
Abstract
The South Sandwich Islands, in the South Atlantic Ocean, are a major biological hot spot for penguins and other seabirds, but their remoteness and challenging coastlines preclude regular biological censuses. Here we report on an extensive survey of the South Sandwich Islands, the first since the late 1990s, which was completed through a combination of direct counting, GPS mapping, and interpretation of high-resolution commercial satellite imagery. We find that the South Sandwich Islands host nearly half of the world’s Chinstrap Penguin (Pygoscelis antarctica) population (1.3 million breeding pairs), as well as c. 95,000 breeding pairs of Macaroni Penguins (Eudyptes chrysolophus), and several thousand breeding pairs of Gentoo Penguins (Pygoscelis papua). Despite being at the northern edge of their breeding range, we found an unexpectedly large (≥125,000 breeding pairs) population of Adélie Penguins (Pygoscelis adeliae). Additionally, we report that nearly 1900 pairs of Southern Giant Petrels (Macronectes giganteus) breed in the South Sandwich Islands, 4 % of the global population, almost all of which are found on Candlemas Island. We find that the South Sandwich Islands have not experienced the same changes in penguin abundance and distribution as the rest of the Scotia Arc and associated portions of the western Antarctic Peninsula. This discovery adds important context to the larger conversation regarding changes to penguin populations in the Southern Ocean.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Changes in the distribution and abundance of penguin populations on the Antarctic Peninsula and associated islands of the Scotia Arc have been widely reported. On the western Antarctic Peninsula, Adélie Penguins (Pygoscelis adeliae) and Chinstrap Penguins (Pygoscelis antarctica) are declining north of Marguerite Bay whereas Gentoo Penguins (Pygoscelis papua) are increasing in abundance and expanding their breeding range to the south (Trivelpiece et al. 2011; Lynch et al. 2012b; Casanovas et al. 2015). Macaroni Penguins (Eudyptes chrysolophus) are declining sharply at South Georgia (Trathan et al. 2012), while King Penguin (Aptenodytes patagonicus) populations have increased (Trathan et al. 2007). While these trends appear quite robust, the drivers responsible for these changes are far less clear; hypotheses include the direct influence of sea ice on habitat suitability for ice-intolerant species, the indirect effects of sea ice on krill availability, krill harvesting, tourism and human disturbance, and cetacean recovery (Ducklow et al. 2007; Murphy et al. 2007; Trathan et al. 2007; Ainley et al. 2010; Lynch et al. 2010; Trivelpiece et al. 2011).
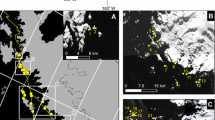
The South Sandwich Islands (Fig. 1), a long isolated chain of volcanic islands along the Scotia Arc connecting South Georgia with the Antarctic Peninsula, have long been identified as a major ecological hot spot for penguins, but their remote location and challenging coastline have made it difficult to gain accurate estimates of seabird breeding abundance and distribution. Several factors make the South Sandwich Islands unique and potentially important for disentangling the drivers of changes seen elsewhere along the Scotia Arc. The extent of mid-winter sea ice (as defined by the boundary of 15 % sea ice concentration) cuts right through the South Sandwich Islands in most years; as a result, the northernmost islands of Zavodovski and Visokoi are often ice-free all year whereas the southernmost islands are rarely, if ever, ice-free all year round (Murphy et al. 2007). The waters surrounding the South Sandwich Islands have some of the highest Antarctic krill (Euphausia superba) and chlorophyll-a densities in the Southern Ocean (Atkinson et al. 2008), yet some of the lowest rates of krill harvest (CCAMLR 2015). While tourism activity has increased tenfold over the last 20 years on the Antarctic Peninsula (Bender et al. 2016), commercial cruise trips to the South Sandwich Islands are rare. For all of these reasons, the South Sandwich Islands represent a key element of our regional understanding of how environmental or human influences affect seabird abundance and distribution.

South Sandwich Islands relative to Antarctica (inset)
Prior surveys of the South Sandwich Islands include Baker et al. (1964), Cordier et al. (1981), and Convey et al. (1999), though of these, only Convey et al. (1999) provides estimates of breeding abundance against which to assess changes in the abundance and distribution of seabirds and seals. The recent development of high-resolution commercial satellite imagery as a tool for surveying penguin populations (e.g., Lynch et al. 2012a) provides a promising alternative to direct survey for areas as remote as the South Sandwich Islands. Here we report on a coordinated ground and satellite imagery survey of the South Sandwich Islands which updates Convey et al. (1999), provides some estimates of population change among seabirds and lays the groundwork for future surveys through improved distribution maps, geo-referenced photographs, and suggested guidelines for future ground surveys.
