consisted of vitellogenic oocytes (572.9 ± 20.9 µm), characterized by yolk vesicles of different sizes filling completely the cytoplasm from the central nucleus towards the periphery, and a few scattered primary growth oocytes (Fig. 7f).
In winter (July), all males were in the spawning capable phase (i.e. they are developmentally and physiologically able to spawn in the current season), whereas females were in developing, spawning capable or regressing phases (cessation of spawning). Males had large gonads with lumina of lobules and spermiduct massively filled by spermatozoa, and a few spermatogonia present in the periphery of testes (Fig. 7g). Spawning capable females had ovaries entirely composed of late vitellogenic oocytes (1260.3 ± 58.3 µm), characterized by yolk granules starting to fuse together in a single yolk plate and a thick multilayered zona radiata surrounded by the granulosa cells (Fig. 7h, i). In regressing females, ovaries were flaccid and contained postovulatory follicle (POF) and atretic vitellogenic oocytes at various stages of resorption, as well as a few sparse primary growth oocytes (Fig. 7l).
The size frequency distribution of oocytes recorded in the ovaries of the spawning capable females revealed two well-separated groups (group synchronous development). The smaller group was composed of primary growth oocytes and the larger one consisted of yolked oocytes at the end of the vitellogenic process (Fig. 8). Total fecundity (Ftot) ranged between 477 and 732 eggs/female (534 ± 104), corresponding to a relative fecundity between 227 and 458 eggs/g (307 ± 77) for seven females between 61.6 and 71.5 mm TL caught in August.

Frequency distribution of oocyte diameter in the ovary of Harpagifer bispinis spawning capable females (n = 7)
Age and growth
Individual age was estimated by otolith microincrement counts in three unsexed specimens ranged between 210 and 220 days (Fig. 9a). The hatch-date, back-calculated by date of fish capture ranged between 22 September and 2 October. The life span was the same for males and females and ranged between 0.5 and 7 years (Fig. 9b). Ageing precision between readers was good (APE = 5.5%; CV = 7.7), and bias was not detected between readings (Fig. 10). The likelihood ratio test indicated that the von Bertalanffy growth parameters did not differ between sexes and thus a single VBGF was fitted to the H. bispinis population (Fig. 11). The estimated parameters and their ± 95% confidence interval were as follows: L∞ = 80.7 (± 0.82), K = 0.25 (± 0.01), t0 = − 2.31. The growth performance index was 3.21.

Micrographs of Harpagifer bispinis sagittal otoliths from the Beagle Channel. a Transverse section from an unsexed fish (38 mm TL), showing the microincrement pattern and the primordium (arrow); b whole otolith from a 4-year-old male (64 mm TL), showing the annulation pattern of translucent and opaque zones

Age bias plot between readers, showing mean age and the 95% confidence interval of second reader (error bars) for each age class assigned by the first reader

Von Bertalanffy growth curve fitted to the age-at-length data pairs estimated for the whole population of Harpagifer bispinis from the Beagle Channel
Discussion
This study represents the first attempt to provide insights into some biological aspects of the Magellan plunderfish Harpagifer bispinis inhabiting the inshore waters of the Beagle Channel. We focused our attention on the demographic structure of the population in terms of size and sex ratio, age and growth, reproductive effort and gonad cycle. Analysing present results in a comparative context within the genus Harpagifer, the majority of the biological information available elsewhere concerns almost exclusively H. antarcticus, the Antarctic counterpart of H. bispinis distributed off the Antarctic Peninsula and neighbouring islands north of it. The close phylogenetic relationship between the two species (Hüne et al. 2015), together with the different environments in which they live, allowed us to evaluate if and how environmental conditions might have contributed to shape their own life strategies. We also discuss latitudinal trends in specific biological parameters.
