
Abstract
Glacier surfaces have a surprisingly complex ecology. Cryoconite holes contain diverse invertebrate communities, while other invertebrates, such as Collembola, often graze on algae and windblown dead organic material on the glacier surface. Glacier mice (ovoid unattached moss balls) occur on some glaciers worldwide. Studies of these glacier mice have concentrated on their occurrence and mode of formation. There are no reports of the invertebrate communities. But, such glacier mice may provide a suitable favourable habitat and refuge for a variety of invertebrate groups to colonise the glacier surface. Here, we describe the invertebrate fauna of the glacier mice (moss balls) of the Falljökull, Iceland. The glacier mice were composed of Racomitrium sp. and varied in size from 8.0 to 10.0 cm in length. All glacier mice studied contained invertebrates. Two species of Collembola were present. Pseudisotoma sensibilis (Tullberg, 1876) was numerically dominant with between 12 and 73 individuals per glacier mouse, while Desoria olivacea (Tullberg, 1871) occurred but in far lower numbers. Tardigrada and Nematoda had mean densities of approximately 200 and 1,000, respectively. No Acari, Arachnida or Enchytraeidae were observed, which may be related to the difficulty these groups have in colonising the glacier mice. We suggest that glacier mice provide an unusual environmentally ameliorated microhabitat for an invertebrate community dwelling on a glacial surface. The glacier mice thereby enable an invertebrate fauna to colonise an otherwise largely inhospitable location with implications for carbon flow in the system.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Glacier surfaces are often considered barren and largely devoid of life. But this assertion is beginning to be challenged with the observation of glacier fleas such as Desoria albicornis (Fjellberg 2010), ice worms, for example, Mesenchytraeus solifugus (Hartzell et al. 2005), and the diverse fauna and flora of cryoconite holes (Wharton et al. 1985; De Smet and Van Rompu 1994). Moreover, the importance of these ecosystems to nutrient fluxes is becoming appreciated (Hodson et al. 2005; Anesio et al. 2009). A new addition to this list is the fauna of the glacier mouse or jökla-mýs. Glacier mice (jőkla-mýs of Eythórsson 1951), whether termed the unattached moss polsters of Shacklette (1966) or the supraglacial globular moss cushions of Porter et al. (2008), are ovate balls of moss found on the surface of a few glaciers distributed throughout the world including Iceland, North and South America, and the Himalaya (Eythórsson 1951; Heusser 1972; Perez 1991; Porter et al. 2008). Such mice are comprised of moss balls lying on the glacier surface. Moss is well known to harbour a diverse invertebrate community and may form an especially important habitat in the extreme environments of Arctic regions where moss vegetation may often dominate (Jonsdottir 2005). Consequently, these glacier mice might be expected to possess a characteristic invertebrate fauna. Nonetheless, study to date of glacier mice has largely focused on the physical composition and the mode of formation (Eythórsson 1951; Heusser 1972; Perez 1991; Porter et al. 2008), and the associated faunal constituent has been ignored.
Typically, glacier mice are small balls of moss up to 10 cm in length, often ovate and with a pronounced roundness. They appear to form when moss begins to establish around a clast lying on the glacier surface. The moss continues to grow and in time insulates the glacier surface resulting in the moss becoming elevated on a pedestal as the surrounding ice melts. Eventually, the moss falls from this pedestal (Porter et al. 2008). In many cases, the glacier mouse is lenticular in form with a pronounced flatter lower side but movement across the glacier surface enables the glacier mouse to achieve a rounded form (Shacklette 1966). The formation of the mice appears to be a result of the unusual environment rather than specific species of moss. Glacier mice are comprised of a wide range of moss species including Drepanocladius berggrenii (Heusser 1972), Grimmia longirostris (Perez 1991), Schistidium apocarpum (Shacklette 1966) and Racomitrium fasciculare and R. ericoides (Porter et al. 2008). With a high organic content and fine silt accumulated by trapping aeolian dust, the glacier mice have a great water-holding ability (Perez 1991). This moist organic environment potentially provides a suitable habitat for many species of invertebrate. For example, Rotifera, Tardigrada, Acari and Collembola are all known to inhabit mosses in other Arctic regions such Svalbard (European High Arctic) (Coulson 2007 and references there in; Dastych 1985; De Smet et al. 1988; De Smet and Van Rompu 1994).