Methods
Ground survey
We (RW and AB) surveyed the South Sandwich Islands from January 1 to February 5, 2011, using the MS Golden Fleece, skippered by Jerôme Poncet, as a platform for survey and access to the islands. We worked from north to south through the archipelago making Zodiac landings to conduct biological surveys on all the main islands in the group, with the exception of Leskov, which (by all available accounts) presents an extraordinarily difficult landing (Table 1). Our methods for surveying breeding seabirds varied depending on the species involved, the stage of the breeding cycle, the size of the colony, and the time available onshore. In general, we counted smaller colonies (<10,000 individuals) in full by direct counting of occupied nests and/or chicks using handheld tally-counters. In larger colonies or areas with limited time ashore, we mapped colony boundaries with GPS to facilitate the interpretation of high-resolution satellite imagery (e.g., Lynch et al. 2012a). Tables 1 and 2 provide further details on the field survey. Our survey was biased toward identifying the locations of penguin colonies, and no attempt was made to record the exact locations of cliff nesting petrels (Antarctic Fulmar, Cape Petrel, Snow Petrel). It is important to note that we do not yet understand the breeding phenology of seabirds on the South Sandwich Islands well enough to deduce the date of peak nesting and thus correct abundance estimates for nest failures; however, we do make note of the approximate phase of breeding for each site where it provides context for our reported counts.
Satellite imagery survey
Table 3 contains a summary of satellite imagery used for this analysis. We used the same basic technique for manual interpretation of satellite imagery described in Lynch et al. (2012a) and LaRue et al. (2014). We manually delineated penguin subcolonies using the guano stain associated with the nesting area of the colony. Our identification of guano was based on prior experience with ground validated imagery interpretation at other locations [e.g., Chinstrap Penguins on Deception Island, Naveen et al. (2012); Adélie Penguins on the western Antarctic Peninsula, Casanovas et al. (2015)], GPS data obtained during the ground survey, and photodocumentation. Guano stains are distinguished from background substrates (usually volcanic sand, mineral soils, or bare rock) by their spectral reflectance (their ‘color’ as seen in false color images), texture, size, and distance to the shoreline. In many cases, interpretation of satellite imagery was aided by GPS data collected on the ground (e.g., tracklogs encircling subcolonies) and photodocumentation, the latter of which was particularly important for distinguishing different species where they were nesting in adjacent or even fully mixed colonies. For Chinstrap Penguins, we were able to use field counts of subcolonies paired with satellite imagery of those same subcolonies to estimate the ‘apparent’ density of nesting penguins at Zavodovski Island; this number was used for all of the South Sandwich Islands. Note that this is not necessarily the true density as would be measured in the field, but the number of occupied nests in the guano-stained area identified in the satellite imagery as belonging to that subcolony [see LaRue et al. (2014)]. For the other species, we used nesting density estimates from the literature. It is important to note that while it is not possible to differentiate nesting penguins and non-nesting penguins from satellite imagery, the guano stain is created by penguins incubating nests over an extended period of time and reflects long-term occupation of an area rather than transient occupation by non-nesting penguins that may wander through the colony.
Survey uncertainties
Irrespective of the source for our estimates of abundance (direct ground count or satellite-based), we have adopted the ordinal scheme for denoting census precision used by previous authors (e.g., Croxall and Kirkwood 1979; Woehler 1993): accuracy = 1: accurate to better than ±5 %; accuracy = 2: accurate to 5–10 %; accuracy = 3: accurate to 10–15 % (in practice, we extend this upper limit to 25 % to ensure continuity); accuracy = 4: accurate to 25–50 %; accuracy = 5: accurate to nearest order of magnitude. Accuracy was assessed based on the observer’s prior experience counting seabirds and the methodology used. Counts with high precision (N1 or C1) represent direct counts of occupied nests and/or chicks by an observer standing just outside the colony (~5 m from the edge). In other cases, binoculars would have been used to estimate the sizes of populations, and in these cases, the precision of the counts would be downgraded accordingly. Where counts of individual groups of penguins within the site were counted with different accuracies, the site-wide accuracy is calculated by propagating errors accumulated in each colony count assuming independence among the colony counts.
