
Abstract
The distribution and diversity of isopods (Crustacea, Isopoda; families Desmosomatidae Sars, 1897 and Nannoniscidae Hansen, 1916) was examined in Icelandic waters where a diversity of water masses (temperature range −0.9 to 12°C) occurs and the topography is characterized by the large and shallow Greenland–Iceland–Faeroe (GIF) Ridge extending across the North Atlantic in an east-west direction. Both families were species rich in the area, in total occurring with 34 species in 20 genera. Most of the species were restricted either to the north (10) or to the south (14) of the GIF Ridge, occurring either in cold (−0.8 to 2.8°C) or warm (>2°C) water masses. Five species were found on both sides of the Ridge, occurring at a wide range of temperatures (−0.9 to >4°C), while another five species extend across the GIF Ridge. Most species occurred in two and more water masses and the species north of the Ridge occurred usually within narrower temperature range than the species living south of the Ridge. The water masses in association with the Ridge seem to limit distribution of most species. Despite similar number of species occurring in the areas north and south of the Ridge, the areas differ considerably in diversity pattern with depth.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
A variety of factors shape species distributions, both historic and ecological, and these are often difficult to separate. Temperature has for a long time been recognized as one of the most important factors shaping distributions, not at least in the polar and sub-polar marine regions, where subzero temperatures dominate in large areas. The shallow and deep waters of the Arctic Ocean and the Nordic Seas (Greenland, Iceland and Norwegian Seas) hold a unique diversity of water masses, with temperatures of different ranges from −0.9°C to approximately 12°C (Hansen and Østerhus 2000). The northernmost part of the North Atlantic is further characterized by having a Ridge of shallow depth, the Greenland–Iceland–Faeroe (GIF) Ridge, extending across the Atlantic in an east-west direction and separating the Nordic Seas and the Arctic Ocean from the North Atlantic proper. Due to the deepest sill depth at about 860 m depth between the Faeroe Islands and Scotland, the deep water basins (>4,000 m) to the north and to the south of the GIF Ridge are largely separated from each other (Hansen and Østerhus 2000). The average sill depth in the region of the study area is 620 m in the Denmark Strait and 480 m between Iceland and the Faeroe Islands.
Extensive information on distribution on benthic invertebrates in the GIF Ridge area indicates that many species have their distributional limits in the vicinity of the Ridge. Furthermore, there are considerable differences in the species composition between the cold areas north of the Ridge and the warm areas south of the Ridge (e.g. Hansen 1916; Stephensen 1937, 1943; Svavarsson et al. 1993; Negoescu and Svavarsson 1997; Gudmundsson 1998; Weisshappel 2000, 2001; Schuchert 2000; Stransky and Svavarsson 2006; Dijkstra et al. 2009). It is not clear what shapes the species distribution at the Ridge, if it is the topography of the area, i.e. the physical presence of the Ridge itself, or the environmental parameters, i.e. temperature, sediments, food availability, etc., at the Ridge and the overlying waters.
The Desmosomatidae Sars, 1897 (Isopoda, Asellota) are among the most diverse families of deep-sea isopods. The members of the Desmosomatidae and the Nannoniscidae Hansen, 1916 are benthic species dwelling in the uppermost centimetre of the bottom (Hult 1941; Hessler and Strömberg 1989). Like all asellote isopods these are brooders, so their dispersal abilities are suspected to be poor compared to other macrofaunal taxa with pelagic larvae. Copulatory males may, however, enter the water column in search for females. Actually, one has to think about the speed of bottom currents as both asellote families treated here are epifaunal. Valsangkar (2004) mentions a transport of sediment in the deep sea may reach a distance of 15 km within 6 days. The families are closely related (see Hessler 1970a; Siebenaller and Hessler 1977, 1981; Svavarsson 1984; Wilson 2008a). Combined, the families are represented by about 207 described species in the world oceans, and they are frequently collected from the deep sea (Wolff 1962; Poore et al. 1994; Brandt et al. 2004, 2007; Brix 2006a, b, 2007, Kaiser and Brix 2005, 2007; Schotte et al. 2009; Wilson 2008b). They are well represented in the North Atlantic Ocean and the Nordic Seas, being commonly found in shallow waters at high latitudes and at bathyal and abyssal depths in the Nordic Seas and the Arctic Ocean (Hansen 1916; Wolff 1962; Hessler 1970a; Svavarsson 1988). The Desmosomatidae can be regarded as a true deep-sea family, considering that upper limit of the bathymetrical distributions of >50% of known species in below 1,000 m (Svavarsson et al. 1993). Polar emergence has been postulated for the family (Hessler 1970b). Isopods in general have been studied for more than a century in the Nordic Seas, the Arctic Ocean and the northernmost part of the North Atlantic (Sars 1899; Hansen 1916; Gurjanova 1946; Menzies and Mohr 1962; Just 1970, 1980; Paul and George 1975; Svavarsson 1982, 1984, 1988, 1997; Svavarsson et al. 1990, 1993), the most extensive information of the isopods at the GIF Ridge being gathered by during the Danish Ingolf Expedition in 1895 and 1896 (Hansen 1916).
The BIOICE project (Benthic invertebrates in Icelandic waters) was designed to investigate the faunal composition and species distribution in the northernmost part of the North Atlantic and the Nordic Seas. Here, we evaluate the distribution, faunal composition and diversity of the Desmosomatidae and Nannoniscidae in Icelandic waters. We evaluate the following:
Do the isopods occur at wide or narrow ranges in the temperature and are they restricted to any specific water mass or masses (water bodies with certain temperature and salinity characteristics) found in the area?
Does the topography of the GIF Ridge shape the distribution of asellote isopods or do other abiotic factors, like temperature, salinity, sediment or depth, limit the species distribution in the area?
Materials and methods
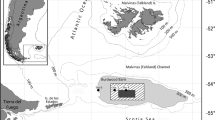
Study area (Fig. 1)
The study area comprises the northernmost part of the North Atlantic and the south-western part of the Nordic Seas (i.e. the Iceland, Greenland and Norwegian Seas). This area is characterized by a prominent topographic feature, the GIF Ridge. The Ridge is generally shallow, with a maximum sill depth of 620 m in the Denmark Strait between Greenland and Iceland, a maximum sill depth of 480 m between Iceland and the Faeroes and a maximum sill depth of 840 m between the Faeroes and Scotland (Hansen and Østerhus 2000). Generally, the Ridge is considerably shallower than the sill depths and most of the Denmark Strait is shallower than 300 m.