Demographic characteristics
In agreement with previous studies done on H. bispinis (Hüne and Vega 2016; Fernández et al. 2018), the Beagle Channel population is characterized by a balanced sex ratio. In addition, this species did not present sexual dimorphism in size and both sexes showed the same type of isometric body growth. The maximum size attainable by H. bispinis should be updated to 80 mm TL, a value slightly larger than the one previously reported by Eastman (2019). Consistently with a low level of genetic divergence, the demographic characteristics of H. bispinis are similar to those found in H. antarcticus, for which a balanced sex ratio and no sex difference in fish size, body growth, length at age and life span have been demonstrated (Daniels 1978, 1983; Tomo and Cordero Funes 1972; White and Burren 1992).
Reproductive traits
The sampling program enabled to follow the annual gonad cycle of H. bispinis in the Beagle Channel. Gonad maturation was synchronous in both sexes and the GSI increased from November to July when it reached a peak of 23 to 42% in spawning females. Spawning capable and regressing females were found at the end of July, indicating that the spawning season begins during mid-July and likely extents throughout August. The spawning period reported for H. antarcticus is also during the austral winter, taking place between May and July at Signy Island (White and Burren 1992) and during June–August off the Antarctic Peninsula (Daniels 1978).
The GSI at the spawning time is similar between females of H. bispinis in the Beagle Channel (Fig. 5) and of H. antarcticus (between 14 and 39%) off the Antarctic Peninsula (Tomo 1981). At Signy Island (South Orkney Islands), the maximum GSI recorded for spawning females of H. antarcticus was considerably higher (55–60%), but it was estimated using the eviscerated weight (White and Burren 1992).
In nototheniod fishes, there are considerable differences in the total fecundity among the families (Kock 1992). Within Harpagiferidae, Van der Molen and Matallanas (2004) estimated a total fecundity of 1227 for H. spinosus, while in H. antarcticus the total fecundity ranged from 1113 to 1522 (White and Burren 1992). Both values are considerably higher than the fecundity estimated for H. bispinis. In this family, the low number of eggs could be related to parental care and nesting behaviour along with small size (White and Burren 1992; Van der Molen and Matallanas 2004).
As commonly described in notothenioid fishes (Kock and Kellermann 1991), two clutches of oocytes were recorded in H. bispinis spawning females. The more advanced will be ovulated during the current spawning season whereas the smaller one represents the oocyte reserve for the next year. According to the synchronous gonad maturation process and the seasonal trend of GSI, H. bispinis can be considered a total spawner with a single annual spawning season. As a consequence, the hypothesis of a second spawning period during late summer previously suggested by some authors cannot be supported (Rae and Calvo 1995, 1996). However, another sub-Antarctic species (Harpagifer spinosus) from the Crozet Island has been described as a potential fractional spawner with separate ovulatory events (Van der Molen and Matallanas 2004).
Coupling the spawning season with the hatching time (between late September and early October), an incubation period of approximately two months can be inferred for H. bispinis in the Beagle Channel. This period is considerably lower than those reported for H. antarcticus, which ranged between 4 and 5 months at Signy Island (White and Burren 1992) and between 3 and 4 months off the Antarctic Peninsula (Daniels 1978). Since the hatching size of the two species is very similar (~ 5 mm) (La Mesa et al. 2017; Bruno, pers. comm.), differences in egg incubation time could be explained by the higher water temperatures in the Beagle Channel (4.9 °C in winter to 10.1 °C in summer, Almandoz et al. 2011) than in the Antarctic Peninsula (− 1.8 °C in winter to 1.8 °C in summer, Krebs 1983) and at Signy Island (− 1.8 °C in winter to 0.3 °C in summer, Clarke et al. 1988). In spite of this difference, hatching time and larval stage occur during spring and summer when better conditions exist in southern marine environments (Kellerman 1990; White and Burren 1992; La Mesa et al. 2017). In this regard, as an adaptation to maximize larval survival and increase the likelihood of success of juveniles during the next winter, both Harpagifer species tuned the larval occurrence to the seasonal phytoplankton blooms which trigger the food chain.
