
Abstract
To evaluate the movement rates of semi-domesticated reindeer (Rangifer tarandus tarandus) during the bare-ground season, we used successive GPS positions from 48 female reindeer. Data were collected during the summers of 2002 and 2003 in two Sámi reindeer herding districts in the Swedish mountains, Handölsdalen, and Sirges. The movement rates were analysed at five different time periods: over the whole season, and over the sub-seasons spring, and early summer, mid summer and early autumn. Variation in movements were analysed in relation to vegetation type, altitude, terrain ruggedness, temperature, wind speed, and proximity to hiking trails. We hypothesised that the foraging quality and different weather conditions is an important factor in determining movement rates. We found that reindeer movement rates were similar between study areas and were dependent on vegetation type and on weather conditions. Studying the circadian movements, in mid summer period when daytime oestrid activity are expected to be high, the reindeer stayed at higher altitudes where food quality was low, but moved to low altitudes at night where the food quality was higher. Therefore, we suggest that oestrid activity forces the reindeer to stay in low-quality vegetation types. Reindeer movements were linked to disturbance in areas of intermediate human activity. We found that in Handölsdalen, where hikers are abundant, the movement rates of reindeer decreased closer to the trails whereas in Sirges, where hikers are less abundant, the movement rates of reindeer increased closer to the trails.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
During the bare-ground season, most of the reindeer in Fennoscandia graze in the mountains. Although the majority of reindeer in Fennoscandia are domesticated and herded, they graze freely during the main part of the summer season, and their behaviour and habitat selection are comparable to wild reindeer. Reindeer play an important ecological and cultural role in Swedish mountain ecosystems (Bråthen et al. 2007; Swedish Government 2000). The Swedish Government has declared that the mountain range should be kept as a grazed landscape with its cultural and recreational assets. In this context, it is important to understand how reindeer use the landscape and how they are affected by different environmental factors.
Animal decision making can be considered a hierarchical process where selection occurs at different scales including: (1) the landscape at the regional level; (2) feeding sites or communities at the landscape level; and (3) feeding stations or plants at the patch level (Johnson 1980; Senft et al. 1987). Different factors also affect the animals’ decisions or selections at different scales and analyses of animal movements can aid our understanding of animal responses to the environment at these different scales (Ager et al. 2003). Understanding movement patterns is vital in studies of highly mobile species, such as reindeer.
The way in which large herbivores exploit their spatial environment is primarily driven by the distribution of resources (Danell et al. 2006). In summer, reindeer are dependent on fresh and nutritive forage, such as herbs and graminoids (Klein 1990; Skogland 1980). The main preferred vegetation types by reindeer in summer are therefore meadows, snow-beds, and heath lands.
Insect harassment is also a driving force for reindeer behaviour and habitat selection in summer (Espmark and Langvatn 1979; Mörschel and Klein 1997; White et al. 1981). It is well documented that herbivores harassed by insects prefer higher altitudes and wind-exposed sites, which provide relief from the insects (Hagemoen and Reimers 2002; Mooring et al. 2003; Powell et al. 2006; Skarin et al. 2004; White et al. 1975). However, these sites may not necessarily correspond to those preferred for foraging (Mårell et al. 2002; Powell et al. 2006; Skogland 1980). Previous research has also shown that insect activity can be related to weather parameters including temperature and wind speed (e.g. Downes et al. 1986; Gaare et al. 1975; White et al. 1975; Russell et al. 1993; Mörschel and Klein 1997; Hagemoen and Reimers 2002).
With the expansion of urban areas and increased levels of hunting and ecotourism, human–reindeer interactions have also increased. In the Swedish mountain chain, there are 8000 km of hiking trails, and these are unevenly distributed across the region. Hikers are abundant along the trails during certain time periods and they are more abundant in certain areas (Vuorio 2003). Many hikers expect to see reindeer during their treks, especially hikers from other countries (Wall-Reinius 2009). The eagerness to spot or interact with reindeer can create a potential conflict if the reindeer flee as hikers approach. This extra activity causes the reindeer to use energy, loose grazing time and potentially takes them from better habitats and pastures to poorer locations.
Previous studies of reindeer during the bare-ground season have considered their reactions and responses to harassment by oestrids (Hypoderma tarandi and Cephenemyia trompe), mosquitoes and species of Simuliidae (Colman et al. 2003; Mörschel and Klein 1997), and to disturbance from human activities (Colman et al. 2001; Dyer et al. 2001; Vistnes 2008). Although previous studies have focused on reindeer responses to insect harassment and human activities during the summer season, these two factors have rarely been considered together. However, there is some evidence to suggest that reindeer movement occurs in order to avoid insect harassment more than to avoid human activity (Noel et al. 1998; Skarin et al. 2004). Studying reindeer movement rates simultaneously with other environmental factors in areas with different hiking pressures may enable reindeer responses to hiking pressure to be distinguished.
