
Abstract
Arctic cisco Coregonus autumnalis have a complex anadromous life history, many aspects of which remain poorly understood. Some life history traits of Arctic cisco from the Colville River, Alaska, and Mackenzie River basin, Canada, were investigated using molecular genetics, harvest data, and otolith microchemistry. The Mackenzie hypothesis, which suggests that Arctic cisco found in Alaskan waters originate from the Mackenzie River system, was tested using 11 microsatellite loci and a single mitochondrial DNA gene. No genetic differentiation was found among sample collections from the Colville River and the Mackenzie River system using molecular markers (P > 0.19 in all comparisons). Model-based clustering methods also supported genetic admixture between sample collections from the Colville River and Mackenzie River basin. A reanalysis of recruitment patterns to Alaska, which included data from recent warm periods and suspected changes in atmospheric circulation patterns, still finds that recruitment is correlated to wind conditions. Otolith microchemistry (Sr/Ca ratios) confirmed repeated, annual movements of Arctic cisco between low-salinity habitats in winter and marine waters in summer.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Arctic cisco (Coregonus autumnalis) is an important cultural and subsistence resource in Alaska and Canada (Reist and Bond 1988; Gallaway et al. 1989). For example, in Alaska, an annual subsistence fishery occurs in the Colville River as fish gather to overwinter in brackish waters of the Colville River Delta (Fig. 1). In addition to the Colville River, Arctic cisco overwinter in the Sagavanirktok River (Fig. 1) but are not believed to spawn in any Alaska rivers (Craig 1984; Moulton 1989; Morales et al. 1993). Rather, Arctic cisco in Alaskan waters are thought to originate from spawning aggregations found over 650 km to the east in the Mackenzie River, Canada (Fig. 1), the only documented spawning population in North America (Moulton 1989; Fechhelm et al. 2007). This proposed scenario, in which Arctic cisco caught in Alaskan rivers are the progeny of Arctic cisco spawning in Canada, has been called the “Mackenzie hypothesis” (Gallaway et al. 1983; Bickham et al. 1989; Morales et al. 1993).

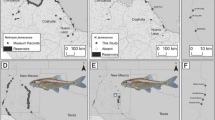
Genetic sample collection locations (black circles) for Arctic cisco from the Colville River and Mackenzie Rivers, 2005–2007
According to the Mackenzie hypothesis, it is assumed that age-0 fish emerge from spawning habitats in the Mackenzie River and are flushed or move downstream into ice-free waters of the Mackenzie River delta (Craig 1989). It is assumed that some age-0 fish that are abundant in delta channels during their first summer (Reist and Bond 1988; Fechhelm et al. 2007) are transported westward in the Beaufort Sea by wind-driven currents. Recruitment of Arctic cisco is controlled by wind speed and strength (Fechhelm and Griffiths 1990), and strong easterly winds result in higher abundance of Arctic cisco in coastal Alaskan waters and rivers, whereas lower recruitment occurs when westerly winds prevail (Fechhelm and Fissel 1988; Fechhelm and Griffiths 1990; Schmidt et al. 1991; Colonell and Gallaway 1997). Once in Alaskan waters, Arctic cisco are assumed to make annual migrations between summer feeding habitats in the Beaufort Sea and overwintering habitats in river deltas (Craig 1989). Between the ages of 6 and 8 years, Arctic cisco are thought to reach sexual maturity and return to the Mackenzie River system to spawn.
While predominately based on observed relationships between recruitment patterns into Alaskan rivers and wind speed and direction, the Mackenzie hypothesis has also been examined using genetic tools. Previous investigations using DNA restriction site analysis and allozymes did not find evidence for genetic differentiation between collections of Arctic cisco from the Colville River and Mackenzie River basin, a finding that supports the Mackenzie hypothesis (Bickham et al. 1989; Morales et al. 1993). There is limited evidence, however, for genetic differentiation among sample collections of Arctic cisco from Mackenzie River tributaries (Bickham et al. 1989; Lockwood and Bickham 1991) and between collections from the Mackenzie River and Beaufort Sea near Point Barrow (Morales et al. 1993), findings that do not support the Mackenzie hypothesis. In addition, studies of age structure support the Mackenzie hypothesis. Arctic cisco are iteroparous (Reist and Bond 1988) and reported to live up to 19 years (Bond and Erickson 1997), but fish older than age-8 are rare in Alaskan waters (Murphy et al. 2007). Presumably, after the first spawning event, Arctic cisco remain within the Mackenzie River and delta and make annual migrations to feed at sea but do not return to rivers in Alaska. If there were local spawning populations in Alaska, it is assumed that fish older than age-8 would be more common.
While the Mackenzie hypothesis remains the most plausible and accepted explanation of recruitment of Arctic cisco to Alaska, we conducted this study to reexamine the Mackenzie hypothesis for three reasons. First, in spite of all the evidence to the contrary, both local residents and some scientists continue to ask whether some recruitment of Arctic cisco could originate from local-Alaska-spawning populations or from Asia (MBC Applied Environmental Sciences 2004). Second, methodological advances in genetic analyses warranted a reassessment using new techniques. Third, and most important, climatic and oceanographic changes during the last decade have resulted in diminished sea ice, warming, and changes in northern wind and atmospheric circulation patterns (Overland et al. 2008; Overland and Wang 2010). Given the critical role that temperature and wind play in the early life history and recruitment of Arctic cisco in Alaska (von Biela et al. 2011, 2013), a reassessment of the Mackenzie hypothesis, including recruitment and wind data that expands previous studies and includes data from the recent warm period (Overland et al. 2008), was warranted to better understand how changes in climate may affect this important subsistence resource.