Invertebrates are recognised to exploit habitats on the surface of ice. Collembola are known from glacier surfaces (Kopeszki 2000; Fjellberg and Bernard 2009; Fjellberg 2010). Enchytraeid worms, “ice worms” (Hartzell and Shain 2009), are observed inhabiting the upper centimetres of the glacial ice of a number of glaciers in Alaska and the Himalaya (Hartzell et al. 2005; Hartzell and Shain 2009). Moreover, cryoconite holes contain diverse communities including Protozoa, Rotifera and Tardigrada (Wharton et al. 1985; Säwström et al. 2002; Porazinska et al. 2004). Nonetheless, glaciers on the whole provide a poor habitat for soil microarthropods being cold, exposed and, for the most part, devoid of food resources. Glacier mice possibly offer a potential habitat for the invertebrate colonisation of local regions of the glacier surface feeding on ice algae and allochthonous organic debris. Moreover, in addition to providing a habitat in themselves, they create a potential refuge enabling animals foraging on the glacier surface to periodically retreat to shelter and hence exploit a greater area of the glacier surface. Since these glacier mice can be redistributed across the surface of the glacier via the action of wind, water and movement of the ice (Porter et al. 2008), they may also offer a means of limited dispersal across a generally hostile surface while remaining within a favourable microhabitat. Nevertheless, the invertebrate fauna inhabiting this novel microhabitat has not attracted attention. We here describe the invertebrate fauna of glacier mice from the Falljökull glacier in Iceland and consider their importance to glacier ecology.
Materials and methods
Field site
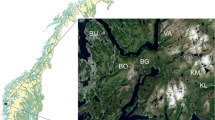
Falljökull is an outlet glacier of Öræfajökull, which is part of the larger Vatnajökull in south-east Iceland (the terminus is located c. 63°58′N 16°48′W, Fig. 1). Falljökull descends from the high plateau of Öræfajökull down a steep highly-crevassed icefall with around 1.5 km length of largely crevasse-free glacier snout below the icefall and the terminal margin adjoining the adjacent Virkisjökull. The terminus of Falljökull is predominantly debris-free ice, with the exception of the south-east lateral margin of the glacier, which has a thin supraglacial debris cover that is laterally extensive. The north-west lateral margin of Falljökull adjoins the adjacent Virkisjökull, which also has a thin supraglacial debris cover that is laterally extensive along its south-east lateral margin. Dead ice features in the proglacial area indicate that both Falljökull and Virkisjökull are currently experiencing rapid recession of the ice front. The merged Virkisjökull and Falljökull complex was at its Neoglacial maximum as early as A.D. 1740 according to Chenet et al. (2010), but the lichenometric dating studies have proved controversial (Dąbski 2010; Chenet et al. 2011). Recession of around 1.5 km has occurred since this Neoglacial maximum.

Location of the Falljökull, Iceland, with sampling site indicated
Climate
The climate of the area is characterised by high precipitation and also by higher temperatures than a position adjacent to the arctic circle might imply. The nearest Icelandic Meteorological Office weather station to Falljökull is at Fagurhólsmýri with a monthly temperature and precipitation data set from 1949 to 2007 (Fig. 2). From 1949 to 2007 at Fagurhólsmýri, the mean annual precipitation was 1,814 mm and the mean annual temperature was 4.8 °C.

Climate data from 1949 to 2007 at the Fagurhólsmýri weather station (data supplied by the Icelandic Meteorological Office). Mean monthly temperature (solid line), mean monthly precipitation (bars)
Glacier mice characteristics
Five Onset HOBO Pendant G data loggers were each placed within a glacier mouse to measure mouse motion on an area of the glacier with an overall slope angle of c. 10°. The Pendant G data logger records combined x-axis, y-axis and z-axis acceleration (g) and tilt (°), so can be used to detect motion. The stated accuracy of the logger is ±0.075 g at 25 °C and ±0.105 g at −20 to +70 °C with a resolution of 0.025 g. The size of each data logger (58 × 33 × 23 mm) meant that larger glacier mice were preferentially selected for observation with the aim to minimise the impact that the addition of the data logger would have on glacier mouse motion. A 30-s logging interval was used for the duration of the logging period.