In a previous study, Skarin et al. (2008) described resource utilisation function models (RUF; Marzluff et al. 2004), in which reindeer home range use was correlated to vegetation type, topography, distance to hiking trails and other human interactions. These models showed that the reindeer home range use was dependent on vegetation type, aspect, and landscape ruggedness. However, an analysis of reindeer behaviour and movement rate within the boundaries of the home range is necessary to fully understand habitat selection and the extent of home ranges in relation to different environmental factors.
The aim of this study was to evaluate reindeer movement rates at two different temporal extents (whole season and sub-seasons) using GPS-location fixes with 1-hour time intervals. We assessed the reindeer movement rates over the whole summer season as well as over different sub-seasons in relation to different environmental factors to identify general patterns of reindeer movement. The factors assessed were vegetation type, altitude, terrain ruggedness, temperature, wind speed, and disturbance from human activities (measured as the distance to the nearest hiking trail). We further describe the reindeer circadian movements within the sub-seasons, and we investigate the correlations between reindeer movement rate and their home range size in an effort to explain some of the results found.
Materials and methods
Study areas
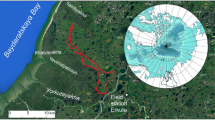
The study was conducted in the summer ranges of Handölsdalen (63°00′N, 12°30′E) and Sirges (67°00′, 17°10′E), which are both reindeer herding districts in the Scandinavian mountain range. This included three study areas, Handöl situated in Handölsdalen, and Vaisa and Sarek located in Sirges (Fig. 1). Vaisa and Sarek are separated by lakes and rivers, which limit but do not completely prevent migration of reindeer between the areas. The reindeer graze freely from May to October within the study areas, but are gathered for calf marking in July and bull slaughter in September. There were approximately 8,000 reindeer in Handöl, and 6,000 reindeer between Vaisa and Sarek.

Location of the three study areas: Handöl, Vaisa, and Sarek. Lakes and rivers separate Vaisa and Sarek, making extensive migration between these areas difficult for reindeer. Light colours in the topographic maps are higher altitudes. The weather stations Ritsem and Sylarna are also marked on the maps ©Lantmäteriet Gävle 2010. Permission I 2010/0055
The hiking season in the Swedish mountain range starts in July and ends in mid September (Vuorio 2003). The study areas were chosen dependent on the number of visitors. Handöl is the most popular of the three areas for backcountry hiking, and has a number of main trails connecting three mountain stations and four lodges. Generally, the highest frequency of visitors to the area occurs during the last week of July and first week of August (Mats Almlöf, Swedish Touring Club, Sylarna, personal communication, July 2006). Fewer tourists visit Vaisa and the area is mainly used for reindeer husbandry. In Sarek, the hiking trails follow the major valleys and are frequently used by backcountry hikers.
The area of the Handöl study site is 2,000 km2, Vaisa 1,000 km2, and Sarek 4,000 km2. The elevation in Handöl ranges from 600 m to the highest peaks at 1,796 and 1,761 m at Helags and Sylarna, respectively. In Vaisa, the elevations mostly range between 500 and 1,000 m, with the highest summit (Rautåive) at 1,516 m. Sarek has the broadest elevation range of the three sites from 500 to 2,015 m; this site also has long deep valleys and a more alpine character than the other two sites. All three areas have an alpine climate. The annual precipitation in Handöl is 700–1,300 mm, the mean daytime temperature in July is 10°C and the vegetation period (mean daytime temp >5°C) is 120–130 days (data averages for 1961–1990; Lantmäteriet 2002). In Vaisa, the annual precipitation is 900–1,500 mm and the mean temperature in July is 9°C. Sarek has an annual precipitation of 900–1,700 mm and the lowest mean temperature (7°C in July) of the three areas.
The vegetation period in both Vaisa and Sarek is 100–110 days. Since Handöl is further south than Vaisa and Sarek, the tree line occurs at higher elevations (900 m compared to 600 m in Vaisa and Sarek). The vegetation in Handöl is dominated by heaths, ranging from wet to extremely dry (Table 1). In Vaisa, the common vegetation types are dry heaths, meadows, grass heaths, bare rock, and sparsely vegetated areas. Sarek has approximately the same distribution of vegetation types as Vaisa, but with more glaciers and bare rock. In Handöl, 85% of the area consisted of high-quality vegetation, and in Vaisa and Sarek it covered approximately 60% of the area.
GPS data collection
The data was collected in two phases. From July 15th to September 15th 2002, 10 randomly sampled adult female reindeer at both Vaisa and Handöl were fitted with GPS collars (Posrec®, 650 g, manufactured by TVP Positioning AB). From May 10th to September 15th 2003, 10 new adult female reindeer in both areas and 10 additional reindeer in Sarek were fitted with GPS collars. Adult females were chosen as study animals since they represent the majority of the herds and they also tend to remain with the herd (Thomson 1972), and thus are likely to provide the most representative picture of how the reindeer herds use their ranges. The females were randomly sampled among those which were either rearing calves (2002) or pregnant (2003). In 2002 all reindeer had a calf, both at the calf marking stage and when the collars were removed in September. In 2003, most of the marked females had a calf (visually confirmed at calf marking), but when the collars were taken off in autumn 2003 only 60% of the reindeer still had a calf following them. The GPS equipment recorded and stored positions, and the data were downloaded after the collars were removed. In 2002, positions were collected every 2 h to increase battery life for the following year and in 2003 data were collected every hour. Less than 2% of the positions were missing due to malfunctioning equipment. In 2002, two collars (one in each area) failed to collect positions due to technical problems. In 2003, some migration occurred between Sarek and Vaisa: 12 reindeer spent most of their time in Sarek and 7 in Vaisa. In addition, one reindeer moved from Sarek to Vaisa on the 9th of August. One collar in Vaisa stopped working on the 18th of August 2003.
