
Abstract
Anthropogenic activities affect habitat use by Rangifer tarandus, a particularly vulnerable species due to grouping behavior, extensive movements, and grazing ecology. We studied habitat use of GPS-collared reindeer in relation to surface mining activities during the snow-free season in Finnmark, Norway over a period of 7 years. Based on information about the mine’s level of operation (amount of people, vehicles, and equipment in operation) and rock blasting schedule, we divided data into high-activity periods (workdays) and low-activity periods (mine closed for ca 2.5 days on weekends and a yearly 3-week holiday period). We further divided workdays into periods with and without rock blasting and associated high-noise days. We found that reindeer significantly reduced habitat use at closer distances to the mine, indicating an influence zone up to 1.5 km. Reductions in use were strongest closest to the mine in high-activity periods. No avoidance effect of the mine was found beyond approximately 0.9 km for the 3-week holidays, 1.0 km for weekends, and 1.5 km for workdays with or without rock blasting. Compared to holidays and weekends, probability of use was reduced by 30–34% within 1.3 km from the mine for workday blasting periods, and up to 35% within 1.4 km for other workdays. Since averted areas can be partly utilized again within days or weeks following intensive mining activity periods, reduced mining activity in crucial periods for reindeer, such as during calving and migration, can be an effective mitigation measure.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Surface mining for aggregates and metals generates revenue, jobs, and necessary raw materials for society, yet it is by nature a disruptive industry that often affects sites in otherwise pristine nature around the globe (Haley et al. 2011). Negative effects of surface mining have been shown for entire ecosystems (Palmer et al. 2010), including effects on behavior and habitat use for a number of wildlife species such as caribou Rangifer tarandus (e.g. Hebblewhite 2008; Polfus et al. 2011; Boulanger et al. 2012; Plante et al. 2018), grizzly bear Ursus arctos (Cristescu et al. 2016a, b), bighorn sheep Ovis canadensis (Sargeant and Oehler 2007), American pronghorn Antilocapra antilocapra, elk Cervus elaphus (Sawyer et al. 2007), and mule deer Odocoileus hemionus (Sawyer et al. 2006). Surface mining creates relatively high levels of disturbance stimuli, including vegetation stripping, drilling, blasting, processing, and driving bulldozers and dump trucks, so wildlife in surrounding areas perceive these activities and indirect habitat loss is inevitable. While some studies discuss specific negative effects of mining on caribou, such as disturbance from human activity (e.g. Polfus et al. 2011) and dustfall on pastures (Boulanger et al. 2012; Chen et al. 2017), knowledge about how surface mining affects reindeer in Fennoscandia is still limited.
As a keystone species of Arctic and sub-Arctic ecosystems, sustainable management of Rangifer populations has important ecological, cultural, and societal implications (Sandström et al. 2003). From a management perspective, precise knowledge on how human activities disturb wildlife is crucial in order to make sound decisions for new and existing mining projects. Geological resources are abundant in Fennoscandia, and a large increase in mining is expected based on increasing demands for aggregates for building infrastructure and mineral resources for existing and emerging industries (Bjerkgård et al. 2016). Surface mines in Fennoscandia are often located in remote areas, and mountainous areas overlapping with reindeer habitat. Reindeer herders in Norway, have land rights for reindeer grazing in a large part of the territory north of 62°N, and conflicts in land use have increased substantially in recent years (Pape and Löffler 2012; Johnsen 2016). There is a pressing need for improved knowledge to avoid or mitigate negative effects of present and future mining (Herrmann et al. 2014; Johnsen 2016), but this requires a good understanding of what those negative effects consist of.
Mountain- and tundra-dwelling R. tarandus (reindeer and caribou) graze in large groups over extensive ranges in relatively open landscapes, and due to this large space requirement are known to be more vulnerable towards human disturbances than most cervids (Stankowich and Reimers 2015). Possible effects of disturbances require large scale second- and third-order home range selection studies, since reindeer has been shown to avoid relatively large areas surrounding infrastructure and human activity (e.g. Anttonen et al. 2011; Boulanger et al. 2012; Panzacchi et al. 2013; Plante et al. 2018). However, large spatial scale effects reported for reindeer are often based on analyses of data taken over limited periods, thereby introducing the difficulty of accounting for confounding factors and of separating effects actually related to disturbances from spatiotemporal fluctuations in habitat use (Colman et al. 2017; Flydal et al. 2019). For instance, Dyer et al. (2001), Vistnes and Nellemann (2001), Skarin et al. (2004), Skarin (2007), Dahle et al. (2008), Polfus et al. (2011), and Helle et al. (2012) have only 1 or 2 years/seasons of data with all data merged for each year/season. Moreover, effect estimates are highly uncertain for studies on reindeer habitat use that rely solely on data from the period after the disturbance was introduced (see review by Bartzke et al. 2014).