The acceleration values (g) from the three axes were used to obtain a single change in angle value (θ°) using the following dot product formula from one vector to the next:
A ternary plot (Graham and Midgley 2000) was employed to describe the shape (Fig. 3). This plot describes the full continuum of shape possibilities from equidimensional to oblate or prolate. Inspection of this plot indicates a clear tendency towards an equidimensional character with no apparent differences in shape between those glacier mice with accelerometers (diamond symbols), those extracted for the invertebrate fauna (square symbols) and the glacier mouse used to assess temperature characteristics (triangle symbol).

Ternary diagram (Graham and Midgley 2000) describing the full continuum of glacier mice shape possibilities from: top a = b = c = 1 equidimensional; bottom left a = b = 1 and c = 0 oblate; bottom right a = 1 and b = c = 0 prolate. Square symbols indicate the glacier mice extracted for the invertebrate fauna, diamond symbols the accelerometer samples and the triangle symbol the glacier mouse with the temperature record
An Onset HOBO Pro V2 temperature logger was used to measure air temperature at the frontal margin of Falljökull between 27 July until 12 August 2010. There is a gap of 1 day in the data due to logger malfunction. The air temperature logger was mounted within a solar radiation shield at 1.25 m above the glacier surface. An external probe from the temperature logger was inserted centrally within the core of a single glacier mouse and used to measure internal glacier mouse temperature at the site. A 60-s logging interval was used for both air and glacier mouse temperature measurements.
Invertebrate extraction
Ten glacier mice were sampled from the surface of the Falljökull close to the terminus on 29 July 2010 (Fig. 4a, b) from an area under 10 m2 and returned to the University Centre in Svalbard (UNIS), Longyearbyen, Svalbard, Norway. The microarthropod fauna of eight mice was extracted in Tullgren funnels, while that of the remaining two mice was extracted in Baermann funnels to collect the Tardigrada, Enchytraeidae and Nematoda. The Collembola are deposited in the reference collection at UNIS.

a The glacier mice of the Falljökull, Iceland 2007, b glacier mouse FJ-2010-03
Age classes of the Collembola
The lengths of the extracted Collembola were measured under a Leica MZ16 stereomicroscope to determine age classes.
Moisture content
After the extraction of the invertebrate fauna, the mice were placed in a drying oven at 70 °C for 24 h until thoroughly dry. Moisture content (ρ) was calculated as (wet weight − dry weight)/dry weight.
Statistics
Spearman correlation and linear regression were performed using SigmaPlot v. 11 (Systat Software Inc.) to determine relationships between size, weight, moisture content and total numbers of Collembola. Collembola were not analysed by species due to the overwhelming dominance of one species. Samples extracted using Baermann funnels were not inspected statistically due to the n size of two.
Results
Invertebrates
Two species of Collembola were found in the glacier mice; Pseudisotoma sensibilis (Tullberg, 1876) and Desoria olivacea (Tullberg, 1871) (Table 1). Pseudisotoma sensibilis dominated the Collembola with numbers per mouse varying between 0 and 73 individuals. Desoria olivacea was represented by only three individuals from the eight mice extracted in the Tullgren funnels. The age classes of P. sensibilis are presented in Fig. 5. Two peaks in size classes are present with a juvenile cohort centred on 1.0 mm and an adult peak at 2.6 mm.

Size classes of P. sensibilis. Size classes of 0.4 mm with bars centred on middle of each size class
Tardigrada were common in the two glacier mice wet extracted with approximately 200 individuals in both samples. While no Enchytraeidae were found, Nematoda were common with over 1,000 individuals in mouse FJ-2010-02 (Table 1). A small number of Collembola were collected as a by-catch during the wet extractions.