Data analyses
Linear mixed models of movement rates and associated habitat data were fitted for each area and period with individual reindeer as random effect using the Mixed Model procedure in the SAS 9.2 software (SAS Institute Inc© 2002–2008).
In the statistical analyses, the data from 2003 were sorted into five different time periods: the whole study season and four sub-seasons: spring, early summer, mid summer and early autumn. The sub-seasons were the same as those used by Skarin et al. (2008) for home range calculations; however, for this paper we changed the name late summer to early autumn since this is more appropriate. In 2002, we only collected data during half the season and every second hour. Therefore, this data set was only used to verify the analyses of the 2003 data set within the sub-seasons, mid summer and early autumn. The movement rates determined from the two-hourly dataset are expected to underestimate movement rate by 40% compared to 30% for the hourly data (Pepin et al. 2004). The time periods for the sub-seasons were set according to seasonal weather characteristics and with respect to handling events in reindeer husbandry (Table 2). The last day of the spring was defined as the average date when snow cover ended (Pershagen 1969); this sub-season has scarce forage due to snow cover. The last day of early summer was defined as the day before the reindeer herders started to gather the reindeer for calf marking. The early summer is the sub-season where vegetation growth begins, providing relatively good forage availability (Mårell and Edenius 2006). Mid summer started after the calf marking had finished and ended when the mean daily temperature during each study year fell below 6°C. This is the temperature at which insect activity is shown to decrease (Anderson et al. 1994; Mörschel and Klein 1997; Russell et al. 1993). The mid summer is the main growth season when the reindeer had good access to forage (Mårell and Edenius 2006). The end of the early autumn corresponded to the day before the reindeer were gathered for the bull slaughter in mid-September. Early autumn is a sub-season with good grazing conditions but with reduced insect harassment compared to mid summer.
The movement rates were calculated as the distance moved between every successive GPS location recorded divided by the time elapsed (Table 2 show the mean numbers of positions used). Movement rates were not calculated if there were one or more positions missing between the successive locations. The reindeer were actively herded during the calf marking periods, and therefore the positions from these periods were removed from the datasets before analyses.
Variables used to explain the movement rates were vegetation type; the altitudinal position of the reindeer; a terrain ruggedness index (a scale from 1 to 59, representing the difference in elevation between adjacent grid cells of a digital elevation grid as described by Riley et al. 1999, but transformed to a logarithmic scale); the slope of the movement (the altitudinal change between two positions divided by the distance moved); temperature; wind speed; and distance to the nearest hiking trail transformed to a logarithmic scale (since a trail far away most likely do not affect the reindeer). The digitised geographical data for vegetation type (Swedish CORINE Land Cover, 25 m raster resolution) and altitude (digital elevation model, 50 m raster resolution) were provided by The National Land Survey (http://www.lantmateriet.se). For the ‘heaths’ vegetation types, two different classification resolutions were available. We used the finer classification where available; otherwise the coarser classification was used. For the coarser resolution, all heath vegetation types (except grass heath) were combined into one category called ‘heaths’. This represented less than seven per cent of all the vegetation types available within each area (Table 1). The areas with the coarser classification are also in the outskirts of the study areas where the reindeer spent less time. Rare or similar vegetation types were further combined in all analyses to reduce the number of vegetation classes. For interpretation of our results, we also classified vegetation types into high- or low-quality vegetation type according to Klein (1990) and Skogland (1980), as shown in Table 1. Temperature (°C) and wind speed (m s−1) records from the weather stations at Sylarna and Ritsem (Fig. 1) for the Handölsdalen and Sirges study areas, respectively, were obtained at three-hourly intervals from the Swedish Meteorological and Hydrological Institute (http://www.smhi.se). Temperatures and wind speeds for every hour were calculated by linear interpolations. Possible correlations between the continuous variables were checked and significant correlations were found only between wind speed and temperature at Sylarna weather station (r = −0.135, P < 0.001). Therefore, a combination of temperature and wind speed was derived [temperature × (wind speed + 1)−1], but this variable had no significant effects in the analyses. In the mixed model analyses, the mean values of the continuous habitat attributes at the two successive GPS locations were used for each rate observation. The dominating vegetation type at each position was given the value 1 and other the value 0. For each rate observation, the mean values of the vegetation types were used as vegetation variable to which the models were fitted.