The kelp forests of Macrocystis pyrifera characterize the inshore waters of the Beagle Channel and represent a nursery ground for the early life stages of H. bispinis (Bruno et al. 2018). In proximity of our sampling site (Lapataia Bay 54° 51′ S-68° 34′ W), early larvae of H. bispinis between 5 and 10 mm TL were collected during August–October (Bruno, pers. comm.), supporting our estimates on hatching period. Despite the fact that it is not possible to provide a direct estimate of the larval period of H. bispinis, it should last for less than seven months, considering the back-calculated age range (210–220 days) of the smaller individuals already settled. Similarly, H. antarcticus exhibited a pelagic period lasting for about four months (Kellermann 1990; White and Burren 1992; La Mesa et al. 2017).
From an evolutionary point of view, the extent of the larval period has significant implications. The proposed mechanism of speciation between H. antarcticus and H. bispinis rests on three main argumentations: (i) the position of the ACC and the APF underwent major northward latitudinal shifts during the middle Pleistocene (0.9 Ma), providing suitable conditions for the colonization of South America from Antarctica; (ii) the antifreeze protection might have conferred H. bispinis the ability to cope with harsh habitat conditions and colonize unoccupied shoreline habitats during glacial maxima; and (iii) the long larval pelagic life reported in H. antarcticus might have promoted larval dispersal (Hüne et al. 2015). Nevertheless, special attention should be paid to the latter point, as low fecundity, nesting behaviour and parental care reported for H. antarcticus likely determine a low potential for larval dispersal (La Mesa et al. 2017). In addition, these authors suggested that gene flow through connectivity among populations of H. antarcticus from islands separated by deep oceanic waters might be considerably reduced or absent and thus promoting species isolation. The transport of adults and juveniles on floating kelp through the west wind drift represents an alternative explanation for the colonization of H. spinosus from the South Orkney to Crozet and Kerguelen Islands (Matallanas 1997).
Age and growth
In the Beagle Channel, H. bispinis attained a maximum size of 80 mm TL and a life span of 7 years. These parameters were slightly different when estimated through otolith reading for H. antarcticus off the Antarctic Peninsula, being, respectively, 95 mm SL and 9 years (Daniels 1983). As the von Bertalanffy growth parameters K and L∞ are inversely related, we calculated the growth performance index (Φ′) to compare the body growth of Harpagifer species. As a result, the Φ′ was 3.21 for H. bispinis and 3.18 and 3.19 for males and females of H. antarcticus, indicating no difference between them. Moreover, these values were at the lower end compared to the growth performance of high Antarctic notothenioid fishes that were recalculated using K and L∞ values provided by La Mesa and Vacchi (2001).
Final remarks
In conclusion, H. bispinis shares most of the common reproductive characteristics of notothenioids, such as iteroparity, group synchronous ovary development in maturing females, low and determinate fecundity and single annual spawning season (Kock and Kellermann 1991). By contrast, in H. bispinis the vitellogenesis lasts one year, whereas in most Antarctic notothenioids it is a biennial process. Compared to their Antarctic counterpart, H. bispinis inhabiting the Beagle Channel attained a smaller size and a relatively lower life span, as well as a shorter spawning period and egg incubation time. Along with Harpagifer marionensis and Harpagifer palliolatus, H. bispinis is the only species within the genus exclusively distributed north of the sub-Antarctic Polar Front. In allopatry, Antarctic and Patagonian populations of Harpagifer may have evolved some key adaptations under different environmental conditions. As an experimental example, when H. antarcticus was exposed to a seawater temperature and salinity values similar to those found in the Beagle Channel, this produced a negative impact on its energy budget and higher mortality rate (Navarro et al. 2019).
The ecological plasticity in terms of reproductive strategies, growth and feeding habits recorded in Harpagiferids at different localities (e.g. Daniels 1983; White and Burren 1992; Casaux 1998; Van der Molen and Matallanas 2004; Hüne and Vega 2016; La Mesa et al. 2017; present work) enabled these fishes to colonize the intertidal zone and the coastal waters of the Antarctic Peninsula, the sub-Antarctic islands and the Patagonian region.