Physical environment of the glacier mice
The mice were composed almost completely of the moss Racomitrium with very little organic soil. It was not possible to determine which species of moss comprised the glacier mice due to the unusual growth form of the moss into the ovoid mice (Figs 3, 4a, b). The mice varied in size from 5.4 to 12.1 cm long and a wet weight from 64.3 to 468.5 g (Table 1). Water comprised typically around 50 % of the wet weight of the mice (Table 1). No statistically significant relationships, or relationships approaching significance, were observed between total Collembola numbers and wet weight, dry weight, volume or moisture content (p > 0.05).
The glacier mouse temperature has a maximum recorded temperature of 12.4 °C and a minimum recorded temperature of 1.5 °C, but typical glacier mouse temperature ranged from just over 2 to around 6 °C. Glacier mouse temperature was predominantly lower than air temperature during the observation period. On a single occasion, when the glacier mouse temperature rose to the recorded maximum of 12.4 °C, it was 1.5 °C warmer than the surrounding air temperature at the time. Typical air temperature ranged from around 6 to 10 °C but displayed strong diurnal variation with a maximum recorded temperature of 14.7 °C and a minimum recorded temperature of 5.3 °C (Fig. 6).

Air temperature (solid line) and internal glacier mouse temperature (dotted line). Data are missing for the period 6 August due to logger malfunction
Movement of the glacier mice
Three types of glacier mouse motion are illustrated by the accelerometer data sets: (1) stick; (2) creep; and (3) roll. The stick motion behaviour type only appears after the fresh placement of a glacier mouse and probably only occurs following relocation to a fresh ice surface. The creep motion type is of minimal important for motion, whereas the roll motion type is the most significant in terms of glacier mouse movement.
Two types of creep are identified. Type 1 creep (roll build-up) occurs immediately prior to a roll with a gradual increase in the rate of rotation from close to 0° to over 6° per hour. This is followed by a roll of the moss ball. Type 2 creep (without roll) again shows a build-up similar to that preceding a roll and elevated rotation rates occur over around 90 min with rotation of up to 15° per hour observed. This form of rotation is not followed by a subsequent roll.
The minimum time before a roll occurred was only 12.2 h with a resulting roll of 41.8°. The maximum time before a roll occurred was 65.6 h with a resulting roll of 30.1°. The biggest single roll that occurred was 154.8°. Typically roll events occur after 12 to 40 h and are between around 30° to 60° of rotation. While some glacier mice did not exhibit any roll events during the observation period, a total of 5 roll events were recorded for a single glacier mouse over a seven day observation period (Fig. 7). While each roll is the rotation observed within a 30 s time window, the rotation is likely to occur over a period of a few seconds at most.

Vector of rotation of one glacier mouse. Glacier mouse roll events over a 7-day observation period
Discussion
In the glacier, mice from Falljökull three invertebrate groups were identified, Collembola, Tardigrada and Nematoda. Nonetheless, and despite the apparent suitability of the habitat for soil invertebrates, the fauna observed was species poor. Although it should be recognised that the fauna sampled, and described here, is partly a function of the extraction techniques employed. Extraction efficiency of differing taxa also varies with extraction procedure (Southwood and Henderson 2000) and there will be some unavoidable bias in the results. Only two species of Collembola were present despite 149 species being recorded from Iceland as a whole (Fjellberg 2007a). The Collembola identified are both common Holarctic species (Babenko and Fjellberg 2006; Fjellberg 2007b). Tardigrada and Nematoda were numerous in the two glacier mice wet extracted but these were not identified to species. No Enchytraeidea were found, nor were there any Acari or Araneae that might have been expected. The lack of Acari was particularly surprising. Acari are well known from moss habitats in other regions (Krantz and Walter 2009), and the Oribatidae are often referred to as ‘moss mites’ (Walter and Procter 1999). However, their absence, as well as that of the Enchytraeidae and Aranaea, may well be accounted for by the inherent difficulty of colonising small isolated ephemeral habitats on the glacier surface.