In addition, during the mid summer sub-season, temperature and wind speed were used to derive two variables reflecting oestrid, and mosquito and Simuliidae activities. Although, there are several studies that have investigated insect activity with respect to weather conditions, it is not possible to find sharp temperature thresholds or exact wind speed intervals that determine insect activity. For this reason, these variables were only used to discuss the possible correlations between reindeer movement rate and insect harassment. Temperature thresholds reported for oestrid (OES) fly activity are 7°C (Downes et al. 1986), 10°C (Mörschel 1999; Anderson et al. 1994), and 13°C (Colman 2000; White et al. 1975). The upper wind speed limit for OES activity has been reported as 8–9 m/s (Anderson et al. 1994), and 4 m/s as the average wind speed (Mörschel 1999). Mosquitoes and species of Simuliidae (MOS) have been reported to be active at temperatures between 6 and 18°C and wind speeds below 6 m/s (Russell et al. 1993), to decline in activity above 16°C and 7.5 m/s (Hagemoen and Reimers 2002), or to have the highest activity at 10.3°C and 3.8 m/s (Downes et al. 1986). Based on these studies, we assumed that OES activity occurred when wind speed was below 9 m/s and the temperature was above 11°C, and that MOS were mainly active at temperatures between 7 and 17°C and at wind speeds below 7 m/s, with discrepancy for Siimulidae that probably also is active above 17°C. Temperatures and wind speeds outside these ranges were assumed to have no insect activity (NOI). The OES and MOS variables are confounded to some extent.
To estimate the reindeer utilisation distributions (UD) or home ranges within each sub-season from the GPS-positions, we used fixed-kernel techniques (Worton 1989; Seaman and Powell 1996). Depending on time periods, between 236 and 1075 positions were available in the home-range estimations (Table 2). For the UD estimation, we used ArcView GIS 3.3 software (ESRI©1992–2002) with the Animal Movement extension (Hooge and Eichenlaub 2001). To decrease computational load, the smoothing parameter was calculated using the ad hoc method (Worton 1989). The spatial extent of the UD was defined as the 99% fixed-kernel home range boundary and was displayed in a 50 × 50 m raster.
Using the whole dataset, we initially tried to fit a single mixed model, with areas and sub-season as fixed effects and reindeer as random with first order interactions between fixed effects and continuous variables. However, in an effort to find a parsimonious model using Akakie information criterion (AIC), we found that removing non-significant interactions terms increased the AIC-value. Thus, the full model was probably the most appropriate among models with first-order interactions, but this model left us with interaction terms difficult to explain. We therefore divided the data by each sub-season, and fitted separate linear mixed models of the movement rates for the different habitat variables for each area and period with reindeer as random effect. We also fitted separate mixed models at the whole season scale within each area with reindeer as random effect. A least squares estimate for each vegetation type was obtained and expressed as deviation from the mean of estimates of those vegetation types that all the reindeer had visited in each study area. We used AIC to select the most parsimonious model. We compared AIC-values for the full model in each subset with models where continues variables with the highest P-value >0.20 were removed; if this model had a lower AIC-value we continued and excluded next variable until we found the most parsimonious model. The individual average movement rates within each sub-season were also correlated with the individual home range size using Pearson’s correlation. Further, we plotted circadian movement rate and mean altitude change within the three different weather categories of expected insect activity within the mid summer period. We also plotted the per cent of positions by the hour within high- and low-quality vegetation type during each sub-season.
Results
Movement rates over the whole summer
The mean movement rate for all animals was 344 ± 460 m/h, and there was no difference in the movement rate between the three areas (ANOVA, P = 0.711). The model selection ended up with using the full model for each area at this temporal interval. Furthermore, the reindeer in Handöl moved faster within five out of seven of the significant high-quality vegetation types (fresh heaths, grass heaths, meadow, birch forest and lakes (which included open surfaces and shores)), while they moved slower in the lower-quality vegetation types bare rock and sparsely vegetated areas (Table 3). In Vaisa, the reindeer moved slower within four out of six high-quality vegetation types (dry heath, fresh heaths, meadow, and mires), and in Sarek the reindeer moved slower through fresh heaths and willow thickets. In grass heaths though, the reindeer always moved faster. The reindeer also increased their movement rates with increasing temperature in all study areas. In Handöl and Sarek, the movement rates decreased with wind speed. The reindeer movement rates decreased with increasing altitude and in rugged terrain (Handöl and Sarek). In Sarek, the reindeer also increased their movement rate with increased distance to the hiking trails.
Movement rates within sub-seasons
Movement rates varied between the sub-seasons, with an increase from spring to mid summer and then a decrease in early autumn (Table 2). The model selection for the sub-season interval models resulted in excluding altitude in the model for Handöl in early summer and mid summer, and for Vaisa in spring and early autumn, otherwise the full model was used. There was a divergent pattern of movement rate within the different vegetation types (Tables 4, 5). Within spring the reindeer moved slower in the open high-quality vegetation types, while they moved faster in the forested high-quality vegetation types. In early summer, the reindeer moved slower within the majority of the high-quality vegetation types and faster within the low-quality vegetation, and in mid summer the pattern was reversed. Further, we found increased movement rates within grass heath in all areas in early summer and in Sarek in mid summer. In the early autumn, there was no obvious response in the movement rates within the different vegetation types. For further details about the movement rates within the vegetation type in each area and sub-season see Tables 4 and 5.