To address these issues, we have studied area use by reindeer in connection with one of the world’s largest quartzite mines, located in Finnmark, Norway (NGU 2015). Our main objective was to estimate the effect of different amounts of human activity and noise in connection with surface mining on reindeer habitat use. Local herdsmen (Frode Utsi and Stig Rune Smuk, ‘Pers. comm.’) have informed us anecdotally that their reindeer’s grazing patterns changed following the establishment of the mine in 1973. While their reindeer had previously used the area intensively, the herdsmen reported that reindeer now avoid the mining area and show alarm reactions during loud human activities (e.g. bulldozing, drilling, rock blasting, and dumping rocks). Considering the operational setting of the mine, we chose to compare periods with high versus low levels of mine operation and with or without blasting and its associated activities (drilling, bulldozing, dumping rocks, etc.). Lacking a Before–After Control–Impact (BACI) design, with data from before establishment of the mine, this “off–on” study design allows for robust conclusions about how reindeer habitat use changes in response to different levels of mining activity. Furthermore, due to terrain barriers (steep mountains and fjord) and manmade barriers (fence bordering the seasonal pasture), the study area was limited to only include areas on the mine side of these barriers. Within this area, the animals could move freely up to approximately 10 km away from the mine. Since the study area is relatively small, we know for sure that a large proportion of it is directly affected by the human activity related to the mine, both through smell, sound and visual stimuli. Combined with the large number of temporal replications of high and low-activity periods (see Table 1), it is possible to make better conclusions about cause and effect compared to studies within larger areas, having fewer temporal replications (e.g. Polfus et al. 2011; Boulanger et al. 2012). We proposed the following predictions:
-
(1)
Given similar habitat and range properties, reindeer would decrease their habitat use closer to the mine.
-
(2)
The reduction in use closer to the mine would be stronger in periods with high mining activity (workdays with and without blasting) versus low-mining activity (holidays and weekends).
-
(3)
The reduction in use closer to the mine would be stronger for workdays with rock blasting and associated activities than workdays without blasting.
-
(4)
The reduction in use closer to the mine would be stronger for short periods of low-mining activity (i.e. weekends) than for the 3-week holidays. We assumed minimal differences in the mine’s activity levels during weekends compared to holidays, thus the predicted difference in avoidance for those periods would relate to length of the low-activity period (ca 2.5 days for weekends and 3 weeks for holidays).
Materials and methods
Study area
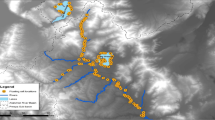
The mine, opened in 1973 by the Sydvaranger mining company (Pevik 2015), is located in Finnmark county, Northern Norway at 70°28′10″N, 28°31′3″E (Fig. 1). It is located within the spring, summer, and autumn pasture lands for reindeer belonging to the herding district Rakkonjarga. The winter herd size varied between 3717 and 4168 animals during the years of our study, lasting from 2011 to 2017. Semi-domesticated reindeer herds in Norway typically consist of almost 80% females (e.g. Holand et al. 2003), and the summer herd is between 50 and 75% larger than the winter herd, depending on calving rate and survival. Since we studied effects of the mine in summer and autumn, the herd may have been > 6500 reindeer, including calves. We defined our study area based on the surrounding landscape, including reindeer fences and information from herders about local habitat use (Fig. 1). Pastures surrounding the mine contain birch forest at altitudes of < 200 m.a.s.l., and low-alpine vegetation in altitudes of 200–350 m.a.s.l., with a few peaks slightly over 400 m.a.s.l. The reindeer moves into this area mostly from the north, and graze here predominantly during summer and autumn (Frode Utsi, ‘Pers. Comm.’).

Location map of the study area showing the mine (marked in dark brown color) and road network. The boundaries of the study area were defined by landscape barriers, such as steep slopes, ocean, and main road. (Color figure online)
The mine is situated along the coastline to the west, and the study area extends out to approx. 10 km east of the mine. Along the eastern border of the study area, the terrain rises steeply towards a mountain plateau of 300–600 m, with less preferred forage, and decreased availability of habitats due to natural barriers of rocky and steep terrain (Frode Utsi, ‘Pers.comm.’). A reindeer fence approx. 4–5 km south of the mine and the ocean on the western side restrict movement out of the area on those sides. Thus, herders are able to keep their animals gathered in this area both before leading the herd to another area for calf-marking and slaughtering in a pen approx. 8–9 km east of the study area in summer. The reindeer are here again after the rut, before migration to their late autumn and winter pastures in autumn begins (Frode Utsi, ‘Pers. Comm.’). The late autumn pastures are located 5–40 km south of the mine, while the winter pasture is located more than 40 km to the south. A few animals may also move through our study area from south to north during spring on their way to calving ranges in the north.