While nest and chick counts have been reported separately, archipelago-wide total abundance estimates for each penguin species assume each penguin chick is associated with one nest when combining nest and chick counts. This conversion assumes ~50 % egg or chick loss prior to the survey, which may or may not be appropriate for each island. Therefore, archipelago-wide abundance estimates are less precise than each of the island-wide abundance estimates.
Results
Detailed island-specific accounts for each species, along with maps and photographs for each island, have been included as an Online Resource for use by other researchers (Online Resource 1). High-resolution satellite imagery, merged where necessary with existing coastline information, was used to create a GIS shapefile for the coastlines of the South Sandwich Islands (Online Resource 2).
Penguins
While large uncertainties preclude a quantitative assessment of population trends, our findings strongly suggest that Adélie, Chinstrap, and Macaroni Penguin populations are at least stable and are likely to have increased in abundance since the survey of Convey et al. (1999) (Table 4).
Chinstrap Penguins are the most abundant bird species in the South Sandwich Islands, and the massive scale of the Chinstrap colonies made direct census challenging at most locations. Nevertheless, we estimate a minimum population of 1.3 million breeding pairs (Table 4). Most pairs were still brooding chicks during the survey, so we used active nests as our unit of counting for Chinstrap Penguins, though an increased number of birds moving toward the crèche stage was observed at Thule Island at the end of the survey period. We estimate a minimum breeding population of 125,000 pairs of Adélie Penguins in the South Sandwich Islands (Table 4). Adélie Penguins were absent from the northern islands, with the northernmost record coming from Candlemas Island. Moving further south, Adélies became a more prominent part of the seabird community. Breeding was well advanced in all colonies with chicks in the crèche stage, and some birds already fledged; for this reason, we counted chicks rather than adults. Gentoo Penguins were present in low numbers within the surveyed area, typically in loose colonies at very low densities just above the beach. We estimate a minimum population of 1902 breeding pairs of Gentoo Penguins for the South Sandwich Islands. Most Gentoo Penguin pairs had large chicks that had reached the crèche stage at the time of census, so chick counts were used to derive population estimates. Where appropriate, an additional estimate was made of active nests with birds incubating eggs or small chicks. King Penguins were present in small numbers at all islands, typically <50 individuals, with most onshore to molt. The exception to this was at Zavodovski Island where, within the large Chinstrap Penguin colony on the southeast coast, two birds were seen in a posture that strongly suggested they were incubating eggs or brooding small chicks.
Macaroni Penguins were present at most locations, and we estimate a minimum abundance of 95,000 breeding pairs in the South Sandwich Islands (Table 4). Because Macaroni Penguins were usually present in small numbers within much larger Chinstrap Penguin colonies, locating birds in the field was not straightforward, and it was difficult to guarantee that all birds within a colony had been counted. However, it quickly became apparent that the Macaroni Penguin colonies could be located by their darker appearance (Figure S11a). This was thought to be a combination of the stance of the birds (more hunched for Macaroni Penguins brooding small chicks, more upright in Chinstrap Penguins brooding larger chicks) and the density of birds (higher in Macaroni than in Chinstrap). We do not know whether Macaroni and Chinstrap Penguins are always differentiable in this manner or whether we captured them at a fortuitous time in the breeding cycle. Nevertheless, as described in Lynch et al. (2012a), these differences were also apparent in high-resolution satellite imagery, allowing us to differentiate between Macaroni Penguins and Chinstrap Penguins (Figure S11b) and estimate populations from satellite imagery where field counts were highly uncertain or unavailable. Many birds were present on territories without apparently breeding. Those that were breeding were mainly brooding a small chick, and so active nests were counted.
Flying birds
Both Southern and Northern Giant Petrels (Macronectes giganteus and Macronectes halli, respectively) were a prominent feature of the penguin colonies visited, where they were scavenging/predating penguins both onshore and in coastal waters. Southern Giant Petrels were present throughout the islands and were the only species found breeding in the South Sandwich Islands. We found nearly 1900 pairs of Southern Giant Petrels breeding in the South Sandwich Islands, almost all of which were found on Candlemas Island (Table 4). White morph birds were noted at several sites. Northern Giant Petrels were recorded inshore and in coastal waters of all the islands, and this species was frequently observed scavenging/predating penguins. The largest number recorded of this species was 25 birds at Irving Point on Visokoi Island.