The reindeer response to the weather parameters was quite consistent. Movement rates increased with temperature in all sub-seasons. In all areas, the reindeer movement rates decreased with wind speed during mid summer, while an increase in reindeer movement rate with wind speed occurred in spring in Handöl, in Vaisa in early summer, and in all areas in early autumn. The reindeer movement rates were generally lower in rugged terrain and with increasing altitude. However, in Handöl in spring and in Sarek in early autumn, movement rates were higher when the reindeer moved over rugged terrain. Furthermore, the reindeer showed increased movement rates with increasing distance to hiking trails in most areas and sub-seasons; however, in Handöl during spring and in Vaisa and Sarek during mid summer, the reindeer increased their movement rate with decreasing distance to the hiking trails.
A strong positive correlation was found between the reindeer average home range sizes and average movement rate over the sub-seasons (r MR,HR = 0.722, P = 0.0001, N = 117). However, splitting the data into a single sub-season and one area, only a few correlations were found between home range size and movement rates (Table 2). In mid summer, the reindeer home ranges in Sarek were almost twice as large as those in the other areas, while their movement rates were similar at all sites. In Vaisa in all sub-seasons, but especially in spring and early summer, the reindeer had smaller home ranges compared to the other areas.
Circadian movements
During the days of expected insect harassment in mid summer, the movement rates had pronounced peaks at the beginning (around 0500 h) and at the end of the day (around 1900 h) as shown in Fig. 2a. These movements appeared to be associated with cyclical altitude positioning, i.e., the reindeer increased their movement rates as they moved up the hillsides in the mornings and down in the evenings (Fig. 2b).

a Patterns of average reindeer movement rates (m/h) in all three study areas in mid summer 2003, within the different weather categories, when the insects were predicted to be active. The assumed active insects were oestrid flies (OES) when temperature was ≥11°C and wind speed was ≤9 m/s, and mosquitoes and Simuliidae (MOS) when temperatures were ≥7°C and <17°C and wind speed was ≤7 m/s. The weather categories outside these thresholds were assumed to result in no insect harassment (NOI). Between 1400 and 1900 h, there were no NOI observations from Vaisa and Sarek. b The average altitude change (m) in the position of reindeer in all three study areas during mid summer 2003
The number of positions registered in vegetation types with high or low forage quality (Klein 1990; Skogland 1980; Table 1) differed over a 24-h cycle within the different sub-seasons (Fig. 3). Within the mid summer period, in all three areas, there was an increase in the number of positions registered in low-quality vegetation types in the day, while there were more positions found in vegetation types with high-quality forage at night. In spring and early summer and early autumn, there were no major differences in the use of different vegetation types during the circadian cycle.

The percentage of reindeer positions (by the hour of the day) on the high-quality vegetation types (meadows, grass heath, dry heath, fresh heath, wet heath, thickets and birch, and coniferous forests) in 2003, (according to Klein 1990). (Bare rock, glaciers and permanent snowfields, extremely dry heath, and sparsely vegetated areas were classified as sparsely vegetated habitat types)
Discussion
We found evidence for association of forage quality and movement rate in the response of movement rate on the different vegetation types. Within early summer, the reindeer had slower movement rates within the high-quality vegetation types (Klein 1990; Skogland 1980), dry heaths, wet heaths, heaths, meadows, willow thickets and birch forest and higher movement rates through low-quality vegetation types, such as bare rock and bedrock outcrops and sparsely vegetated areas. This implies that the reindeer stopped for foraging at the high-quality sites and passed over the lower-quality sites. Mårell et al. (2002) also found that reindeer preferentially browsed within heath and willow vegetation types especially at the beginning of the summer season. The unexpected high movement rates in grass heath; although this is normally a preferred vegetation type (Skarin 2007), may be the result of reindeer moving across them quickly to seek insect relief. According to the reindeer herders in Vaisa, the reindeer in this area preferentially graze on grass heath close to snow patches, where they are close to insect relief habitats (Nils-Johan Utsi, reindeer herder in Vaisa, personal communication February 2003). The faster movements over high-quality vegetation types in mid summer may also be explained by the reindeer seeking insect relief and will be discussed later. In rugged terrain, slower movement may depend on higher forage quality (Mårell and Edenius 2006) or it could simply be due to the ruggedness of the terrain making it difficult to move across the area. The slower movements in the high and dense vegetation types such as, willow thickets, fresh heath and wet heath may also be caused by the vegetation restricting the reindeer movements.
The distribution of the reindeer within each study area also seemed to be a reflection of forage availability. Interestingly, we found that the reindeer movement rate was the same irrespective of the size of the study area and the size of the home range, but differed between sub-seasons. This indicates that the movement are mostly associated with the forage quality and the characteristics of the different areas. The large home ranges in Sarek in mid summer was probably caused by the more alpine character of the area, with long deep valleys; which forced reindeer to walk along the valleys to reach new grazing areas, thus enlarging their home ranges. The smaller home ranges in Vaisa might be a reflection of its flat landscape compared to the other areas. Thus, the reindeer did not need to expand their home ranges to the same extent as at other sites to reach new areas. According to the herders in the Sirges reindeer-herding district, the reindeer from Vaisa always weigh more and are in good physical shape, while the reindeer in Sarek have lower slaughter weights and are not as healthy. Our results suggest that the forage quality at Vaisa is exceptionally good, since the reindeer have such small home ranges. The reindeer manage to gain more weight even though they forage over a smaller area than the reindeer in the other areas.