Methods
We studied habitat use by an average of 31 (range 23–34) GPS-collared adult female reindeer per year. GPS Plus collars from Vectronics Aerospace GmbH were programmed to register animal position every three hours. We investigated reindeer habitat use in the study area from between when they arrived in their summer range in April/May until they left in October/November during 2011–2017. Since reindeer grazing preferences and their habitat use naturally changes during the year (Reimers et al. 2014), we divided the year into three seasons: spring (May–June), summer (July–August), and autumn (September–October). However, we had too little data for the spring season (see Table 1) and excluded this season from further analysis. Descriptive statistics (i.e. used/available points) for each of the two seasons (summer and autumn) across distance zones are presented to show general distribution of use (see Online Resource: Fig. S1). A pre-analysis of reindeers’ habitat use in relation to distance to the mine and season (i.e. categorical variable with two levels: summer and autumn), using generalized linear mixed model (GLMM) with a use-availability design (Manley et al. 2002), showed a similar avoidance effect for summer and autumn (see Online Resource: Fig. S2). Thus, we combined summer and autumn data in the final analysis.
We defined our test periods based on information from the mining company about differing activity levels in the mine. First, we divided the dataset into two main categories, (1) high activity periods, i.e. “workdays” (0700 Monday–2100 Friday = 4 days and 14 h) when human activity was high and activities like running heavy machinery, vehicle traffic and drilling were more likely to happen, and (2) low-activity periods, i.e. “weekends” (2100 Friday–0600 Monday = 2 days and 10 h) and the yearly, “3-week summer holiday”, when human activity was low and the activities mentioned above were much less likely to happen. Then, based on daily data on rock blasting, we divided the high activity period into two new categories, i.e. workdays with blasting and without blasting. The mining company informed us that rock blasting was characterized by heavy drilling about 1 day before and transport of aggregate about one day after the actual blasting day, so we included one day before and after blasting in the “workdays with blasting” category. The activity periods used in the final analysis were thus defined as: holidays (i.e. 3 weeks in summer), weekends, workdays with blasting, and workdays without blasting. Occasional blasting activities occurred during weekends, but so infrequently (see Table 1) that these “weekends with blasting” were excluded from the analyses. This also allowed for better isolation of low-activity mining periods from high-activity mining periods. We used the low-activity periods as reference levels in our models. We assumed that even in the low-activity periods there are some human activity in the mine potentially affecting reindeer, and thus, there were never levels of “no” activity.
In addition to the main variable of interest in this study (i.e. distance to the mine), we derived elevation, slope, and aspect variables that are suspected to influence reindeer habitat selection in this area (Iversen et al. 2014). All variables were screened for collinearity using variance inflation factors (VIF; Zuur et al. 2009), with VIF ≥ 3.0 as a threshold for removing a variable. Because of high collinearity for vegetation (VIF = 6.39), and its correlation with elevation in high Arctic areas such as this study area, we did not include vegetation in the analysis (Mårell et al. 2006; Colman et al. 2013). We used a decay function to account for a probable decrease in the impact of infrastructure with increasing distance (Aue et al. 2012). We then followed (Buchanan et al. 2014) to develop the decay distance variable as a function of Euclidean distance to the mine with the use of the form e(−d/a), where “d” was the distance from each pixel to the mine in meters and “a” represents constant values of 100, 500, 1000, 1500, 2000, 3000, 4000, and 5000. The constant values shape the function (Carpenter et al. 2010), each value corresponds to approximate influence zones (i.e. < 0.5 up to < 10 km; see Online Resource: Fig. S3), as animals’ responses to landscape features probably decrease at greater distances (Nielsen et al. 2009; Skarin et al. 2018). The decay function scaled the distance variables between 1 and 0, with values increasing in proximity to the mine (Nielsen et al. 2009; Buchanan et al. 2014).
Data analysis
We predicted reindeer’s habitat use using RSF (resource selection function) models with a use-availability design (Manley et al. 2002) by fitting binomial family generalized linear mixed models (GLMM) in R with the lme4 package (Bates et al. 2014) and glmmsr package (Ogden 2015). The response variable was binomial (used/available), consisting of used reindeer GPS positions and an equal amount of randomly sampled available points within the defined study area, i.e. equivalent to a total of 19,020 used GPS-locations (see Table 1). The explanatory variables included decay distance from the mine (i.e. the decay indexes mentioned above), activity periods (i.e. “holidays”, “weekends”, “workdays without blasting”, and “workdays with blasting”), slope, elevation, and aspect. We included individual reindeer year as random factor to account for individual yearly variations (Zuur et al. 2009). Akaike’s information criterion corrected for small sample sizes (AICc)-values and delta AICc were used to identify the most parsimonious model (see Online Resource: Table S1). To illustrate the results from the RSF models, we calculated predicted probabilities of selection and the 95% confidence intervals (CI) to show the marginal effects of the distance to the mine for the different activity periods. All analyses were done in R version 3.5.1 (R Core Team 2018).