The moss balls form at isolated supraglacial outcrops from clasts and the aeolian deposition of sediment. However, glacier mice are not observed on all glaciers and their development is likely dependent on the presence of both suitable supraglacial material and the meteorological conditions (Fig. 2), which enable moss growth. Given these often remote and inaccessible growth locations, it seems likely that the initial invertebrate colonisation route is a random wind dispersal event. It is appreciated, or speculated, that accidental anemochory may be important for the colonisation of new habitats by some invertebrate groups such as Collembola, spiders and mites (Pugh and McInnes 1998; Gjelstrup 2000; Hawes et al. 2007). The lack of Enchytraeidae in the glacier mice may be explained by potential difficulties of this taxon in colonising the isolated supraglacial outcrops via wind dispersal.
The glacier mice provide a characteristic environment; moist, relatively warm and with a ready food source. Although anhydrobiotic Tardigrada are suspected of dispersing great distances in the Arctic via wind dispersal (Pugh and McInnes 1998), desiccation susceptible taxa such as Collembola (Block et al. 1990; Hodkinson et al. 1994; Makkonen et al. 2011) may face a greater challenge. Collembola are recognised to exploit the surfaces of glaciers (Fjellberg 2010) and glacier mice will provide these animals with a habitat on the largely inhospitable glacier surface from which they can emerge to graze on algae and deposited organic material. Within the glacier mice, temperatures rarely attain air temperature. This is in stark contrast to other habitats in the Arctic where ground temperatures may attain temperatures considerably above air temperature (Coulson et al. 1993; Scherrer and Korner 2010). This seemingly anomalous result is likely due to the high specific heat capacity of water and the high moisture content of the moss thermally buffering the glacier mice against the diurnal swings, the low angle of the sun at the moderately high latitude of just under 64°N and consequent reduced solar insolation per unit ground area and, finally, close contact with the ice of the glacier surface. However, despite the temperature of the glacier mouse being substantially colder than that of the air, the internal temperature of the glacier mouse is nonetheless far greater than that of the glacier surface at approximately 0 °C. Hence, compared with the glacier surface, the glacier mouse provides a thermally ameliorated environment. It must also be appreciated that thermal input for the glacier mice must come from a combination of solar radiation and precipitation (as rain). Input of warm rain is interpreted to be the cause of the highest glacier mouse temperature. Hence, during the summer period, although cooler than air temperature, the microhabitat within the glacier mice is considerably warmer than that of the surface of the glacier. Consequently, the glacier mice provide a thermally advantageous microhabitat amid the more hostile landscape.
Body length of Collembola is often used as a proxy measure for individual age (Birkemoe and Sømme 1998; Birkemoe and Leinaas 1999). While some care must be employed in interpreting such data since Collembola with poor food resources can display the phenomenon of degrowth (Hopkin 1997), body size does nonetheless provide a useful tool by which to observe age classes and elucidate life histories (Birkemoe and Sømme 1998; Birkemoe and Leinaas 1999). In addition, the two peaks we observed here may be the result of random dispersal/colonisation processes of windblown specimens. The numerically abundant small juveniles may be more easily carried away from the source area to the glacier mice than the larger size classes rather than being hatched in the glacier mice. But, the two peaks in body length of P. sensibilis indicate the presence of adults and juveniles strongly suggesting a reproducing population. It is therefore reasonable to assume that the glacier mice are exploited as more than just a temporary refuge, rather that the mice harbour resident populations.
The glacier mouse may also provide an additional advantage for the inhabitants. The ovoid shape of the glacier mice is a result of the gradual rolling motion of the mice. The distances moved by the glacier mice, either self-induced via growth imbalances or wind action, are unknown. However, there is a clear potential for redistribution on the glacier surface, although the main axis of movement is likely to be down the prevailing slope towards the glacier snout (Porter et al. 2008).
Glacier mice therefore form a novel, if limited, glacial habitat for invertebrate faunas from a range of groups. For taxa such as Collembola, glacier mice may provide a refuge from the extreme environment of the ice surface for individuals venturing out to exploit the organic material and algae the glacial surface as a food resource. Moreover, the glacial mice provide a semi-permanent habitat for other taxa such as Nematoda and Tardigrada.