The gradual increase in movement rate and home range size throughout the bare ground season may have several explanations. One possible explanation for the continuous change is the increase in body size and moving capacity of the calves. The mothers stay in close contact with the calves during the first month, when the calves only move short distances. At one-month old, the calves become gradually more independent, after which their positions within the grazing herd are as random as that of any other individual (Espmark 1971). However, our results show that the behaviour is due to a combination of factors.
Irrespective of the temporal period investigated, weather conditions were important in determining reindeer movement rates. Insect avoidance at higher altitudes and at wind-exposed sites might be an explanation for the pattern we report (Downes et al. 1986; Gaare et al. 1975; Mooring et al. 2003; Powell et al. 2006). Insect harassment, especially from oestrids, has been found to be most severe in the middle of the summer (Anderson et al. 1994; Hagemoen and Reimers 2002). Therefore, the reindeer response to changes in weather conditions and correlated changes in insect activity probably interact, especially during the mid summer sub-season. As there was a strong response to the weather parameters during this sub-season, we tried to relate the variables derived from temperature and wind speed, aiming to reflect insect activity, with reindeer movement rates. However, as we did not have direct observations of insect activity these results are not shown. Nevertheless, the models showed that the reindeer had higher movement rates during periods of expected OES activity and lower movement rates during periods of expected MOS activity, as seen in Fig. 2a. This supports the view that insect activity causes the movement patterns found. The increased movement during the assumed OES activity periods appears to be specific to Rangifer, since other ungulates are known to decrease their activity during parasite attack (Espmark and Langvatn 1979; Mooring and Hart 1992; Mörschel and Klein 1997). The decrease of movement rate in the mid summer sub-season with increasing altitude and within the low-quality vegetation types, bare rock and sparsely vegetated area was probably also a response to finding insect relief at higher altitudes. Decreased movement rates during expected mosquito harassment have been reported previously (Downes et al. 1986; Mörschel and Klein 1997) and this is common herder knowledge that is used to facilitate herding since the reindeer form large herds at this time. It has also been suggested that an individual is less likely to be harassed by insects if it stands still in a larger herd, because of the increased probability of neighbours being harassed instead (Helle and Aspi 1983; Mooring and Hart 1992).
In mid summer, reindeer appeared to use sparsely vegetated habitat types in the day and vegetation of high quality at night (Fig. 3), which may be linked to the assumed insect activity. The high movement rates in the high-quality vegetation types during this sub-season may also be linked to the insect harassment. This implies, that not only does the reindeer lose grazing time due to insect harassment (Colman et al. 2003), they are also forced into areas with lower grazing quality. When forage quality decreases, the search time for forage also increases, making feeding more difficult (Illius 2006). The reindeer’s own impact on the vegetation by trampling may also increase, which in turn might lead to increased wear of the vegetation, especially in vegetation types that are sensitive to trampling (Moen and Lagerström 2008; Bråthen et al. 2007). Outside the insect season, the reindeer habitat selection within the home range seemed to be more dependent on forage quality, since they stayed in high-quality vegetation types during both day and night.
We expected a response to human disturbance along the hiking trails, especially in mid summer, which coincides with the hiking season. The response to expected human activity differed among the areas, however. In Handöl during mid summer, where hikers are frequent, the reindeer appeared to have habituated to human activity; it seemed more important to avoid insect harassment and to reach high-quality grazing grounds than to totally avoid hiking trails. This contradicts with our findings in Sarek, where the reindeer showed increased movement rates with decreased distance to the hiking trails. In Sarek there are few hikers (Wall-Reinius 2009) compared to Handöl (Vuorio 2003). As a result, the reindeer in Sarek may not have become habituated to hikers, to the same degree as in Handöl. On the other hand, the reindeer in Handöl had less opportunities to avoid the hiking trails; the mean distance (for each day and reindeer) to hiking trails in Handöl never exceeded 4.2 km while in Sarek the reindeer mean distance to hiking trails was up to 18 km. Vistnes (2008) suggests that reindeer is affected by human activities within a zone of 4 km from the activity, but in Handöl this was hardly possible. Even though, our result supports the view that if animals perceive an activity as spatially predictable and non-threatening, they may habituate to that activity (Colman et al. 2001; Whittaker and Knight 1998), and the question remains if reindeer may perceive human activities as predictable. The reindeer response to hiking trails in Handöl in spring was not expected, since human activity would have been low in the area but still they avoided hiking trails. However, one explanation may be the presence of snowmobiles and skiers in the early spring sub-season. Further, the reindeer use of areas close to hiking trails probably coincided with the fact that these often contain preferred vegetation types that are snow-free early (Skarin 2007). The slower movement rates of reindeer closer to the hiking trails in spring and early summer probably coincided with proximity to attractive vegetation types.