References
Adami ML, Gordillo S (1999) Structure and dynamics of the biota associated with Macrocystis pyrifera (Phaeophyta) from the Beagle Channel, Tierra del Fuego. Sci Mar 63:183–191
Almandoz GO, Hernando MP, Ferreyra GA, Schloss IR, Ferrario ME (2011) Seasonal phytoplankton dynamics in extreme southern South America (Beagle Channel, Argentina). J Sea Res 66:47–57
Balestrini C, Manzella G, Lovrich GA (1998) Simulacion de corrientes en el Canal Beagle y Bahia Ushuaia, mediante un modelo bidimensional. Serv Hidrog Naval Dto Oceanog Inf Tec 98:1–58
Brown-Peterson NJ, Wyanski DM, Saborido-Rey F, Macewicz BJ, Lowerre-Barbieri SK (2011) A standardized terminology for describing reproductive development in fishes. Mar Coast Fish 3:52–70
Bruno DO, Victorio MF, Acha EM, Fernández DA (2018) Fish early life stages associated with giant kelp forests in sub-Antarctic coastal waters (Beagle Channel, Argentina). Polar Biol 41:365–375
Bujalesky G (2011) The Flood of the Beagle Channel (11,000 YRB.P.). Tierra del Fuego An Inst Patagon 39:5–21
Burren PJ (1988) Reproductive biology of Harpagifer sp. at Signy Island, South Orkney Islands. Unpubl Master's Thesis, University College of North Wales, pp 1–56
Campana SE (2001) Accuracy, precision and quality control in age determination, including a review of the use and abuse of age validation methods. J Fish Biol 59:197–242
Campana SE, Annand MC, McMillan JI (1995) Graphical and statistical methods for determining the consistency of age determinations. Trans Am Fish Soc 124:131–138
Casaux RJ (1998) The contrasting diet of Harpagifer antarcticus (Notothenioidei, Harpagiferidae) at two localities of the South Shetland Islands, Antarctica. Polar Biol 19:283–285
Clarke A, Holmes LJ, White MG (1988) The annual cycle of temperature, chlorophyll and major nutrients at Signy Island, South Orkney Islands, 1969–82. Br Antarct Surv Bull 80:65–86
Cruz-Jiménez AM (2019) Ensambles de peces en los bosques de kelp de Macrocystis pyrifera en el Canal Beagle, Tierra del Fuego: estructura comunitaria y variación espacio-temporal. Unpubl PhD Thesis, Universidad Nacional de La Plata, Argentina, pp 1–193
D’Onofrio E, Orsi A, Locarnini R (1989) Estudio de marea en la costa de Tierra del Fuego. Serv Hidrog Naval Dto Oceanog Inf Tec 49:1–81
Daniels RA (1978) Nesting behaviour of Harpagifer bispinis in Arthur Harbour, Antarctic Peninsula. J Fish Biol 12:465–474
Daniels RA (1983) Demographic characteristics of an Antarctic plunderfish, Harpagifer bispinis antarcticus. Mar Ecol Progr Ser 13:181–187
Derome N, Chen W, Dettai A, Bonillo C, Lecointre G (2002) Phylogeny of Antarctic dragonfishes (Bathydraconidae, Notothenioidei, Teleostei) and related families based on their anatomy and two mitochondrial genes. Mol Phylogenet Evol 24:139–152
Diez MJ, Romero MC, Obenat S, Albano MJ, Tapella F (2009) Distribution of benthic invertebrates in the Beagle Channel, Argentina. An Inst Patagon 37:29–40
Duhamel G, Hulley P-A, Causse R, Koubbi P, Vacchi M, Pruvost P, Vigetta S, Irisson J-O, Mormède S, Belchier M, Dettai A, Detrich HW, Gutt J, Jones CD, Kock K-H, Lopez Abellan LJ, Van de Putte AP (2014) Chapter 7. Biogeographic patterns of fish. In: De Broyer C, Koubbi P, Griffiths HJ, Raymond B, Udekem d’Acoz C, et al. (eds) Biogeographic Atlas of the Southern Ocean. Scientific Committee on Antarctic Research, Cambridge, pp 328–362
Eastman JT (2005) The nature of the diversity of Antarctic fishes. Polar Biol 28:93–107
Eastman JT (2017) Bathymetric distributions of notothenioid fishes. Polar Biol 40:2077–2095
Eastman JT (2019) An analysis of maximum body size and designation of size categories for notothenioid fishes. Polar Biol 42:1131–1145