In this study, we revisit the Mackenzie hypothesis using several lines of investigation. First, we use newer genetic analyses including microsatellite markers and mitochondrial DNA (mtDNA) sequence data to test for differentiation among collections of Arctic cisco from the Colville River and five tributaries of the Mackenzie River. Second, we reexamine the link between wind conditions in the Beaufort Sea, recruitment of Arctic cisco to the Colville River region, and subsequent harvest in the Colville River area using new data. Third, we examine the relation between recruitment during their first winter and age structure of Arctic cisco to confirm that a lack of recruitment of age-0 fish results in missing age-classes in subsequent fall migrations as proposed by Fechhelm and Griffiths (1990). Fourth, we compare the age composition of Arctic cisco in the Colville River fishery to those collected from the Peel River, a tributary of the lower Mackenzie River to determine whether there are differences consistent with the Mackenzie hypothesis. Finally, we examine patterns of strontium in otoliths of Arctic cisco to test the migration chronology of Arctic cisco that was proposed by Craig (1989). Strontium is substituted for otolith calcium in levels reflecting the concentration of strontium in the environment, which is generally greater in seawater than freshwater. Strontium-to-calcium ratios (Sr/Ca), therefore, can be used to trace the movement of fish among environments of different salinity (Campana 1999; Zimmerman et al. 2003; Brown et al. 2007). Combining these lines of inquiry, we provide a new test of the Mackenzie hypothesis in light of new methods of analysis and longer time series to improve our understanding of how this important resource may respond to future climate and development scenarios.
Materials and methods
Genetic sample collections
Arctic cisco were collected from the subsistence fishery on the Colville River near the village of Nuiqsut in 2005, 2006, and 2007 (n = 86, 176, and 136, respectively) (Fig. 1). Mature fish were also collected in late summer from five tributaries of the lower Mackenzie River where Arctic cisco are presumed to spawn: the Arctic Red (n = 60), and Peel (n = 54) rivers in 2007 and the Great Bear (n = 55), Carcajou (n = 52), and Mountain (n = 59) rivers in 2008 (Fig. 1). Arctic cisco were caught using monofilament gill nets at all collection locations.
DNA extraction and microsatellite amplification
Genomic DNA was extracted from fin, gill, or muscle tissue using the DNeasy Tissue Kit (Qiagen, Valencia, CA, USA) or Puregene DNA Extraction Kit (Gentra Systems Inc., Minneapolis, MN, USA) following the manufacturer’s protocols. Sixteen microsatellite loci were identified as nuclear genetic markers to assess population structure of Arctic cisco including the following: seven microsatellite loci originally isolated from congenerics (Cisco-90, Cisco-157, Cisco-181, Cocl-Lav4, Cocl-Lav6, Cocl-Lav8, and LGL BWF2; Patton et al. 1997; Turgeon et al. 1999; Rogers et al. 2004), three previously published microsatellite loci developed from Arctic cisco (Aut135, Aut139, and Aut151; Ramey et al. 2008), two unpublished loci developed from Arctic cisco for this study (Aut134 and Aut189, Table 1), and four unpublished microsatellite loci taken from GenBank (Bom3c, Bom17c, Bom21c, and Bom22b). Five microsatellite loci (Aut135, Bom21c, Cisco-90, Cocl-Lav6, and Cocl-Lav8) were subsequently excluded from our panel after preliminary analyses because of low allelic polymorphism (<4 alleles per locus).
Microsatellite loci were amplified using PCR in 10-μL volumes containing 4–50 ng of genomic DNA, 10 mM Tris–HCl (pH 8.3), 1.5 mM MgCl2, 50 mM KCl, 0.01 % gelatin, 0.01 % NP-40, 0.01 % Triton X-100, 3.7–4.0 μM each unlabeled primers, 0.1–0.3 μM labeled tail primer, 2 mM each dNTP, and 0.3–0.5 U of Taq DNA polymerase. Reactions were carried out in MJ Research (BIORAD, Hercules, CA, USA) or MWG thermocyclers (MWG Biotech Inc., Huntsville, AL, USA) with an initial denaturation time of 2 min at 94 °C followed by variable thermoprofiles (Table 1) and a final 10-min elongation step at 72 °C. PCR products were separated on 6 % polyacrylamide gels using a LI-COR Model IR2 automated fluorescent DNA sequencer (Lincoln, NE, USA). Forward primers were either directly fluorescently labeled to visualize alleles (Bom3c, Bom17c, Bom22b, LGL BWF2, Cisco-151, Cisco-181, and Cocl-Lav4) or were synthesized with universal tails on the 5′ ends and visualized by adding fluorescently labeled complementary-tailed sequences to the PCR reaction (Aut134, Aut139, Aut151, and Aut189). Size standards were created by determining allele sizes for specific samples at each locus relative to the M13 phage single nucleotide ladder, and these samples were then used in subsequent gels. To verify allele sizes across all loci for quality control, a minimum of 10 % of all samples were independently amplified and scored. GeneImagIR v4.05 (Lincoln, NE, USA) software was used to assign allele scores.