Results
The decay index 1/500 gave the best model fit compared to the other decay indexes (Online Resource: Table S1), indicating an approximate influence zone of < 1.5 km, i.e. the effect of distance to the mine becomes imperceptible after 1.5 km. The variables identified by the most parsimonious model include elevation, slope, aspect, and decay distance to mine interacting with mine activity levels (see Online Resource: Table S1).
We found that reindeer significantly reduced their use of habitats closer to the mine in high-activity periods (Fig. 2; Table 2). In comparison with the 3-week holiday, model predictions on relative probability of use showed an average reduction of 34% for workdays with blasting within 1.3 km, and 35% within 1.4 km for workdays without blasting. Compared to weekends, we found a reduction in relative probability of use within 1.3 km by 30% and 34% for workdays with and without blasting, respectively (Fig. 2; Table 2). We found no difference in habitat use for reindeer on workdays with vs. without blasting, suggesting that rock blasting events did not have additional negative effect. Probability of use did not differ between holidays and weekends (Fig. 2; Table 2).

Predicted probability of use (±95% CI) in relation to distance to the mine and mining activity periods from the top predictive model for reindeer resource selection (RSF) in the snow-free season (i.e. summer and autumn) during 2011–2017 in Finnmark, Northern Norway. The predicted probabilities for the distance predictor variable were calculated while keeping the other continuous predictor variables constant (at their mean values)
In general, there was still a baseline reduction in habitat use during both holidays and weekends. The reduced use in relation to the mine becomes minimal after 0.9 km for the 3-week holiday, 1.0 km for weekends, and 1.5 km during workdays (both with and without blasting) (Fig. 2). We also found effects of elevation, slope, and aspect (Table 2), showing reduced use at lower or higher elevations, on steeper slopes, and more use for the north facing slope.
Discussion
Our findings show a general negative effect from a surface mine on reindeer habitat use, confirming prediction 1. Furthermore, periods of high activity result in a stronger negative response, indicating that human activity represents a key disturbance stimuli for reindeer, confirming prediction 2. This becomes clear when using the period with the lowest level of operations (the 3-week holiday) as a reference level, and is also supported by Polfus et al. (2011), which only found minor effects from mines in seasons when human activity was low. Similarly, Eftestøl et al. (2016) only found negative effects during construction for power lines when human activity was high. However, there was no difference in effects between workdays with and without blasting or when comparing the 2.5 day weekends vs. the 3-week holiday. This means that neither prediction 3 or 4 was supported. Possibly, at the high level of general mining operations during workdays, blasting does not add additional stimuli leading to increased negative reactions by reindeer. Furthermore, it seems like a day or two with low activity is enough to allow animals to resume grazing closer to the mine.
Through separating (1) higher mining activity periods of minimum 3 days’ length (workdays with blasting) and up to ca 4.5 days’ length (workdays without blasting), and (2) low-activity periods of ca 2.5 days (weekends) and 3 weeks (holidays) over a period of 7 years, we were able to make up to 38 temporal replications for each season (see Table 1). The large number of replications in our study assures that effect estimates for each activity period are not affected differently by confounding factors. Thus, the 30–35% reduced use of habitats up to 1.4 km from the mine in high activity periods compared to periods with low-mining activity is likely caused by the change of human activity. This shows how functional habitats are affected as a consequence of mining disturbance, and are similar to findings for other large herbivores (e.g. White and Gregovich 2017).
A general increase in area use farther from the mine, up to approx. 0.9 (low activity) and 1.5 km (high activity), suggests that avoidance responses occur within the same distance interval. However, our analysis did not include data on habitat use from before the mine was established, thus we are uncertain about these estimates. Some effects of reindeer habitat preference, like seasonal pasture difference, and terrain steepness (Skarin et al. 2008), are partly accounted for by including elevation, slope, and aspect in our model predictions. However, other factors such as landscape features (e.g. mountains hilltops, rivers, lakes, etc.), reindeer seasonal migration pattern, predators, insect harassment, and other human activities most likely also affect reindeer use of this area independent of the mine (e.g. Skogland 1984; Nybakk et al. 1999; Frid and Dill 2002; Pape and Löffler 2012; Iversen et al. 2014). For instance, according to the herdsmen, the mine site was previously more preferred than the rest of the study area due to high-quality pasture, low human disturbance, and insect relief near the coast (Frode Utsi, ‘Pers. comm.’). This suggests that general effects of the mine, including both high and low-mining activity periods, are likely larger than shown in our analysis.