South Georgia Shags (Phalacrocorax [atriceps] georgianus) were present throughout the islands in small numbers, with rarely more than ten birds at any location. No effort was made to visit specific locations where this species has been recorded breeding in the past so updated population estimates are not available. Wilson’s Storm-petrels (Oceanites oceanicus) were frequently seen flying over land at probable breeding locations during the day, but no nests were located. Black-bellied Storm-petrels (Fregata tropica) were confirmed breeding on Candlemas and Montagu Islands. On the former, several dead chicks were found; on the latter, a dead chick and an occupied nest were found. Antarctic Terns (Sterna paradisaea) were present at Zavodovski Island where breeding was confirmed when chicks were seen on several offshore stacks at the northern end of the island. Adults were also seen carrying food flying northwest past Fume Point. An adult was also seen carrying food at Beach Point, Thule Island, in what seemed to be courtship display.
Kelp Gulls (Larus dominicanus) were present in small numbers (<100 fully grown birds) at all sites surveyed, but we recorded little evidence of breeding. Confirmed breeding was recorded for Zavodovski Island (three nest sites with chicks located on the north coast), Candlemas Island (on at least five sites on Breakbones Plateau, no chick count), Thule Island (minimum eight chicks at Hewison Point), and Bellingshausen Island (minimum 11 chicks).
Antarctic Skuas (Stercorarius antarctica) were present at all islands associating with penguin colonies and often seen in skua clubs, including up to 50 birds at Zavodovski Island. However, breeding activity was not widespread. The unusually large concentration of birds at Candlemas Island reported by Convey et al. (1999) was again noted, but a count of nests was abandoned due to time limitations. The next largest breeding population appeared to be on Bellingshausen Island where at least 20 nests were found. Otherwise, breeding activity was either limited to single nests or absent altogether. Individual South Polar Skuas (Stercorarius maccormickii) were seen on shore at several islands, including at Allen Point on Montagu Island and at Hewison Point on Thule Island. While there was no evidence of breeding, a South Polar Skua paired with an Antarctic Skua was found at Allen Point on Montagu Island.
Antarctic Fulmars (Fulmarus glacialoides) were observed breeding on steep slopes and cliff ledges on all islands except Zavodovski and Bellingshausen Islands. Although no direct counts of nests were attempted, breeding sites were photodocumented when encountered. In terms of distribution and relative abundance, the observations made during this survey are consistent with findings by Convey et al. (1999). Breeding Cape Petrels (Daption capense) were sighted on all the islands visited, despite generally in low numbers. The largest breeding aggregation of this species was observed on the west coast of Thule Island, where several thousand nests were observed and photographed. Snow Petrel (Pagodroma nivea) nests were seen on all the islands visited except Zavodovski Island. Like the Cape Petrel, the largest breeding aggregation of Snow Petrels was observed on the west coast of Thule Island. No attempt was made to census the island-wide population of Snow Petrels. We observed two Light-mantled Sooty Albatrosses (Phoebetria palpebrata) flying over Candlemas Island on January 17, 2011. These birds were behaving as a pair flying in tandem high over the island. They were not seen to settle at any point and departed after 15 min of observation after an encounter with some Antarctic Skuas. Finally, it is important to note that despite deliberate effort to search for Snowy Sheathbills (Chionis alba), no individuals were recorded at any of the South Sandwich Islands.
Discussion
Summary of observations and apparent trends
Our observations regarding the species composition of the South Sandwich Islands accord with those of previous surveys (Cordier et al. 1981; Convey et al. 1999). No new species were recorded breeding in the islands; however, we did identify six new locality records. The following species were confirmed breeding on islands where they had not previously been recorded: Gentoo Penguins were confirmed breeding on Zavodovski and Bristol Islands; Adélie Penguins were confirmed breeding on Cook Island; Macaroni Penguins were confirmed breeding on Vindication and Thule Islands; and Black-bellied Storm-petrels were confirmed breeding on Montagu Island.
While the census of breeding penguins was by no means complete, several observations can be made about penguin populations in the region. Gentoo Penguin numbers are stable or increasing overall. Locally, a decline may have been observed at Irving Point, Visokoi Island, although it is possible that part of the colony was not located. A marked increase in the Gentoo Penguin population was noted at Saunders Island where 300 pairs recorded in 1997 is now a colony producing over 800 chicks. At other sites, numbers are comparable with Convey et al. (1999), including Hewison Point which, having increased from 20 pairs in 1955 (Wilkinson 1956) to 600 pairs in 1997 (Convey et al. 1999), now appears to have stabilized.