Mitochondrial DNA sequencing
A 594 base pair (bp) fragment of the mitochondrial ATP synthase VI subunit gene (ATPase 6) was sequenced for a subsample of 372 Arctic cisco (Colville River, n = 258; lower Mackenzie River system, n = 114) using previously published primers (H9208 and L8558; Giuffra et al. 1994). The ATPase 6 gene was selected for use in the current study as it has been used in previous genetic investigations of North American ciscoes (Turgeon and Bernatchez 2001, 2003). Primers were modified on the 5′ ends with a universal M13F(-29) or M13R tail (Oetting et al. 1995). Mitochondrial DNA (mtDNA) was amplified using a MJ Research thermocycler using PCR conditions as follows: initial denaturation at 94 °C for 1 min followed by 35 cycles of 94 °C for 1 min, 62 °C for 1 min, and 72 °C for 1 min ending with a final elongation step at 72 °C for 10 min. Sequencing reactions were conducted using a simultaneous bi-directional sequencing kit (Epicentere technologies, Madison, Wisconsin) and fluorescently labeled primers. The sequencing reaction was conducted using a RoboCycler® thermocycler (Stratagene, La Jolla, CA, USA) with the following conditions: initial denaturation at 92 °C for 2 min followed by 30 cycles of 92 °C for 30 s, 60 °C for 30 s, and 70 °C for 1 min. Sequences were visualized on a LI-COR Long ReadIR 4200 automated sequence using a 5.5 % acrylamide gel. Sequence data were aligned and proofed using AlignIR software (LI-COR, Lincoln, NE, USA). Nucleotide positions for sequences amplified from Arctic cisco were confirmed by alignment with the published sequence for the complete mtDNA genome of European whitefish (C. lavaretus) (Miya and Nishida 2000; GenBank Accession No. AB034824).
Population genetic statistics
Microsatellite allelic size ranges, total number of alleles (A), and observed and expected heterozygosity (H o and H e, respectively) were generated by locus and by population using FSTAT v2.9.3 (Goudet 2001). The average number of alleles was calculated using GENALEX (Peakall and Smouse 2006). Average allelic richness by population, allelic richness by locus, and private allelic richness across 11 nuclear loci were calculated using HP-RARE 1.0 (Kalinowski 2005), which uses the rarefaction procedure to account for variable sample sizes. An exact test for Hardy–Weinberg equilibrium (HWE) was done by locus using Arlequin v3.01 (Excoffier et al. 2005) with a Bonferroni correction (Sokal and Rohlf 1995; k = 11 loci, α = 0.05/11 or 0.0045). Arlequin v3.01 was used to test for pairwise linkage disequilibrium between all possible pairs of loci with a Bonferroni correction (k = 55 comparisons, α = 0.05/55 or 0.0009). Samples were divided into sample collections by location and year (n = 8) and major river systems (Colville and Mackenzie rivers) to examine population structure. Pairwise F ST comparisons between all sample collection pairs and major river systems were calculated using Arlequin v3.01, and a Bonferroni correction was applied (α = 0.0017 k = 28; α = 0.0167 k = 3; α = 0.025 k = 2, respectively). An analysis of molecular variance (AMOVA) using Arlequin v3.01 was conducted to quantify genetic variability and estimate its distribution among sampling collections (between years for the Colville samples and tributaries for the Mackenzie) and major river systems (Colville and Mackenzie).
The frequency of mtDNA haplotypes, haplotype diversity, and nucleotide diversity were calculated using Arlequin v3.01, and NETWORK version 4.5.1.6 (Fluxus Technology Ltd. 2009; http://www.flexus-engineering.com/sharepub.htm) was used to generate a median-joining network (Bandelt et al. 1999) of mtDNA haplotypes. Arlequin 3.01 was used to calculate pairwise F ST comparisons of haplotype frequencies between Arctic cisco from the Colville River and the Mackenzie River system.
Model-based clustering analysis
Allele frequencies for microsatellite loci were used to probabilistically assign genotypes to populations or clusters using STRUCTURE v2.3.1 (Pritchard et al. 2000). Two sets of analyses were conducted: one using genotypic data for all samples collected from the Colville River and Mackenzie River system (n = 678) and a second using only data derived from samples collected from five tributaries of the lower Mackenzie River system where Arctic cisco are presumed to spawn (n = 280). The number of populations or clusters (K) was assessed for values of K from 1 to 10 for the analysis of genotypic data for all sample collections combined (e.g., the possibility of up to ten “source populations” was explored). Values of K from 1 to 5 were assessed for genotypic data derived from samples collected from the lower Mackenzie River system (e.g., structuring among five tributaries was explored). Five replicate model runs were conducted for each assumed value of K using the admixture model with correlated allele frequencies for 100,000 Markov chain Monte Carlo repetitions after a burn-in of 10,000. The mean natural log probability of sample genotypes clustering to K populations for five replicates was used to calculate the posterior probability for each value of K. The highest penalized posterior probability of K, where mean natural log likelihoods for each value of K were penalized by one half of their variance, was inferred to be the most likely number of clusters or populations among sample genotypes (Hubisz et al. 2009).
Retrospective analyses of catch data
Recruitment of Arctic cisco to the Colville River region, catch rates of young-of-year fish during summer surveys in the Prudhoe Bay region, historical harvest data for the commercial and subsistence fall fishery, and environmental data compiled by Murphy et al. (2007) were examined. First, a recruitment index was constructed by standardizing log-transformed catch-per-unit-effort (CPUE) in Prudhoe Bay during summer (days of year 200–258) for age-0, age-1, and age-2 Arctic cisco and then averaging the age-specific indices corresponding to a given cohort. Thus, an improved recruitment estimate was obtained for each cohort or year-class based on catch rates at three ages sampled in three consecutive summers. The main environmental indicator to link recruitment to wind conditions in the Beaufort Sea was an annual index of average easterly wind speed for the period from July 1 to August 31 at the Deadhorse airport located at Prudhoe Bay (Murphy et al. 2007). The recruitment index was modeled as a function of easterly winds using a linear model with a threshold. Residuals from the model were used as an index of recruitment anomalies to examine the effect of climate conditions (i.e., near-shore temperature, ice conditions; Murphy et al. 2007) on anomalous recruitment using correlations and linear regression analyses.