In conclusion, reindeer movement rates were dependent on forage quality, topography and weather parameters. During periods of expected insect harassment, the reindeer seemed more occupied by finding insect relief than finding good forage. In areas with low hiking activity, the reindeer reacted on both expected insect harassment and human activity. Thus, in management and planning of new hiking areas or hiking trails it is important to take in consideration reindeer and reindeer husbandry, especially during the high season of insect harassment. However, this might be difficult since this often is the high season for hiking and trails often are located in productive parts of the landscape. Therefore, we suggest a careful spatial planning of mountain areas, where vegetation types or areas with high-quality forage attractive to reindeer need to be prioritised.
References
Ager AA, Johnson BK, Kern JW, Kie JG (2003) Daily and seasonal movements and habitat use by female rocky mountain elk and mule deer. J Mammal 84:1076–1088
Anderson JR, Nilssen AC, Folstad I (1994) Mating behavior and thermoregulation of the reindeer warble fly, Hypoderma tarandi L. (Diptera:Oestridae). J Insect Behav 7:679–706
Bråthen KA, Ims RA, Yoccoz NG, Fauchald P, Tveraa T, Hausner VH (2007) Induced shift in ecosystem productivity? Extensive scale effects of abundant large herbivores. Ecosystem 10:773–789
Colman JE (2000) Behaviour patterns of wild reindeer in relation to sheep and parasitic flies. Dissertation, University of Oslo
Colman JE, Jacobsen BW, Reimers E (2001) Summer response distances of Svalbard reindeer Rangifer tarandus platyrhynchus to provocation by humans on foot. Wildl Biol 7:275–283
Colman JE, Pedersen C, Hjermann DØ, Holand Ø, Moe SR, Reimers E (2003) Do wild reindeer exhibit grazing compensation during insect harassment? J Wildl Manag 67:11–19
Danell K, Bergström R, Duncan P, Pastor J (2006) Large herbivore ecology, ecosystem, dynamics and conservation. Cambridge University Press, Cambridge
Downes CM, Theberge JB, Smith SM (1986) The influence of insects on the distribution, microhabitat choice, and behaviour of the Burwash caribou herd. Can J Zool 64:622–629
Dyer SJ, O’Neill JP, Wasel SM, Boutin S (2001) Avoidance of industrial development by woodland caribou. J Wildl Manag 65:531–542
Espmark Y (1971) Mother-Young relationship and ontogeny of behaviour in reindeer (Rangifer tarandus L.). Z Tierpsychol 29:42–81
Espmark Y, Langvatn R (1979) Lying down as a means of reducing fly harassment in red deer (Cervus elaphus). Behav Ecol Soc 5:51–54
Gaare E, Thomson BR, Kjos-Hansen O (1975) Reindeer activity on Hardangervidda. In: Wielgolaski FE (ed) Fennoscandian tundra ecosystems, Part 2. Animals and system analysis, vol 17. Springer, Berlin, pp 206–215, 337
Hagemoen RM, Reimers E (2002) Reindeer summer activity pattern in relation to weather and insect harassment. J Anim Ecol 71:883–892
Helle T, Aspi J (1983) Does herd formation reduce insect harassment among reindeer? A field experiment with animal traps. Acta Zool Fenn 175:129–131
Hooge PN, Eichenlaub B (2001) Animal movement extension 2.0 to ArcView. Alaska biological science center, U.S. Geological Survey, Anchorage, Alaska, USA. Available at: http://www.absc.usgs.gov/glba/gistools
Illius AW (2006) Linking functional responses and foraging behaviour to population dynamics. In: Danell K, Bergström R, Duncan P, Pastor J (eds) Large herbivore ecology, ecosystem, dynamics and conservation. Cambridge University Press, Cambridge, pp 77–96
Johnson DH (1980) The comparison of usage and availability measurements for evaluating resource preference. Ecology 6:65–71
Klein DR (1990) Variation in quality of caribou and reindeer forage plants associated with season, plant part, and phenology. Rangifer Spec Issue 3:123–130
Lantmäteriet (2002) National atlas of Sweden. Vol. 2004. Metria Kartcentrum, Vällingby
Mårell A, Edenius L (2006) Spatial heterogeneity and hierarchical feeding habitat selection by reindeer. Arct Antarct Alp Res 38:413–420
Mårell A, Ball JP, Hofgaard A (2002) Foraging and movement paths of female reindeer: Insights from fractal analysis, correlated random walks, and Levy flights. Can J Zool 80:854–865
Marzluff JM, Millspaugh JJ, Hurvitz P, Handcock MS (2004) Relating resources to a probabilistic measure of space use: forest fragments and Steller’s Jays. Ecology 85:1411–1427
Moen J, Lagerström A (2008) High species turnover and decreasing plant species richness on mountain summits in Sweden: Reindeer grazing overrides climate change? Arct Antarct Alp Res 40:382–395