Eastman JT, Eakin RR (2016) Nototheniod species list. https://people.ohio.edu/eastman/research.htm. Accessed 25 Oct 2019
Eschmeyer WN, Fricke R, van der Laan R (eds) Catalog of fishes: genera, species, references.https://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp. Accessed 7 Oct 2019
Everson I (1968) Larval stages of certain Antarctic fishes. Br Antarct Surv Bull 16:65–70
Fernández DA, Ceballos SG, Malanga G, Boy CC, Vanella FA (2012) Buoyancy of sub-Antarctic notothenioids including the sister lineage of all other notothenioids (Bovichtidae). Polar Biol 35:99–106
Fernández DA, Bruno DO, Llompart FM (2018) Length–weight relationship of six notothenioid species from sub-Antarctic waters (Beagle Channel, Argentina). J Appl Ichthyol 35:597–599
Gordillo S, Rabassa J, Coronato A (2008) Paleoecology and paleobiogeographic patterns of mid-Holocene mollusks from the Beagle Channel (southern Tierra del Fuego, Argentina). Rev Geol Chile 35:321–333
Hüne M, Vega R (2016) Feeding habits in two sympatric species of Notothenioidei, Patagonotothen cornucola and Harpagifer bispinis, in the Chilean Patagonian channels and fjords. Polar Biol 39:2253–2262
Hüne M, González-Wevar C, Poulin E, Mansilla A, Fernández DA, Barrera-Oro E (2015) Low level of genetic divergence between Harpagifer fish species (Perciformes: Notothenioidei) suggests a Quaternary colonization of Patagonia from the Antarctic Peninsula. Polar Biol 38:607–617
Hureau JC (1990) Harpagiferidae. In: Gon O, Heemstra PC (eds) Fishes of the Southern Ocean. JLB Smith Institute of Ichthyology, Grahamstown, pp 357–363
Isla F, Bujalesky G, Coronato A (1999) Procesos estuarinos en el canal Beagle, Tierra del Fuego. Rev Asoc Geol Arg 54:307–318
Kellermann A (1990) Catalogue of early life stages of Antarctic notothenioid fish. Ber Polarforsch 67:45–136
Kimura DK (1980) Likelihood methods for the von Bertalanffy growth curve. Fish Bull 77:765–776
Kock K-H (1992) Antarctic fish and fisheries. Cambridge University Press, Cambridge, p 359
Kock KH, Kellermann A (1991) Reproduction in Antarctic notothenioid fish-a review. Antarct Sci 3:125–150
Koubbi P, Duhamel G, Camus P (1991) Early life stages of Notothenioidei from the Kerguelen Islands. Cybium 14:225–250
Krebs WN (1983) Ecology of neritic marine diatoms, Arthur Harbor, Antarctica. Micropaleontology 29:267–297
La Mesa M, Vacchi M (2001) Age and growth of high Antarctic notothenioid fish. Antarct Sci 13:227–235
La Mesa M, Vera-Duarte J, Landaeta MF (2017) Early life history traits of Harpagifer antarcticus (Harpagiferidae, Notothenioidei) from the South Shetland Islands during austral summer. Polar Biol 40:1699–1705
Lloris D, Rucabado J (1991) Ictiofauna del Canal Beagle (Tierra de Fuego), aspectos ecológicos y análisis biogeográfico. Publ Espec Inst Esp Oceanogr 8:1–182
Matallanas J (1997) Sobre algunos peces con interés biogeográfico de las Islas Orcadas del Sur. Bol Real Soc Esp Hist Nat (Sec Biol) 93:87–92
Navarro JM, Paschkeb K, Ortiza A, Vargas-Chacoff L, Pardoa LM, Valdivia N (2019) The Antarctic fish Harpagifer antarcticus under current temperatures and salinities and future scenarios of climate change. Progr Oceanogr 174:37–43
Near TJ, Cheng CHC (2008) Phylogenetics of notothenioid fishes (Teleostei: Acanthomorpha): Inferences from mitochondrial and nuclear gene sequences. Mol Phylogenet Evol 47:832–840
North AW (1991) Review of the early life history of Antarctic notothenioid fish. In: di Prisco G, Maresca B, Tota B (eds) Biology of Antarctic fish. Springer, Berlin, pp 70–86