Second, we examined harvest rates as indices of recruitment to the fishery. We examined estimates of age-specific catch rates (ages 5–8) for both the commercial and subsistence fishery. To test whether catch rates of 5-, 6-, 7-, and 8-year-old fish are related to variability in recruitment 5–8 years earlier, we computed Pearson’s product moment correlations between the available time series of catch rates at each age and the recruitment time series at the appropriate lag. R version 2.12.2 (R Development Core Team 2011) was used for all statistical analyses.
Chronology of migration
Nine otoliths, collected in 1986 (n = 3), 1995 (n = 3), and 2005 (n = 3), were examined in this study. Otoliths were provided by L. Moulton of MJM Research (Lopez Island, Washington) and were drawn from an archive of otoliths collected from subsistence fisheries in the Colville River (von Biela et al. 2011). Prior to preparation for age and microchemical analysis, otoliths were rinsed in deionized water, air dried, and weighed. Each otolith was embedded in thermoplastic cement on a glass slide, ground in the sagittal plane, and polished. Individual otoliths were mounted sulcus side down and ground with 2000-grit wet-dry sandpaper on a glass plate to the level of the nucleus. The slide was then warmed on a hotplate and the otolith flipped with the sulcus side up. The otolith was again ground using 2000-grit wet-dry sandpaper until the primordia were exposed, polished with 12000-grit polishing cloth, and finally, polished with a slurry of 0.05 μm alumina and deionized water. Analysis of otolith Sr and Ca followed the methods of Zimmerman and Nielsen (2003) and Zimmerman (2005).
Otoliths were mounted on a petrographic slide (Donohoe and Zimmerman 2010), which was then rinsed with deionized water, air dried, and coated with a 40-nm carbon layer. Based on annual banding patterns, the fish examined ranged in age from 5 to 8 years old. Elemental analysis was conducted with a JEOL 8900 electron microprobe at the U.S. Geological Survey, Menlo Park, California. A 15-kV, 50-nA, 10-μm-diameter beam was used for all analyses. Strontianite and calcite were used as standards for Sr and Ca, respectively. Each element was analyzed simultaneously and a counting time of 40 s was used to maximize precision. A transect of points from the core to the edge of the otolith was sampled with a spacing of 14–25 μm between sampling points for each otolith. Sr/Ca ratios (molar) were plotted, and annuli were superimposed on these figures to describe the chronology of migration for each fish. Based on daily ring count, the mean number of days represented by a 10-μm-diameter sample point in these otoliths ranged from 5 days during the first year of life to over 30 days in years 6 through 8.
Results
Microsatellites
Eleven microsatellite loci were used to genotype a total of 678 Arctic cisco from Alaska and Canada (Online Resource 1). The number of microsatellite alleles (A) ranged from 15 (LGL BWF2) to 47 (Aut139; Table 1), with a mean A = 28.55. All microsatellite loci conformed to HWE when collections were combined. When sampling groups were analyzed individually, two loci deviated significantly from HWE in one sample collection each after Bonferroni correction (Aut139 in the Great Bear River collection had lower observed heterozygosity than expected, P < 0.0001; and Bom17c in the Colville 2006 collection had higher observed heterozygosity than expected, P = 0.0034). There was no evidence of inbreeding; global F IS = 0.007. No linkage disequilibrium was detected between loci for all collections combined. When sampling groups were analyzed individually, one pair of loci in the Colville 2007 sample collection showed significant evidence of linkage after Bonferroni correction (P = 0.0002). Allelic richness was similar for fish in the Colville River (average A R = 23.99) and Mackenzie River (average A R = 23.45) (Table 2). Private allele richness was 4.10 for fish in the Colville River and 3.56 for fish from the Mackenzie River system. Average observed and expected heterozygosity across all loci were 0.774 and 0.778, respectively. No statistically significant pairwise F ST differences were detected between sample collections (P ≥ 0.126) (Table 3) or major river drainages (P = 0.477). AMOVA results show that over 99.94 % of the allelic variation was found within individuals. The highest posterior probability (1.00) as determined using model-based clustering analyses occurred when K = 1 for genotypic data for all sample collections of Arctic cisco combined (Table 4) and when only genotypic data from the Mackenzie River system were considered (Table 4).
Mitochondrial DNA
Nineteen mtDNA haplotypes and 29 variable nucleotide positions were identified from the amplification and sequencing of a 594-bp fragment of ATPase 6 gene from 372 samples (Fig. 2; Table 5). Haplotype diversity among the 19 Arctic cisco haplotypes (AMN) was 0.414 ± 0.030 and nucleotide diversity averaged across all positions equaled 0.0011 ± 0.0009. Fish from the Colville River and Mackenzie River tributaries shared the five most common haplotypes (AMN001, AMN002, AMN006, AMN007, and AMN012). Colville River samples had 12 unique haplotypes found at low frequencies (1–2 fish each; <1 %) including two highly divergent haplotypes (AMN009 and AMN015). Two haplotypes found at low frequency (1 fish each) in the Arctic Red River sample collection (AMN010 and AMN011) were not identified in Colville River samples. No statistically significant difference was detected in haplotype frequencies for Arctic cisco from the Colville and Mackenzie rivers using pairwise F ST comparison (P = 0.678).