Our results showed a reduced habitat use up to, but not beyond 1.5 km, and supports that the study area, encompassing an area within 10 km, was large enough to test the effects from the mine in question, i.e. the effects would not be larger if we included areas on the far side of the barriers. However, in another context, e.g. if reindeer graze freely over a larger area with more uniform habitats and without barriers, our results may have been different. This applies both to the results in general and in relation to differences between work days with and without blasting even though the reductions in semi-domesticated reindeers’ use of habitats at distances up to 1.5 km from human disturbances are supported by previous findings in Fennoscandia (e.g. Anttonen et al. 2011; Skarin et al. 2015; Eftestøl et al. 2016).
The negative effects found in our study are in line with previous findings in mine disturbance studies for other cervids such as caribou, big horn sheep, and mountain goats (e.g. Jansen et al. 2009; White and Gregovich 2017). It is likely that visual, scent and noise stimuli from heavy machinery, blasting, vehicle traffic, and human activity, all combined disturb or frighten reindeer within the distance of perception (e.g. Stankowich and Reimers 2015). Noise in particular may have disturbing effects at long distances (Drolet et al. 2016), but animals may show behavioral habituation to a repeated noise-stimuli that are not associated with danger (Stankowich 2008; Bejder et al. 2009; Stankowich and Reimers 2015), indicating that ungulates do not consistently associate noise and human activity with an increase in predation risk (or cannot afford to maintain responsiveness to the most frequent human stimuli), but may allow for greater investment in fitness-enhancing activities (Brown et al. 2012). However, a range of mechanisms can lead to changes in tolerance (Bejder et al. 2009). For instance, Helle et al. (2012) reported adverse effects of outdoor activities on reindeer area use, however the tolerance level increased with improved channelling of tourists into fewer and better marked hiking and ski trails. We found the highest use close to the mine site during the 3-week holiday period, and more use of habitats near the mine site during weekends than workdays. It seems, therefore, that avoidance is mainly related to direct disturbance events connected to human activity (Reimers et al. 2007; Reimers and Eftestøl 2012), as well as a long term avoidance of habitats with increased disturbance risk (Bleicher 2017). To a lesser extent, this might also be the case for the surface mine in our study, where even after 3 weeks of low-mining activity, reindeer maintained less use of habitats within 0.9 km of the mine.
Based on the results of our study and other findings (e.g. Johnson et al. 2005; Weir et al. 2007; Jansen et al. 2009; Boulanger et al. 2012; Herrmann et al. 2014; Johnson and Russell 2014; Cristescu et al. 2016a; White and Gregovich 2017), it seems that zones of influence for Rangifer surrounding mine sites could vary from < 1.5 km and up to > 30 km. Some of the studies on effects of mines may lack substantiation (Hebblewhite 2008) or have weaknesses in relation to the methods used (Flydal et al. 2019), but the variation in conclusions may also be related to context dependency. For example, our study is on semi-domesticated reindeer, known to show weaker response to humans compared to wild reindeer (Baskin and Hjalten 2001).
Improved data on area use for mining projects in Rangifer habitat are needed to consider interactions with landscape, migration patterns and herding practice, as well as direct effects from the intensity of mining activity. Although we conclude that more negative effects result from periods of higher mining activity, we are less certain about the general effects of the mine independent of activity periods. Since we have found that reindeer resume some use of previously disturbed habitats within days, it is likely that negative effects of mining can be partly mitigated by reducing the intensity of mining activities in critical periods for reindeer. An effective mitigation measure might be to keep mining activities to a minimum during periods when reindeer uses the area intensively, such as during calving and migration. GPS-monitoring of animals could also be an important tool before development starts (Herrmann et al. 2014) to implement mitigation measures of this kind after initiation of a new mine or other human developments.