References
Anesio AM, Hodson AJ, Fritz A, Psenner R, Sattler B (2009) High microbial activity on glaciers: importance to the global carbon cycle. Glob Change Biol 15:955–960
Babenko A, Fjellberg A (2006) Collembola Septentrionala. A catalogue of springtails of the Arctic regions. KMK Scientific Press Ltd, Moscow
Birkemoe T, Leinaas HP (1999) Reproductive biology of the Arctic collembolan Hypogastrura tullbergi. Ecography 22:31–39
Birkemoe T, Sømme LS (1998) Population dynamics of two collembolan species in an Arctic tundra. Pedobiologia 42:131–145
Block W, Harrisson PM, Vannier G (1990) A comparative-study of patterns of water-loss from two Antarctic springtails (Insecta, Collembola). J Insect Physiol 36:181–187
Chenet M, Roussel E, Jomelli V, Grancher D (2010) Asynchronous Little Ice Age glacial maximum extent in southeast Iceland. Geomorphology 114:253–260
Chenet M, Roussel E, Jomelli V, Grancher D, Cooley D (2011) A response to the commentary of M. Dąbski about the paper ‘Asynchronous Little Ice Age glacial maximum extent in southeast Iceland’. Geomorphology 128:103–104
Coulson SJ (2007) The terrestrial and freshwater invertebrate fauna of the high Arctic archipelago of Svalbard. Zootaxa 1448:41–58
Coulson SJ, Hodkinson ID, Strathdee AT, Bale JS, Block W, Worland MR, Webb NR (1993) Simulated climate change: the interaction between vegetation type and microhabitat temperatures at Ny-Ålesund, Svalbard. Polar Biol 13:67–70
Dąbski M (2010) A commentary to ‘Asynchronous Little Ice Age glacial maximum extent in southeast Iceland’ by Chenet et al. (Geomorphology 114 (2010) 253–260); a case of Fláajökull. Geomorphology 120:365–367
Dastych H (1985) West Spitsbergen Tardigrada. Acta Zool Cracov 28:169–214
De Smet WH, Van Rompu EA (1994) Rotifera and Tardigrada from some cryoconite holes on a Spitsbergen (Svalbard) glacier. Belg J Zool 124:27–37
De Smet WH, Van Rompu EA, Beyens L (1988) Contribution to the Rotifera and aquatic Tardigrada of Edgeoya (Svalbard). Fauna Norv Ser A 9:19–30
Eythórsson J (1951) Correspondence. Jőkla-mýs. J Glaciol 1:503
Fjellberg A (2007a) Icelandic Collembola. Revised checklist and general comments. Insect Syst Evol 64:45–60
Fjellberg A (2007b) The Collembola of Fennoscandia and Denmark. part II: entomobryomorpha and Symphypleona. Fauna Entomol Scand 42:1–264
Fjellberg A (2010) Cryophilic Isotomidae (Collembola) of the Northwestern Rocky mountains, U.S.A. Zootaxa 2513:27–49
Fjellberg A, Bernard EC (2009) Review of Agrenia Borner, 1906 with descriptions of four new species from North America (Collembola, Isotomidae). Zootaxa 2306:17–28
Gjelstrup P (2000) Soil mites and collembolans on Surtsey, Iceland, 32 years after the eruption. Surtsey Res 11:43–50
Graham DJ, Midgley NG (2000) Graphical representation of particle shape using triangular diagrams: an excel spreadsheet method. Earth Surf Proc Land 25:1473–1477
Hartzell PL, Shain DH (2009) Glacier ice worms. In: Shain DH (ed) Annelids in modern biology. Wiley, Hoboken, pp 301–313
Hartzell PL, Nghiem JV, Richio KJ, Shain DH (2005) Distribution and phylogeny of glacier ice worms (Mesenchytraeus solifugus and Mesenchytraeus solifugus rainierensis). Can J Zool 83:1206–1213
Hawes TC, Worland MR, Convey P, Bale JS (2007) Aerial dispersal of springtails on the Antarctic Peninsula: implications for local distribution and demography. Antarct Sci 19:3–10
Heusser CJ (1972) Polsters of the moss Drepanocladius berggrenii on Gilkey Glacier, Alaska. Bull Torrey Bot Club. 99:34–36