Minimum spanning network for mitochondrial DNA ATP synthase VI subunit gene haplotypes detected from putative Arctic cisco. Circles are drawn proportional to the frequency at which haplotypes were observed. Shading represents the sample location from which haplotypes originated: white (Colville River) and black (lower Mackenzie River system). A single mutation separates nodes unless indicated by number. Lines separating nodes are drawn to scale unless indicated by a break
Retrospective recruitment patterns
Retrospective analysis of recruitment confirmed previous research (Fechhelm et al. 2007), showing a correlation between year-class strength of Arctic cisco in the Colville River and wind conditions. Recruitment was best described by a linear model with a threshold below which wind speed did not affect recruitment (r 2 = 0.70, F = 54.3, P < 0.001) (Fig. 3). This implies that year-class recruitment failures occur in years with average July–August winds from the west and that recruitment increases linearly with the strength of easterly winds. Strong recruitment translated into elevated catch rates in the Colville River fishery 5–8 years later, with significant correlations (P < 0.05) between age-specific CPUE and the combined recruitment index ranging from 0.73 (age 7) to 0.86 (age 5) for commercial CPUE and from 0.60 (age 8) to 0.80 (age 6) for subsistence CPUE.

Relationship between average easterly wind speeds (m/s) and Arctic cisco recruitment index for the Colville River, Alaska, 1981–2005. The numbers on the plot indicate the recruitment class
Recruitment anomalies (i.e., residuals from the best-fit model) suggest that after accounting for the effects of wind, recruitment was anomalously low in the early 1980s, increased in the mid-1980s, and declined significantly since then with below-average recruitment in recent years (Fig. 4). No significant effects of climate conditions (near-shore temperatures or ice conditions) during early ocean life on these recruitment anomalies were found; thus, the variation in these anomalies remains unexplained.

Recruitment anomalies (i.e., residuals from the best-fit model) after accounting for the effects of wind given by year, Arctic cisco, Colville River, Alaska, 1981–2005
Age composition of Arctic cisco in the Colville River, estimated from aged samples obtained in the fishery between 1985 and 2004, ranged from 4 to 9 (mean 5.9). Arctic cisco captured in the Peel River in 1998 and 2002 ranged in age from 5 to 19 (mean 10.7; VanGerwen-Toyne et al. 2008). Fish captured in the Peel River were, therefore, older on average than Arctic cisco found in the Colville River.
Chronology of migration
Movement among different salinity environments for the nine Arctic cisco examined by otolith analysis in this study (Fig. 5) followed the life history model of seasonal movements between nearshore and coastal environments first presented by Gallaway et al. (1983) and Craig (1989). During the first year of life, otolith Sr/Ca ratios were low as juvenile fish presumably hatched and migrated downstream in their natal river and then increased as they entered saltwater and migrated west along the coast. Each year thereafter, otolith Sr/Ca ratios oscillated between lower and higher values indicating that fish moved between marine waters of high salinity during summer to areas of fresher or brackish waters to overwinter. Sr/Ca values associated with the first and second winter were often as low as Sr/Ca values associated with young-of-the-year freshwater residence (e.g., Fig. 5f, h), indicating that some Arctic cisco overwintered in freshwater in their first years.

Transects of otolith Sr/Ca, beginning in the otolith core and ending at the edge of the otolith for Arctic cisco captured in the Colville River, Alaska, in 1986, 1995, and 2005. The dashed line indicates the location of the otolith edge, and the solid gray lines indicate location of annuli, which corresponds to the transition from winter to spring
Discussion
Based on our reanalysis of genetics and recruitment, we find no evidence to refute the Mackenzie hypothesis. No genetic differentiation was detected between sample collections of Arctic cisco from the Colville River and the Mackenzie River system. Furthermore, no evidence for structuring was found using genotypic data derived from sample collections from tributaries of the lower Mackenzie River system. Recruitment of Arctic cisco to Alaska continues to be correlated with wind patterns, and year-class recruitment failures occur in years dominated by westerly winds, indicating that recent broad-scale changes to atmospheric circulation patterns have not changed sufficiently to preclude recruitment of Arctic cisco to Alaska. Further, Arctic cisco sampled from the Colville River appear to make repeated, annual migrations between low- and higher-salinity habitats in support of proposed models of Arctic cisco movement patterns.
Genetic analyses were conducted to assess population structure among sample collections of Arctic cisco from the Colville River and tributaries of the Mackenzie River. No statistically significant genetic differences were found between sample collections of fish from the two watersheds. Global F IS across all sample locations and years was low (0.007), suggesting minimal levels of inbreeding in Arctic cisco, adding additional support for a single large interbreeding population. Furthermore, model-based structuring methods support a single admixed population when considering genotypic data from all sample collections. Collectively, these genetic data do not refute the Mackenzie hypothesis and instead support the conclusion of previous investigations that fish spawning in the Mackenzie River are the source of Arctic cisco recruited into overwintering habitats within the Colville River (Bickham et al. 1989; Morales et al. 1993).
Genetic analyses of Arctic cisco from the Mackenzie River drainage did not support population differentiation among collections of fish made from putative spawning tributaries using standard population genetic statistics or model-based clustering methods. Thus, we found no support for genetic differences among spawning aggregations of Arctic cisco in the Mackenzie River system, contrary to a previous assessment (Bickham et al. 1989). Our results (i.e., lack of genetic differentiation among tributaries) suggest weak fidelity of Arctic cisco to natal streams for spawning within the lower Mackenzie River watershed.
Two highly divergent mtDNA haplotypes (AMN009 and AMN015) were found at low frequencies among the Colville River sample collections. These haplotypes could be from interspecific hybrids (Bernatchez and Dodson 1991; Bickham et al. 1997; Rogers et al. 2001) or from fish misidentified to species at the time of collection. Schlei et al. (2008) examined eight species of North American coregonids using DNA barcoding, and, of the 49 individuals analyzed, one fish morphometrically identified as a Bering cisco (Coregonus laurettae) was subsequently identified as an Arctic cisco using molecular techniques. Bering cisco are occasionally caught in the Colville River but not the Mackenzie River and were once considered to be conspecific with Arctic cisco (McPhail 1966).