References
Anttonen M, Kumpula J, Colpaert A (2011) Range selection by semi-domesticated reindeer (Rangifer tarandus tarandus) in relation to infrastructure and human activity in the boreal forest environment, Northern Finland. Arctic 64:1–14
Aue B, Ekschmitt K, Hotes S, Wolters V (2012) Distance weighting avoids erroneous scale effects in species-habitat models. Methods Ecol Evol 3:102–111. https://doi.org/10.1111/j.2041-210X.2011.00130.x
Bartzke GS, May R, Bevanger K, Stokke S, Røskaft E (2014) The effects of power lines on ungulates and implications for power line routing and rights-of-way management. Int J Biodivers Conserv 6:647–662. https://doi.org/10.5897/IJBC2014.0716
Baskin LM, Hjalten J (2001) Fright and flight behavior of reindeer. Alces 37:435–445
Bates D, M. M, Bolker BM, Walker S (2014 ) Fitting linear mixed-effects models using lme4. https://arxiv.org/pdf/1406.5823.pdf. Accessed 04 Dec 2018
Bejder L, Samuels A, Whitehead H, Finn H, Allen S (2009) Impact assessment research: use and misuse of habituation, sensitisation and tolerance in describing wildlife responses to anthropogenic stimuli. Mar Ecol Prog Ser 395:177–185. https://doi.org/10.3354/meps07979
Bjerkgård T et al. (2016) Norway. In: Boyd R, Bjerkgård T, Nordahl B, Schiellerup H (eds) Mineral resources in the Arctic, 1st edn. NGU – Geological Survey of Norway, Tromsø
Bleicher SS (2017) The landscape of fear conceptual framework: definition and review of current applications and misuses. PeerJ 5:e3772
Boulanger J, Poole KG, Gunn A, Wierzchowski J (2012) Estimating the zone of influence of industrial developments on wildlife: a migratory caribou Rangifer tarandus groenlandicus and diamond mine case study. Wildl Biol 18:164–179. https://doi.org/10.2981/11-045
Brown CL, Hardy AR, Barber JR, Fristrup KM, Crooks KR, Angeloni LM (2012) The effect of human activities and their associated noise on ungulate behavior. PLoS ONE 7:e40505. https://doi.org/10.1371/journal.pone.0040505
Buchanan CB, Beck JL, Bills TE, Miller SN (2014) Seasonal resource selection and distributional response by Elk to development of a natural gas field. Rangel Ecol Manag 67:369–379. https://doi.org/10.2111/Rem-D-13-00136.1
Carpenter J, Aldridge C, Boyce MS (2010) Sage-grouse habitat selection during winter in Alberta. J Wildl Manag 74:1806–1814. https://doi.org/10.2193/2009-368
Chen W et al (2017) Does dust from Arctic mines affect caribou forage? J Environ Prot 8:258–276. https://doi.org/10.4236/jep.2017.83020
Colman JE, Eftestøl S, Tsegaye D, Flydal K, Mysterud A (2013) Summer distribution of semi-domesticated reindeer relative to a new wind-power plant. Eur J Wildl Res 59:359–370. https://doi.org/10.1007/s10344-012-0682-7
Colman JE, Bergmo T, Tsegaye D, Flydal K, Eftestøl S, Lilleeng MS, Moe SR (2017) Wildlife response to infrastructure: the problem with confounding factors. Polar Biol 40:477–482. https://doi.org/10.1007/s00300-016-1960-8
Cristescu B, Stenhouse GB, Boyce MS (2016a) Large omnivore movements in response to surface mining and mine reclamation. Sci Rep. https://doi.org/10.1038/srep19177
Cristescu B, Stenhouse GB, Symbaluk M, Nielsen SE, Boyce MS (2016b) Wildlife habitat selection on landscapes with industrial disturbance. Environ Conserv 43:327–336. https://doi.org/10.1017/S0376892916000217
Dahle B, Reimers E, Colman JE (2008) Reindeer (Rangifer tarandus) avoidance of a highway as revealed by lichen measurements. Eur J Wildl Res 54:27–35. https://doi.org/10.1007/s10344-007-0103-5
Drolet A, Dussault C, Cote SD (2016) Simulated drilling noise affects the space use of a large terrestrial mammal. Wildl Biol 22:284–293. https://doi.org/10.2981/wlb.00225
Dyer SJ, O'Neill JP, Wasel SM, Boutin S (2001) Avoidance of industrial development by woodland caribou. J Wildl Manag 65:531–542. https://doi.org/10.2307/3803106
Eftestøl S, Tsegaye D, Flydal K, Colman JE (2016) From high voltage (300 kV) to higher voltage (420 kV) power lines: reindeer avoid construction activities. Polar Biol 39:689–699. https://doi.org/10.1007/s00300-015-1825-6
Flydal K, Tsegaye D, Eftestøl S, Reimers E, Colman JE (2019) Rangifer within areas of human influence: understanding effects in relation to spatiotemporal scales. Polar Biol 42:1–16
Frid A, Dill LM (2002) Human-caused disturbance stimuli as a form of predation risk. Conserv Ecol 6:11
Haley S, Klick M, Szymoniak N, Crow A (2011) Observing trends and assessing data for Arctic mining. Polar Geogr 34:37–61. https://doi.org/10.1080/1088937X.2011.58444