On the evidence of those populations counted, the abundance of Macaroni Penguins in the islands is at least stable, or perhaps increasing. While the largest population in the archipelago (on Zavodovski Island) was not directly counted, we estimate that the colony includes more than the 52,000 pairs reported by Convey et al. (1999). The total population on Visokoi Island is probably between 4000 and 5000 pairs, higher than the 2000 pairs reported by Convey et al. (1999). The number of nests on Candlemas Island was similar to the total in 1997. A new site with about 200 nests was recorded on Vindication Island, where the species had been recorded previously (Holdgate and Baker 1979) but not proven to breed.
The Adélie Penguin population on Candlemas Island has changed little in size since 1997. However, Adélie Penguin colonies on Montagu, Bellingshausen, and Thule Islands were far larger than had been previously reported by Convey et al. (1999). For example, the chick count on Bellingshausen Island was about two-thirds complete with a total of 30,000 chicks; an estimate of the remainder of the colony suggests a total number of chicks in the range of 45,000 (C3). On Thule Island, where Convey et al. (1999) report only 10,000 Adélie pairs, we encountered 26,000–27,000 (C4) chicks even without a survey at Beach Point. No count was possible at Scarlett Point on Montagu Island, but the population clearly numbers in the thousands compared with 200 reported for the whole of Montagu Island by Convey et al. (1999). Initial impressions on Saunders Island were of a colony in excess of 50,000 pairs of Adélie Penguins at Cordelia Bay-Nattriss Point, but this is unconfirmed. As a result, the Adélie Penguin population estimate for the South Sandwich Islands could be doubled to at least 125,000 pairs. Finally, we note that the absence of Adélie Penguins at the two most northerly islands (Zavodovski and Visokoi), which lie north of the 15 % mid-winter sea ice contour, is consistent with other studies suggesting 15 % as a lower limit of acceptability for Adélie Penguins (Ainley 2002).
Chinstrap Penguin colonies encountered were generally too large to permit direct counts. However, in all areas where direct counts or reasonable estimates could be made, colonies were at least as large, if not larger, than those reported by Convey et al. (1999). In some cases, this appeared to be a result of survey methodology, particularly in cases where we landed at colony sites rather than conduct boat surveys that involved obscured views of the colony.
For Adélie, Chinstrap, and Macaroni Penguins, it seems clear that the massive declines reported on South Georgia, in the South Orkney Islands, and along the western Antarctic Peninsula (e.g., Trivelpiece et al. 2011; Lynch et al. 2012b; Trathan et al. 2012) are not reflected in the trends seen in the South Sandwich Islands. The South Sandwich Islands have received considerably less attention than either South Georgia or the western Antarctic Peninsula, and little is known about changes in prey availability, foraging effort, diet, phenology, or demography. Casual examination of the guano suggests that penguins are eating both krill and fish in the South Sandwich Islands, though more careful analysis is clearly required to compare penguin diet in this region to populations elsewhere in the region. Advection in the vicinity of the South Sandwich Islands is complex, and it is not clear what fraction of krill in the South Sandwich Islands are coming from the area of the South Orkney Islands or further to the east from a clockwise gyre in the Weddell Sea (Murphy et al. 2004; Thorpe et al. 2007). As such, it is difficult to know whether the diverging trends between the South Sandwich Islands and the South Orkneys/western Antarctic Peninsula region reflects differences in krill production and subsequent availability or differences in fishing pressure. A more detailed understanding of krill transport to this region will be needed to explain why penguin populations in the South Sandwich Islands have escaped the significant declines seen elsewhere in the region. It was notable that many penguin colonies were located on ‘recent’ volcanic ash overlying ice that, while temporarily stabilized by penguin guano, was thin enough that we would occasionally break through the ‘crust’ to the underlying ice. In some areas, penguin breeding habitat was being eroded as the underlying ice melted. While volcanic activity can create habitat for penguins to colonize, warming of the region may lead to melting and erosion of available nesting habitat.
Compared to Convey et al. (1999), we observed a notable increase in the numbers of Northern Giant Petrels. Only two individuals were recorded during the earlier survey, while we regularly encountered groups of at least 10 individuals onshore with a peak count of 25 birds at Irving Point on Visokoi Island. However, we cannot confirm whether this species is breeding. We found evidence consistent with continued population growth for Southern Giant Petrel populations in the South Sandwich Islands [at Candlemas Island: 520 pairs in 1979 (Cordier et al. 1981); 1516 pairs in 1997 (Convey et al. 1999); 1818 in 2011 (present survey)], though recognize the caveats appropriate to comparing surveys using different methodologies (Creuwels et al. 2005). In contrast, Shags appear to be declining throughout much of the archipelago. While we did not specifically target surveys for previously known locations of this species, Bristol Island was the only location where we found Shags nesting in numbers comparable to those given in Convey et al. (1999).