Approximately 70 % of the variation in recruitment of juvenile Arctic cisco to Alaskan waters was determined by the strength of easterly winds during summer (July–August), leading to variations in catch rates in the fishery 5–8 years later. This supports the hypothesis that Arctic cisco captured in Alaska originate from the east and presumably from the Mackenzie River. Analysis of age structure further supports the hypothesis that Arctic cisco only rear in Alaska rivers and coastal waters until they reach maturity and return to the Mackenzie River. If local populations were established in Alaska rivers, we would have expected to capture fish of older ages as we saw in Mackenzie River tributaries.
The chronology of migration between freshwater and saltwater for Arctic cisco generally follows the conceptual model proposed by Gallaway et al. (1983) and Craig (1989) based on otolith microchemistry; although, it appears that Arctic cisco may be overwintering in lower salinity habitats during their first winters at sea. For example, at ages two and three (Fig. 5b), otolith Sr/Ca ratios decline to values similar to those observed in presumed freshwater rearing during the first year of life in the river, suggesting some fish overwintered in low-salinity habitats. Previous capture data indicate that Arctic cisco overwinter in areas of moderate salinity (~5 ppt) (Schmidt et al. 1989).
Otolith Sr/Ca ratios measured in Arctic cisco were greater than those reported for other Salmonines. For example, molar ratios of otolith Sr/Ca measured in this study ranged from 0.0005 to 0.0056, while a range of 0.0009–0.0034 was reported for several salmonids exposed to salinities ranging from freshwater (salinity = 0 psu) to full seawater (salinity = 33 psu; Zimmerman 2005). It is unclear why otolith Sr/Ca ratios can be so much higher in Arctic cisco, but variation among species can result from different physiological pathways and filters (Campana 1999) and different responses to interactions among environmental variables (Elsdon and Gillanders 2003, 2004). Otolith Sr/Ca ratios associated with overwinter periods increased with fish age, indicating that older fish overwinter in higher-salinity habitats. Because otolith growth slows as fish age, however, the temporal resolution of the 10-μm-diameter electron beam used to measure Sr/Ca ratios also decreases with the age of the fish. As a result, measurement points taken in the first years of life discriminate salinity over a range of only a few days, a time period that is likely to reflect occupation of a salinity-specific habitat. In contrast, measurements taken during age-8 cover a mean of 35 days and, thus, are more likely to integrate movement among different salinity habitats. This variation in temporal resolution among ages makes it more difficult to establish overwinter salinities at older ages. Otolith analyses indicated that Arctic cisco reside in brackish waters throughout the winter leading to additional questions about their overwintering physiology. It is not clear if fish experience yearly resmoltification and undergo physiological changes necessary for freshwater/salt water adaptation in their fall and spring transitions to and from the Beaufort Sea (see similar life history traits described in anadromous Arctic char Salvelinus alpinus by Aas-Hansen et al. 2005).
Climate conditions in the Arctic have undergone significant changes, particularly during the last 20 years (Walsh 2008; Overland et al. 2008; Overland and Wang 2010). These changes include increased warming (Overland et al. 2008) and increased storminess and shifts in atmospheric circulation patterns (Overland and Wang 2010). In spite of these changes, Arctic cisco recruitment patterns are still best described by the Mackenzie hypothesis. How Arctic cisco populations respond to future changes remains a critical question. Murphy et al. (2007) found weak effects of spring and summer temperature conditions on overwinter survival of Arctic cisco, suggesting that survival of young-of-the-year and sub-adults was reduced during summers with above-averaged temperatures. Similarly, von Biela et al. (2011) found that young-of-the-year growth rates of Arctic cisco were strongly positively related to temperature and river flow. Ultimately, Arctic cisco growth, survival, and recruitment dynamics are closely related to a set of environmental variables that are likely to change dramatically over the next 50–100 years. Continued monitoring and assessment of existing theory is needed to ensure continued conservation of this important subsistence resource.
References
Aas-Hansen Ø, Vijayan MM, Johnson HK, Cameron C, Jørgensen EH (2005) Resmoltification in wild, anadromous Arctic char (Salvelinus alpinus): a survey of osmoregulatory, metabolic, and endocrine changes preceding annual seawater migration. Can J Fish Aquat Sci 62:195–204
Bandelt H-J, Forester P, Röhl A (1999) Median-joining networks for inferring intraspecific phylogenies. Mol Biol Evol 16:37–48
Bernatchez L, Dodson JJ (1991) Phylogeographic structure in mitochondrial DNA of lake whitefish (Coregonus clupeaformis) and its relation to Pleistocene glaciations. Evolution 45:1016–1035. doi:10.2307/2409706
Bickham JW, Carr SM, Hanks BG, Barton DW, Gallaway BJ (1989) Genetic analysis of population variation in the Arctic cisco (Coregonus autumnalis) using electrophoretic, flow cytometric, and mitochondrial DNA restriction analyses. Biol Pap Univ Alask 24:112–122