Hebblewhite M (2008) A literature review of the effects of energy development on ungulates: implications for central and eastern Montana. Report prepared for Montana Fish, Wildlife and Parks, Miles City, MT
Helle T, Hallikainen V, Sarkela M, Haapalehto M, Niva A, Puoskari J (2012) Effects of a holiday resort on the distribution of semi-domesticated reindeer. Ann Zool Fenn 49:23–35
Herrmann TM et al (2014) Effects of mining on reindeer/caribou populations and indigenous livelihoods: community-based monitoring by Sami reindeer herders in Sweden and First Nations in Canada. Polar J 4:28–51. https://doi.org/10.1080/2154896X.2014.913917
Holand Ø, Røed KH, Mysterud A, Kumpula J, Nieminen M, Smith ME (2003) The effect of sex ratio and male age structure on reindeer calving. J Wildl Manag 67:25–33. https://doi.org/10.2307/3803058
Iversen M, Fauchald P, Langeland K, Ims RA, Yoccoz NG, Bråthen KA (2014) Phenology and cover of plant growth forms predict herbivore habitat selection in a high latitude ecosystem. PLoS ONE 9:e100780. https://doi.org/10.1371/journal.pone.0100780
Jansen BD, Krausman PR, Bristow KD, Heffelfinger JR, deVos JC (2009) Surface mining and ecology of desert bighorn sheep. Southwest Nat 54:430–438. https://doi.org/10.1894/Tal-01.1
Johnsen KI (2016) Land-use conflicts between reindeer husbandry and mineral extraction in Finnmark, Norway: contested rationalities and the politics of belonging. Polar Geogr 39:58–79. https://doi.org/10.1080/1088937x.2016.1156181
Johnson CJ, Boyce MS, Case RL, Cluff HD, Gau RJ, Gunn A, Mulders R (2005) Cumulative effects of human developments on arctic wildlife. Wildl Monogr 160:1–36
Johnson CJ, Russell DE (2014) Long-term distribution responses of a migratory caribou herd to human disturbance. Biol Conserv 177:52–63. https://doi.org/10.1016/j.biocon.2014.06.007
Manley BF, McDonald LL, Thomass DL, McDonald TL (2002) Resource selection by animals: statistical design and analysis for field studies. Kluwer Academic Publishers, Boston
Mårell A, Hofgaard A, Danell K (2006) Nutrient dynamics of reindeer forage species along snowmelt gradients at different ecological scales. Basic Appl Ecol 7:13–30. https://doi.org/10.1016/j.baae.2005.04.005
NGU (2015) Quartz resources in Norway: a varied spectrum. NGU Focus 11. NGU – Geological Survey of Norway, Tromsø
Nielsen SE, Cranston J, Stenhouse GB (2009) Identification of priority areas for grizzly bear conservation and recovery in Alberta, Canada. J Conserv Plan 5:38–60
Nybakk K, Kjelvik O, Kvam T (1999) Golden eagle predation on semidomestic reindeer. Wildl Soc B 27:1038–1042
Ogden H (2015) Fitting GLMMs with glmmsr. https://cran.r-project.org/web/packages/glmmsr/vignettes/glmmsr-vignette.pdf. Accessed 10 November 2018.
Palmer MA et al (2010) Mountaintop mining consequences. Science 327:148–149. https://doi.org/10.1126/science.1180543
Panzacchi M, Van Moorter B, Jordhøy P, Strand O (2013) Learning from the past to predict the future: using archaeological findings and GPS data to quantify reindeer sensitivity to anthropogenic disturbance in Norway. Landsc Ecol 28:847–859. https://doi.org/10.1007/s10980-012-9793-5
Pape R, Löffler J (2012) Climate change, land use conflicts, predation and ecological degradation as challenges for reindeer husbandry in Northern Europe: what do we really know after half a century of research? Ambio 41:421–434. https://doi.org/10.1007/s13280-012-0257-6
Pevik JA (2015) Geological and mineralogical characterization of the upper part of the Gamasfjell Quartzite at the Vaggecearu Mountain. Thesis, Norwegian University of Science and Technology (NTNU), M.Sc
Plante S, Dussault C, Richard JH, Cote SD (2018) Human disturbance effects and cumulative habitat loss in endangered migratory caribou. Biol Conserv 224:129–143. https://doi.org/10.1016/j.biocon.2018.05.022
Polfus JL, Hebblewhite M, Heinemeyer K (2011) Identifying indirect habitat loss and avoidance of human infrastructure by northern mountain woodland caribou. Biol Conserv 144:2637–2646. https://doi.org/10.1016/j.biocon.2011.07.023
R Core Team (2018) R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. https://www.R-project.org/
Reimers E, Dahle B, Eftestøl S, Colman JE, Gaare E (2007) Effects of a power line on migration and range use of wild reindeer. Biol Conserv 134:484–494. https://doi.org/10.1016/j.biocon.2006.08.034