Several questions arise from the comparison of our survey with previous surveys (Cordier et al. 1981; Convey et al. 1999). For example, how are we to interpret the apparently large increase in the Adélie Penguin population in the southern islands? Cordier et al. (1981) report approximately 1000 pairs of Adélie Penguins at Herd Point. Convey et al. (1999) indicate Adélie Penguins as present at Herd Point [Figure 12 in Convey et al. (1999)], but list the Thule Island population as approximately the same as the total for Hewison Point (10,000 pairs; cf. Convey et al. (1999) Table 3 with text on page 121); from this we infer that at the very least, the Herd Point population was far smaller than the 10,000 pairs found at Hewison Point. So, we are faced with evidence suggesting that in 13 years the Adélie Penguin population at Herd Point has gone from c. 1000 pairs to producing c. 20,000 chicks, while the Hewison Point population has remained stable over a 30-year period (Cordier et al. 1981). Furthermore, Convey et al. (1999) report ‘<10,000’ pairs of Adélie Penguins at Bellingshausen Island, while we estimate the current population to have produced 40,000–50,000 chicks. Another Adélie Penguin site with a recorded landing in Convey et al. (1999) is Scarlett Point on Montagu Island where a 1.5-h landing was made on January 29, 1997. This would appear to be the source of the total of 200 pairs given for Montagu Island in Convey et al. (1999) Table 3, while evidence of thousands of Adélie Penguins were observed at this site during this survey (though we were unable to make a landing for a more precise count). It is difficult to know to what extent these differences stem from survey effort as opposed to genuine population growth, but the consistency between Cordier et al. (1981) and Convey et al. (1999), as well as the fact that three different islands appear to have growing Adélie populations, suggest the latter.
Missing elements of the South Sandwich Islands community
Cordier et al. (1981) noted four species believed to be missing from the South Sandwich Islands breeding bird community: Antarctic Terns, Antarctic Prions, Black-bellied Storm-petrels, and Sheathbills. We confirm Convey et al.’s findings that Antarctic Terns and Black-bellied Storm-petrels nest on the South Sandwich Islands, leaving Antarctic Prions and Sheathbills as the two remaining species that might be considered missing from the South Sandwich Islands’ avian community. Despite what appears to be an abundance of food and suitable habitat, no Sheathbills have ever been reported on the South Sandwich Islands. Cordier et al. (1981) note that the South Sandwich Islands lie in between the ranges of the Snowy Sheathbill (Chionis albus), which breeds to the west (closest known breeding colony is South Georgia, 540 km away), and the Black-faced Sheathbill (C. minor), which breeds to the east (closest known colony is Prince Edward Islands, 4345 km away), and suggest that insufficient prospecting of the islands rather than environmental conditions explains their absence. However, Sheathbills are believed to migrate from the Antarctic Peninsula to Argentina (BirdLife International 2015a), placing the South Sandwich Islands well within their physical flight range (see also Cordier et al. 1981). The apparently unfilled niche for a terrestrial scavenger specialist like the Sheathbill at the South Sandwich Islands remains a curious mystery.
South Sandwich Islands in a global context
While the South Sandwich Islands contain <2 % of the global population of Gentoo Penguins, Adélie Penguins, and Macaroni Penguins, it contains more significant populations of several other species. Our results suggest that the South Sandwich Islands host a minimum population of 1.3 million breeding pairs of Chinstrap Penguins, which is nearly half of the world’s Chinstrap Penguin population (assuming a total population of 2.67 million breeding pairs; Harris et al. 2015), and approximately 4 % of the global population of Southern Giant Petrels [BirdLife International 2015b; see also Patterson et al. (2008)]. In addition, roughly 13–31 % of the global population of Antarctic Fulmars breed in the South Sandwich Islands (Creuwels et al. 2007).
Future survey work
To facilitate future survey efforts, we have included in the Online Resources additional survey notes and advice for future expeditions. We have also provided ArcGIS shapefiles representing the location of penguin colonies, flying birds, and marine mammals. The photographs in Online Resource 1 are only a small subset of images available from the ground survey, and future survey teams are invited to contact us for additional imagery as needed.