Mooring MS, Hart BL (1992) Animal grouping for protection from parasites–Selfish herd and encounter-dilution effects. Behaviour 123:173–193
Mooring MS, Fitzpatrick TA, Fraser IC, Benjamin JE, Reisig DD, Nishihira TT (2003) Insect-defense behavior by desert bighorn sheep. Southwest Nat 48:635–643
Mörschel FM (1999) Use of climatic data to model the presence of oestrid flies in caribou herds. J Wildl Manag 63:588–593
Mörschel FM, Klein DR (1997) Effects of weather and parasitic insects on behaviour and group dynamics of caribou of the Delta Herd, Alaska. Can J Zool 75:1659–1670
Noel LE, Pollard RH, Ballard WB, Cronin MA (1998) Activity and use of active gravel pads and tundra by Caribou, Rangifer tarandus granti, within the Prudhoe Bay oil field, Alaska. Can Field Nat 112:400–409
Pepin D, Adrados C, Mann C, Janeau G (2004) Assessing real daily distance travelled by ungulates using differential GPS locations. J Mammal 85:774–780
Pershagen H (1969) Snötäcket i Sverige 1931–60 [Snow-cover in Sweden 1931–60]. Rep No 5, Swedish Meteorological and Hydrological Institute, Stockholm
Powell DM, Danze DE, Gwinn MA (2006) Predictors of biting fly harassment and its impact on habitat use by feral horses (Equus caballus) on a barrier island. J Ethol 24:147–153
Riley SJ, DeGloria SD, Elliot R (1999) A terrain ruggedness index that quantifies topographic heterogeneity. Intermt J Sci 5:23–27
Russell DE, Martell AM, Nixon WAC (1993) Range ecology of the Porcupine caribou herd in Canada. Rangifer Spec Issue 8:1–167
Seaman DE, Powell RA (1996) An evaluation of the accuracy of kernel density estimators for home range analysis. Ecol 77:2075–2085
Senft RL, Coughenour MB, Bailey DW, Rittenhouse LR, Sala OE, Swift DM (1987) Large herbivore foraging and ecological hierarchies. Bioscience 37:789–799
Skarin A (2007) Habitat use by semi-domesticated reindeer, estimated with pellet-group counts. Rangifer 27:121–132
Skarin A, Danell Ö, Bergström R, Moen J (2004) Insect avoidance may override human disturbances in reindeer habitat selection. Rangifer 24:95–103
Skarin A, Danell Ö, Bergström R, Moen J (2008) Summer habitat preferences of GPS-collared reindeer Rangifer tarandus tarandus. Wildl Biol 14:1–15
Skogland T (1980) Comparative summer feeding strategies of Arctic and Alpine Rangifer. J Anim Ecol 49:81–98
Swedish Government (2000) The Swedish Environmental Objectives—Interim Targets and Action Strategies. In Prop. 2000/01:130. The Swedish Government, Stockholm
Thomson BR (1972) Leadership in wild reindeer in Norway. In: Luick JR, Lent PC, Klein DR, White RG (eds) First international Reindeer and Caribou symposium, 1975. Fairbanks, University of Alaska, pp 462–472
Vistnes II (2008) Impacts of human development and activity on reindeer and caribou habitat use. Dissertation, Norweigan University of Life Sciences
Vuorio T (2003) Information on recreation and tourism in spatial planning in the Swedish mountains—methods and need for knowledge. Licentiate dissertation, Blekinge Institute of Technology
Wall-Reinius S (2009) Protected attractions—tourism and wilderness in the Swedish mountain region. Dissertation, University of Stockholm
White RG, Thomson BR, Skogland T, Person SJ, Russell DE, Holleman DF, Luick JR (1975) Ecology of caribou at Prudhoe Bay, Alaska. In: Brown J (ed) Ecological investigations of the tundra biome in the Prudhoe Bay region, Alaska. Biological papers of the University of Alaska, Special Report Number 2, October, 1975. University of Alaska, Fairbanks, pp 151–187
White RG, Bunnell FL, Gaare E, Skogland T, Hubert B (1981) Ungulates on arctic ranges. In: Bliss LC, Heal OW, Moore JJ (eds) Tundra ecosystems a comparative analysis, The Int Biol Progr. Cambridge University Press, Cambridge, pp 397–483
Whittaker D, Knight RL (1998) Understanding wildlife responses to humans. Wildl Soc Bull 26:312–317
Worton BJ (1989) Kernel methods for estimating the utilization distribution in home-range studies. Ecol 70:164–168
Acknowledgments
The Swedish Foundation for Strategic Environmental Research (MISTRA), through the Mountain Mistra Programme, and the Sami Fund (Samefonden) are acknowledged for their financial support of this study. The study was only made possible through the cooperation of the Sami reindeer herders in the Handölsdalen and Sirges reindeer herding districts. Thanks also to Jonathan Colman and two other anonymous referees for improving the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Skarin, A., Danell, Ö., Bergström, R. et al. Reindeer movement patterns in alpine summer ranges. Polar Biol 33, 1263–1275 (2010). https://doi.org/10.1007/s00300-010-0815-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-010-0815-y