Bickham JW, Patton JC, Minzenmayer S, Moulton LL, Gallaway BJ (1997) Identification of Arctic and Bering ciscoes in the Colville river delta, Beaufort Sea Coast, Alaska. Am Fish Soc Symp 19:224–228
Bond WA, Erickson RN (1997) Coastal migrations of Arctic cisco in the Eastern Beaufort Sea. In: Fish ecology in Arctic North America. Am Fish Soc Symp 19:155–164
Brown RJ, Bickford N, Severin K (2007) Otolith trace element chemistry as an indicator of anadromy in Yukon River drainage coregonine fishes. Trans Am Fish Soc 136:678–690
Campana SE (1999) Chemistry and composition of fish otoliths: pathways, mechanisms and applications. Mar Ecol Prog Ser 188:263–297
Colonell JM, Gallaway BJ (1997) Wind-driven transport and dispersal of age-0 Arctic cisco along the Beaufort Sea coast. Am Fish Soc Symp 19:90–103
Craig PC (1984) Fish use of coastal waters of the Beaufort Sea: a review. Trans Am Fish Soc 113:265–282
Craig PC (1989) An introduction to anadromous fishes in the Alaskan arctic. Biol Pap Univ Alask 24:27–54
Donohoe CJ, Zimmerman CE (2010) A method of mounting multiple otoliths for beam-based microchemical analyses. Environ Biol Fish 89:473–477. doi:10.1007/s10641-010-9680-3
Elsdon TS, Gillanders BM (2003) Reconstructing migratory patterns of fish based on environmental influences on otolith chemistry. Rev Fish Biol Fish 13:219–235
Elsdon TS, Gillanders BM (2004) Fish otolith chemistry influenced by exposure to multiple environmental variables. J Exp Mar Biol Ecol 313:269–284
Excoffier L, Laval G, Schneider S (2005) ARLEQUIN version 3.0: an integrated software package for population genetics data analysis. Evol Bioinform 1:47–50
Fechhelm RG, Fissel DB (1988) Wind-aided recruitment of Canadian Arctic cisco (Coregonus autumnalis) into Alaskan waters. Can J Fish Aquat Sci 45:906–910
Fechhelm RG, Griffiths WB (1990) Effect of wind on the recruitment of Canadian Arctic cisco (Coregonus autumnalis) into the Central Alaskan Beaufort Seas. Can J Fish Aquat Sci 47:2164–2171
Fechhelm RG, Streever B, Gallaway BJ (2007) The Arctic cisco (Coregonus autumnalis) subsistence and commercial fisheries, Colville River, Alaska: a conceptual model. Arctic 60:421–429
Gallaway BJ, Griffiths WB, Craig PC, Gazey WJ, Helmericks JW (1983) An assessment of the Colville River Delta stock of Arctic cisco—migrants from Canada? Biol Pap Univ Alask 21:4–23
Gallaway BJ, Gazey WJ, Moulton LL (1989) Population trends for the Arctic cisco (Coregonus autumnalis) in the Colville River of Alaska as reflected by the commercial fishery. Biol Pap Univ Alask 24:153–165
Giuffra E, Bernatchez L, Guyomard R (1994) Mitochondrial control region and protein coding gene sequence variation among phenotypic forms of brown trout Salmo trutta from northern Italy. Mol Ecol 3:161–171. doi:10.1111/j.1365-294X.1994.tb00117.x
Goudet J (2001) FSTAT, a program to estimate and test gene diversities and fixation indices (version 2.9.3). Available: http://www2.unil.ch/popgen/softwares/fstat.htm
Hubisz MJ, Falush D, Stephens M, Pritchard JK (2009) Inferring weak population structure with the assistance of sample group information. Mol Ecol Res 9:1322–1332
Kalinowski ST (2005) HP-RARE 1.0: a computer program for performing rarefaction on measures of allelic richness. Mol Ecol Notes 5:187–189. doi:10.1111/j.1471-8286.2004.00845.x
Lockwood SF, Bickham JW (1991) Genetic stock assessment of spawning Arctic cisco (Coregonus autumnalis) populations by flow cytometric determination of DNA content. Cytom Part A 12:260–267. doi:10.1002/cyto.990120309
MBC Applied Environmental Sciences (2004) Proceedings of a workshop on the variability of Arctic cisco (Qaaktaq) in the Colville River. OCS Study MMS 2004-033. Unpublished report prepared for the U.S. Dept. of the Interior, Minerals Management Service, Alaska OCS Region, Anchorage, AK
McPhail JD (1966) The Arctic cisco complex in Alaska and Northwestern Canada. J Fish Res Bd Can 23:141–148
Miya M, Nishida M (2000) Use of mitogenomic information in teleostean molecular phylogenetics: a tree based exploration under the maximum-parsimony optimality criterion. Mol Phylogenet Evol 17:437–455
Morales JC, Hanks BG, Bickham JW, Derr JN (1993) Allozyme analysis of population structure in Arctic cisco (Coregonus autumnalis) from the Beaufort Sea. Copeia 1993:863–867
Moulton LL (1989) Recruitment of Arctic cisco (Coregonus autumnalis) into the Colville Delta, Alaska. Biol Pap Univ Alask 24:107–111
Murphy SM, Mueter FJ, Braund SR, Lampe L, Prichard AK, Anderson BA, Seigle JC (2007) Variation in the abundance of Arctic cisco in the Colville River: analysis of existing data and local knowledge. Volumes I and II: Final Report. Unpublished report by ABR, Inc. to U. S. Department of the Interior, Minerals Management Service (MMS), Alaska Outer Continental Shelf Region, Anchorage, AK. Technical Report No. MMS 2007-042. Available from ABR, Inc. P.O. Box 80410, Fairbanks, Alaska 99708-0410, USA
Oetting WS, Lee HK, Flanders DJ, Wiesner GL, Sellers TA, King RA (1995) Linkage analysis with multiplexed short tandem repeat polymorphisms using infrared fluorescence and M13 tailed primers. Genomics 30:450–458
Overland JE, Wang M (2010) Large-scale atmospheric circulation changes are associated with the recent loss of Arctic sea ice. Tellus 62A:1–9