While satellite imagery may permit some regular monitoring of highly visible species, it is impractical to conduct regular direct censuses of all islands within the archipelago. Therefore, it will be necessary to select a representative sample of accessible sites that could be monitored regularly as access to the region permits. We suggest regular monitoring of Southern Giant Petrels and Adélie Penguins on Candlemas Island and Chinstrap, Adélie, and Gentoo Penguins at Hewison Point on Thule Island. We suggest, as a second tier of priority surveys, Adélie Penguins on Montagu Island; Adélie Penguins and Gentoo Penguins at Cordelia Bay, Saunders Island; and Southern Giant Petrels and Chinstrap Penguins on Zavodovski Island. We hope that future visits to these remote and inhospitable islands will permit more rigorous analyses of biological trends in this area, and more comparative work with the Antarctic Peninsula to its south.
References
Ainley DG (2002) The Adélie Penguin: bellweather of climate change. Columbia University Press, New York
Ainley D, Ballard G, Blight LK, Ackley S, Emslie SD, Lescroël A, Olmastroni S, Townsend SE, Tynan CT, Wilson P, Woehler E (2010) Impacts of cetaceans on the structure of Southern Ocean food webs. Mar Mammal Sci 26:482–498
Atkinson A, Siegel V, Pakhomov EA, Rothery P, Loeb V, Ross RM, Quetin LB, Schmidt K, Fretwell P, Murphy EJ, Tarling GA, Fleming AH (2008) Oceanic circumpolar habitats of Antarctic krill. Mar Ecol Prog Ser 362:1–23
Baker PE, Holdgate MW, Longton RE, Tilbrook PJ, Tomblin JF, Vaughan RW, Wynne-Edwards CJC (1964) A survey of the South Sandwich Islands. Nature 203:691–693
Bender NA, Crosbie K, Lynch HJ (2016) Patterns of tourism in the Antarctic Peninsula Region: a 20 year re-analysis. Antarct Sci (in press)
BirdLife International (2015a) Species factsheet: Chionus albus. http://www.birdlife.org. Accessed 17 Jan 2015
BirdLife International (2015b) Species factsheet: Macronectes giganteus. http://www.birdlife.org. Accessed 25 Jun 2015
Casanovas PV, Naveen R, Forrest S, Poncet J, Lynch HJ (2015) A comprehensive coastal seabird survey maps out the front lines of ecological change on the western Antarctic Peninsula. Polar Biol 38:927–940
Commission for the Conservation of Antarctic Living Marine Resources (CCAMLR) (2015) Statistical Bulletin 25
Convey P, Morton A, Poncet J (1999) Survey of marine birds and mammals of the South Sandwich Islands. Polar Rec 35:107–194
Cordier J-R, Mendez A, Mougin JL, Visbeek G (1981) Les oiseaux des l’ile Thulé archipel des Sandwich du Sud (58°28′S, 27°20′W). L’oiseau et RFO 51:147–160
Creuwels JCS, Stark JS, Woehler EJ, van Franeker JA, Ribic CA (2005) Monitoring of a Southern Giant Petrel Macronectes giganteus population on the Frazier Islands, Wilkes Land, Antarctica. Polar Biol 28:483–493
Creuwels JCS, Poncet S, Hodum PJ, van Franeker JA (2007) Distribution and abundance of the Southern Fulmar Fulmarus glacialoides. Polar Biol 30:1083–1097
Croxall JP, Kirkwood ED (1979) The distribution of penguins on the Antarctic Peninsula and islands of the Scotia Sea. British Antarctic Survey, Cambridge
Ducklow HW, Baker K, Martinson DG, Quetin LB, Ross RM, Smith RC, Stammerjohn SE, Vernet M, Fraser W (2007) Marine Pelagic ecosystems: the West Antarctic Peninsula. Philos T R Soc B 362:67–94
Harris CM, Lorenz K, Fishpool LDC, Lascelles B, Cooper J, Coria NR, Croxall JP, Emmerson LM, Fraser WR, Fijn RC, Jouventin P, LaRue MA, Le Maho Y, Lynch HJ, Naveen R, Patterson-Fraser DL, Peter H-U, Poncet S, Phillips RA, Southwell CJ, van Franeker JA, Weimerskirch H, Wienecke B, Woehler EJ (2015) Important Bird Areas in Antarctica 2015 Summary. BirdLife International and Environmental Research & Assessment Ltd., Cambridge
Holdgate MW, Baker PE (1979) The South Sandwich Islands: I. General description. Br Antarct Surv Sci Rep 91:1–76
LaRue MA, Lynch HJ, Lyver POB, Barton K, Ainley DG, Pollard A, Fraser WR, Ballard G (2014) A method for estimating colony sizes of Adélie penguins using remote sensing imagery. Polar Biol 37:507–517