Pérez A, Calvo J, Tresguerres M, Luquet C (2003) Aglomerularism in Harpagifer bispinis: a sub-Antarctic notothenioid fish living at reduced salinity. Polar Biol 23:800–805
Rabassa J (2008) The late Cenozoic of Patagonia and Tierra del Fuego. Elsevier, Amsterdam
Rae GA, Calvo J (1995) Fecundity and reproductive habits in Patagonothen tessellata (Richardson, 1845) from the Beagle Channel, Argentina. Antarct Sci 7:235–240
Rae GA, Calvo J (1996) Histological analysis of gonadal development in Patagonotothen tessellata (Richardson 1845) (Nototheniidae: Pisces) from the Beagle Channel, Argentina. J Appl Ichthyol 12:31–38
Riccialdelli L, Newsome SD, Fogel ML, Fernández DA (2017) Trophic interactions and food web structure of a sub-Antarctic marine food web in the Beagle Channel: Bahía Lapataia, Argentina. Polar Biol 40:807–821
Tomo AP (1981) Contribucion a1 conocimiento de la fauna ictiologica del sector Antartico Argentino. Contrib Inst Antart Argent 14:1–242
Tomo AP, Cordero Funes JR (1972) Edad y crecimiento en largo del Harpagifer bispinis antarcticus. Contrib Inst Antart Argent 96:1–19
Van der Molen S, Matallanas J (2004) Reproductive biology of female Antarctic spiny plunderfish Harpagifer spinosus (Notothenioidei: Harpagiferidae), from Îles Crozet. Antarct Sci 16:99–105
Vanella FA, Fernández DA, Romero MC, Calvo J (2007) Changes in the fish fauna associated with a sub-Antarctic Macrocystis pyrifera kelp forest in response to canopy removal. Polar Biol 30:449–457
White MG (1991) Age determination in Antarctic fish. In: di Prisco G, Maresca B, Tota B (eds) Biology of Antarctic fish. Springer, Berlin, pp 87–100
White MG, Burren PJ (1992) Reproduction and larval growth of Harpagifer antarcticus Nybelin (Pisces, Notothenioidei). Antarct Sci 4:421–430
Zama A, Cárdenas E (1984) Descriptive catalogue of marine and freshwater fishes from the Aysén Region, Southern Chile, with Zoogeographical Notes on the Fish Fauna. Servicio Nacional de Pesca and Japan International Cooperation Agency, Santiago
Zar JH (1999) Biostatistical analysis. Prentice Hall, Michigan
Acknowledgements
This work was supported by the Consejo Nacional de Investigaciones Científicas y Técnicas (Grant Numbers PIP 0440 and PICT 2321). The authors are also grateful to Christopher D. Jones for their useful comments that improved the manuscript, Raimundo Bustos for his help with the map and M. Isabel Ledesma for her assistance with the English language editing of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Ethical approval
Harpagifer bispinis is not protected under wildlife conservation laws (local legislation, International Union for Conservation of Nature [IUCN], or Convention on International Trade Endangered Species [CITES]). Individuals used under this study were captured within the framework of the Pampa Azul inter-ministerial initiative, supported by the Ministerio de Ciencia, Tecnología e Innovación Productiva of Argentina. As the Consejo Nacional de Investigaciones Cientificas y Técnicas (CONICET) does not possess formal Committees regarding fish welfare and sampling protocols, fish handling during sampling was performed following guidelines of the Ethical Committee of the Universities Federation for Animal Welfare (UFAW) Handbook on the Care and Management of Laboratory Animals (available online at https://www.ufaw.org.uk).
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Llompart, F.M., Fernández, D.A., Aureliano, D. et al. Life-history traits of the Magellan plunderfish Harpagifer bispinis (Forster, 1801) in the Beagle Channel (Tierra del Fuego, South America). Polar Biol 43, 1643–1654 (2020). https://doi.org/10.1007/s00300-020-02735-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-020-02735-x