Reimers E, Eftestøl S (2012) Response behaviors of Svalbard reindeer towards humans and humans disguised as polar bears on Edgeoya. Arct Antarct Alp Res 44:483–489. https://doi.org/10.1657/1938-4246-44.4.483
Reimers E, Tsegaye D, Colman JE, Eftestøl S (2014) Activity patterns in reindeer with domestic vs. wild ancestry. Appl Anim Behav Sci 150:74–84. https://doi.org/10.1016/j.applanim.2013.10.010
Sandström P et al (2003) Conflict resolution by participatory management: remote sensing and GIS as tools for communicating land-use needs for reindeer herding in northern Sweden. Ambio 32:557–567. https://doi.org/10.1639/0044-7447(2003)032[0557:Crbpmr]2.0.Co;2
Sargeant GA, Oehler MW (2007) Dynamics of newly established elk populations. J Wildl Manag 71:1141–1148. https://doi.org/10.2193/2006-247
Sawyer H, Nielson RM, Lindzey F, McDonald LL (2006) Winter habitat selection of mule deer before and during development of a natural gas field. J Wildl Manag 70:396–403. https://doi.org/10.2193/0022-541x(2006)70[396:Whsomd]2.0.Co;2
Sawyer H, Nielson RM, Lindzey FG, Keith L, Powell JH, Abraham AA (2007) Habitat selection of Rocky Mountain elk in a nonforested environment. J Wildl Manag 71:868–874. https://doi.org/10.2193/2006-131
Skarin A (2007) Habitat use by semi-domesticated reindeer, estimated with pellet-group counts. Rangifer 27:121–132
Skarin A, Danell Ö, Bergstrom R, Moen J (2008) Summer habitat preferences of GPS-collared reindeer Rangifer tarandus tarandus. Wildl Biol 14:1–15. https://doi.org/10.2981/0909-6396(2008)14[1:Shpogr]2.0.Co;2
Skarin A, Danell Ö, Bergström R, Moen J (2004) Insect avoidance may override human disturbances in reindeer habitat selection. Rangifer 24:95–103
Skarin A, Nellemann C, Ronnegard L, Sandstrom P, Lundqvist H (2015) Wind farm construction impacts reindeer migration and movement corridors. Landsc Ecol 30:1527–1540. https://doi.org/10.1007/s10980-015-0210-8
Skarin A, Sandström P, Alam M (2018) Out of sight of wind turbines—reindeer response to wind farms in operation. Ecol Evol 00:1–14. https://doi.org/10.1002/ece3.4476
Skogland T (1984) Wild reindeer foraging-niche organization Holarct Ecol 7:345–379
Stankowich T (2008) Ungulate flight responses to human disturbance: a review and meta-analysis. Biol Conserv 141:2159–2173. https://doi.org/10.1016/j.biocon.2008.06.026
Stankowich T, Reimers E (2015) Escape decisions in mammals. In: Cooper JE, Blumstein DT (eds) Escaping from predators: an integrative view of escape decisions. Cambridge University Press, London.
Vistnes I, Nellemann C (2001) Avoidance of cabins, roads, and power lines by reindeer during calving. J Wildl Manag 65:915–925. https://doi.org/10.2307/3803040
Weir JN, Mahoney SP, McLaren B, Ferguson SH (2007) Effects of mine development on woodland caribou Rangifer tarandus distribution. Wildl Biol 13:66–74. https://doi.org/10.2981/0909-6396(2007)13[66:Eomdow]2.0.Co;2
White KS, Gregovich DP (2017) Mountain goat resource selection in relation to mining-related disturbance. Wildl Biol. https://doi.org/10.2981/wlb.00277
Zuur AF, Ieno EN, Walker NJ, Saveliev AA, Smith GM (2009) Mixed effects models and extensions in ecology with R. Springer, New York
Acknowledgements
This work was supported by the Norwegian Research Council (NRC project 255635). We would like to thank reindeer herders from the Rakkonjarga district for very meaningful cooperation, lending us their reindeer, providing important background information, and assisting with GPS tagging. We also thank Lex Folk for language editing; Roland Pape, Troy Hegel, and two other anonymous reviewers for their suggestions and improvements on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
The research reported here has been conducted in an ethical and responsible manner and complies with all relevant legislation. All authors agree with the contents of the manuscript and its submission to the journal.
Research involving human and animal participants
All procedures performed in this study were in accordance with the ethical standards of the institutions or practice at which the study was conducted. This article does not contain any studies with human participants performed.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Eftestøl, S., Flydal, K., Tsegaye, D. et al. Mining activity disturbs habitat use of reindeer in Finnmark, Northern Norway. Polar Biol 42, 1849–1858 (2019). https://doi.org/10.1007/s00300-019-02563-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-019-02563-8