Hodkinson ID, Healey V, Coulson S (1994) Moisture relationships of the High Arctic collembolan Onychiurus arcticus. Physiol Entomol 19:109–114
Hodson AJ, Mumford PN, Kohler J, Wynn PM (2005) The High Arctic glacial ecosystem: new insights from nutrient budgets. Biogeochemistry 72:233–256
Hopkin SP (1997) Biology of the Springtails. Insecta: Collembola. Oxford University Press, Oxford
Jonsdottir IS (2005) Terrestrial ecosystems on Svalbard: heterogeneity, complexity and fragility from an Arctic Island perspective. P R Irish Acad B 105:155–165
Kopeszki H (2000) Auf der Suche nach rotten Gletscherflöhen. Funde hochalpiner Springschwänze (Collembola). Vorarlberger Naturschau 8:133–144
Krantz GW, Walter DE (2009) A manual of acarology, 3rd edn. Texas Tech University Press, Lubback
Makkonen M, Berg MP, van Hal JR, Callaghan TV, Press MC, Aerts R (2011) Traits explain the responses of a sub-arctic Collembola community to climate manipulation. Soil Biol Biochem 43:377–384
Perez FL (1991) Ecology and morphology of globular mosses of Grimmia longirostris in the Paramo de Piedras Blancas, Venezuelan Andes. Arct Antarct Alp Res 23:133–148
Porazinska DL, Fountain AG, Nylen TH, Tranter M, Virginia RA, Wall DH (2004) The biodiversity and biogeochemistry of cryoconite holes from McMurdo dry valley glaciers. Arct Antarct Alp Res 36:84–91
Porter PR, Evans AJ, Hodson AJ, Lowe AT, Crabtree MD (2008) Sediment-moss interactions on a temperate glacier: Falljökull, Iceland. Ann Glaciol 48:25–31
Pugh PJA, McInnes SJ (1998) The origin of Arctic terrestrial and freshwater tardigrades. Polar Biol 19:177–182
Säwström C, Mumford P, Marshall W, Hodson A, Laybourn-Parry J (2002) The microbial communities and primary productivity of cryoconite holes in an Arctic glacier Svalbard 79°N. Polar Biol 25:591–596
Scherrer D, Korner C (2010) Infra-red thermometry of alpine landscapes challenges climatic warming projections. Glob Change Biol 16:2602–2613
Shacklette HT (1966) Unattached moss polsters on Amchitka Island, Alaska. Bryologist 69:346–352
Southwood TRE, Henderson PA (2000) Ecological methods. Blackwell, Oxford
Walter DE, Procter HC (1999) Mites: ecology, evolution and behavior. CABI International, Oxford
Wharton RA, McKay CP, Simmons GM, Parker BC (1985) Cryoconite holes on glaciers. Bioscience 35:449–503
Acknowledgments
Fanny Dommanget for invaluable help in the laboratory and Arne Fjellberg Entomological Research for identification of the Collembola. Michael Stech and Hans Kruijer from the National Herbarium of the Netherlands, University of Leiden, for identifying the moss and checking synonyms. Guðrún Þórunn Gísladóttir assisted with the provision of Icelandic Meteorological Office data sets. NGM received funding from Nottingham Trent University to undertake fieldwork in Iceland and was assisted by Oz Godden, Karen Mather and Nikki Sandercock. Mike Pemulis is thanked for advice on the use of the dot product operation. We are also grateful to three anonymous reviewers and the Editor for their constructive comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Coulson, S.J., Midgley, N.G. The role of glacier mice in the invertebrate colonisation of glacial surfaces: the moss balls of the Falljökull, Iceland. Polar Biol 35, 1651–1658 (2012). https://doi.org/10.1007/s00300-012-1205-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-012-1205-4