Overland JE, Wang M, Salo S (2008) The recent Arctic warm period. Tellus 60A:589–597
Patton JC, Gallaway BJ, Fechhelm RG, Cronin MA (1997) Genetic variation of microsatellite and mitochondrial DNA markers in broad whitefish (Coregonus nasus) in the Colville and Sagavanirktok rivers in northern Alaska. Can J Fish Aquat Sci 54:1548–1556
Peakall R, Smouse PE (2006) GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Mol Ecol Notes 6:288–295. doi:10.1111/j.1471-8286.2005.01155.x
Pritchard JK, Stephens M, Donnelly P (2000) Inference of population structure using multilocus genotype data. Genetics 155:945–959
R Development Core Team (2011) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Australia
Ramey A, Graziano SL, Nielsen JL (2008) Isolation and characterization of eight microsatellite loci from the Arctic cisco, Coregonus autumnalis. Mol Ecol Resour 8:357–359
Reist JD, Bond WA (1988) Life history characteristics of migratory coregonids of the lower Mackenzie River, Northwest Territories, Canada. Finn Fish Res 9:133–144
Rogers SM, Campbell D, Baird SJE, Danzmann RG, Bernatchez L (2001) Combining the analyses of introgressive hybridization and linkage mapping to investigate the genetic architecture of population divergence in lake whitefish (Coregonus clupeaformis, Mitchill). Genetica 111:25–41
Rogers SM, Marchand MH, Bernatchez L (2004) Isolation, characterization, and cross-salmonid amplification of 31 microsatellite loci in the lake whitefish (Coregonus clupeaformis Mitchill). Mol Ecol Notes 4:89–92. doi:10.1046/j.1471-8286.2003.00578.x
Schlei OL, Crête-Lafrenière A, Whiteley AR, Brown RJ, Olsen JB, Bernatchez L, Wenburg JK (2008) DNA barcoding of eight North American coregonine species. Mol Ecol Resour 8:1212–1218. doi:10.1111/j.1755-0998.2008.02350.x
Schmidt DR, Griffiths WB, Martin LR (1989) Overwintering biology of anadromous fish in the Sagavanirktok River delta, Alaska. Biol Pap Univ Alask 24:55–74
Schmidt DR, Griffiths WB, Beaubien DK, Herlugson CJ (1991) Movement of young-of-the-year C. autumnalisArctic cisco across the Beaufort Sera, 1985-1988. Am Fish Soc Symp 11:132–144
Sokal RR, Rohlf FJ (1995) Biometry. WH Freeman and Company, New York
Turgeon J, Bernatchez L (2001) Mitochondrial DNA phylogeography of lake cisco (Coregonus artedi): evidence supporting extensive secondary contacts between two glacial races. Mol Ecol 10:987–1001
Turgeon J, Bernatchez L (2003) Reticulate evolution and phenotypic diversity in North American ciscoes, Coregonus ssp. (Teleostei: Salmonidae): implications for the conservation of an evolutionary legacy. Conserv Genet 4:67–81
Turgeon J, Estoup A, Bernatchez L (1999) Species flock in the North American Great Lakes: molecular ecology of Lake Nipigon ciscoes (Teleostei: Coregonidae: Coregonus). Evolution 53:1857–1871. doi:10.2307/2640446
VanGerwen-Toyne M, Walkser-Larson J, and Tallman RF (2008) Monitoring spawning populations of migratory coregonids in the Peel River, NT: The Peel River Study 1998-(2002) Canadian Manuscript Report of Fisheries and Aquatic Sciences 2851
von Biela VR, Zimmerman CE, Moulton LL (2011) Long-term increases in young-of-the-year growth of Arctic cisco Coregonus autumnalis and environmental influences. J Fish Biol 78:39–56. doi:10.111/j.1095-8649.2010.02832.x
von Biela VR, Zimmerman CE, Cohn BR, Welker JM (2013) Terrestrial and marine trophic pathways support young-of-year growth in a nearshore Arctic fish. Polar Biol 36:137–146
Walsh JE (2008) Climate of the Arctic marine environment. Ecol Appl 18:S3–S22
Zimmerman CE (2005) Relationship of otolith strontium-to-calcium ratios and salinity: experimental validation for juvenile salmonids. Can J Fish Aquat Sci 62:88–97
Zimmerman CE, Nielsen RL (2003) Effects of analytical conditions in wavelength dispersive electron microprobe analysis on the measurement of strontium-to-calcium (Sr/Ca) ratios in otoliths of anadromous salmonids. Fish Bull 101:712–718
Zimmerman CE, Stonecypher RW, Hayes MC (2003) Migration of precocious male hatchery Chinook salmon in the Umatilla River, Oregon. N Am J Fish Manag 23:1007–1015
Acknowledgments
Sara Graziano, Sean Burril, and Vanessa von Biela (U.S. Geological Survey; USGS) provided significant contributions to laboratory and statistical analyses and fish collections on the Mackenzie River. Sample collections for this report were made available by Larry Moulton (MJM Research); Kate Wedemeyer (Bureau of Ocean Energy Management); BP Exploration, Inc.; ConocoPhillips, Alaska; and the U. S. Fish and Wildlife Service. USGS sampling in Canada was facilitated by the Department of Fisheries and Oceans Canada and the Aurora Research Institute. Hospitality and assistance from citizens of Fort Good Hope, Norman Wells, and Inuvik was greatly appreciated. Partial funding for this study came from the USGS Outer Continental Shelf Program. We thank three anonymous reviewers and the editor for very helpful comments. Any use of trade names or products is for descriptive purposes only and does not imply endorsement of the US Government.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Zimmerman, C.E., Ramey, A.M., Turner, S.M. et al. Genetics, recruitment, and migration patterns of Arctic cisco (Coregonus autumnalis) in the Colville River, Alaska, and Mackenzie River, Canada. Polar Biol 36, 1543–1555 (2013). https://doi.org/10.1007/s00300-013-1372-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-013-1372-y