Lynch HJ, Crosbie K, Fagan WF, Naveen R (2010) Spatial patterns of tour ship traffic in the Antarctic Peninsula region. Antarct Sci 22:123–130
Lynch HJ, Naveen R, Trathan PN, Fagan WF (2012a) Spatially integrated assessment reveals widespread changes in penguin populations on the Antarctic Peninsula. Ecology 93:1367–1377
Lynch HJ, White R, Black AD, Naveen R (2012b) Detection, differentiation, and abundance estimation of penguin species by high-resolution satellite imagery. Polar Biol 35:963–968
Murphy EJ, Thorpe SE, Watkins JL, Hewitt R (2004) Modeling the krill transport pathways in the Scotia Sea: spatial and environmental connections generating the seasonal distribution of krill. Deep Sea Res Part II 51:1435–1456
Murphy EJ, Watkins JL, Trathan PN, Reid K, Meredith MP, Thorpe SE, Johnston NM, Clarke A, Tarling GA, Collins MA, Forcada J, Shreeve RS, Atkinson A, Korb R, Whitehouse MJ, Ward P, Rodhouse PG, Enderlein P, Hirst AG, Martin AR, Hill SL, Staniland IJ, Pond DW, Briggs DR, Cunningham NJ, Fleming AH (2007) Spatial and temporal operation of the Scotia Sea ecosystem: a review of large-scale links in a krill centred food web. Philos T R Soc B 362:113–148
Naveen R, Lynch HJ (2011) Antarctic Peninsula compendium, 3rd edn. Environmental Protection Agency, Washington
Naveen R, Lynch HJ, Forrest S, Mueller T, Polito M (2012) First direct, site-wide penguin survey at Deception Island, Antarctic, suggests significant declines in breeding chinstrap penguins. Polar Biol 35:1879–1888
Patterson DL, Woehler EJ, Croxall JP, Cooper J, Poncet S, Peter H-U, Hunter S, Fraser WR (2008) Breeding distribution and population status of the Northern Giant Petrel Macronectes Halli and the Southern Giant Petrel M. Giganteus. Mar Ornithol 36:115–124
Thorpe SE, Murphy EJ, Watkins JL (2007) Circumpolar connections between Antarctic krill (Euphausia superba Dana) populations: investigating the roles of ocean and sea ice transport. Deep Sea Res Part I 54:792–810
Trathan PN, Forcada J, Murphy EJ (2007) Environmental forcing and Southern Ocean marine predator populations: effects of climate change and variability. Philos T R Soc B 362:2351–2365
Trathan PN, Ratcliffe N, Masden EA (2012) Ecological drivers of change at South Georgia: the krill surplus, or climate variability. Ecography 35:983–993
Trivelpiece WZ, Hinke JT, Miller AK, Reiss CS, Trivelpiece SG, Watters GM (2011) Variability in krill biomass links harvesting and climate warming to penguin population changes in Antarctica. Proc Natl Acad Sci USA 108:7625–7628
Wilkinson JV (1956) South Sandwich Islands-birdlife. Sea Swallow 9:18–20
Woehler EJ (1993) The distribution and abundance of Antarctic and subantarctic penguins. Scientific Committee on Antarctic Research, Cambridge
Acknowledgments
Thanks to the crew of the MS Golden Fleece for getting us there, getting us ashore and back again, and keeping us fed and watered along the way. It is hard to imagine that it would have been possible to get so much done without the experience of Jérôme Poncet, who surely knows these islands better than anyone else. Thanks to Tom Hart who participated in the survey and provided notes and photographs, as well as to Thomas Hopper for help digitizing the coastline for GIS support. Many thanks are due to Dr. Keith Reid, Dr. Peter Convey, Dr. Jeroen Creuwels, and one anonymous reviewer, all of whom contributed significantly during the review process. HL and RN would like to acknowledge the Tinker Foundation and the Jeniam Foundation for financial support contributing to the expedition, and HL would like to acknowledge the US National Science Foundation Office of Polar Programs and Geography and Spatial Sciences (Award Nos. 07-39515 and 12-55058) for financial support of the data analysis.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Lynch, H.J., White, R., Naveen, R. et al. In stark contrast to widespread declines along the Scotia Arc, a survey of the South Sandwich Islands finds a robust seabird community. Polar Biol 39, 1615–1625 (2016). https://doi.org/10.1007/s00300-015-1886-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-015-1886-6