BIOICE stations used in this study
The hydrography of the area is complicated and diverse, with several primary water masses often overlying each other (Stefánsson 1962; Hansen and Østerhus 2000; Malmberg and Valdimarsson 2003). All water masses occur also at the bottom. For the BIOICE stations observed during this study the intersections of the water masses with the benthic boundary layer are known and shown in Fig. 2. The water masses dominating the study area are Modified North Atlantic Waters (MNAW; 7.0–8.5°C, salinity 35.10–35.30), Labrador Sea Water (LSW; 3–4°C, salinity 34.90–34.95), Iceland Sea Overflow Water (ISOW; 2–3°C, salinity 34.85–35.00); Modified East Icelandic Water (MEIW; 1–3°C, salinity 34.70–34.90), Norwegian Sea Deep Water (NSDW; <−0.5°C, salinity <−34.40), Norwegian Sea Arctic Intermediate Water (NSAIW; −0.5 to 0.5°C, salinity 34.85–34.90), Arctic/Polar Water (A/PW; 0–2°C, salinity 34.30–34.90) and Coastal Water (CW; 5–12°C, salinity <34.50) all differing in salinity (total range 34.30–35.45) and temperature (total range −0.9 to 12°C). Individual stations were assigned to water masses using Stefánsson (1962), Hansen and Østerhus (2000) and Malmberg and Valdimarsson (2003).

Intersections of the water masses with the benthic boundary layer for the RP sledge stations: Modified North Atlantic Waters (MNAW), Labrador Sea Water (LSW), Iceland Sea Overflow Water (ISOW); Modified East Icelandic Water (MEIW), Norwegian Sea Deep Water (NSDW), Norwegian Sea Arctic Intermediate Water (NSAIW), Arctic/Polar Water (A/PW) and Coastal Water (CW)

Distribution of species in the sampling area. Northern species. a Austroniscus groenlandicus Hansen, 1916; b A. norbi Svavarsson, 1988; c Cryodesma agnari Svavarsson, 1988; d Desmosoma strombergi Svavarsson, 1988; e Exiliniscus hanseni Just, 1970; f Hebefustis sp. A; g Nannoniscoides angulatus Hansen 1916; h Nannoniscus reticulatus Hansen 1916

Distribution of species in the sampling area. a, b northern species. a Nannoniscus cf. aequiremis Hansen, 1916; b Torwolia creper Hessler, 1970. c–f northern species with tendency to cross the Ridge southwards. c Desmosoma latipes (Hansen, 1916); d Eugerdella cf. armata (Sars, 1864); e Eugerda tenuimana (Sars, 1864); f Eugerdella hessleri (Just, 1979). g, h, northern and southern species. g Eugerda cf. globiceps Meinert, 1890; h Eugerda cf. reticulata (Gurjanova, 1946)

Distribution of species in the sampling area. a–c Northern and southern species. a Nannoniscus profundus Svavarsson, 1988; b Oecidiobranchus nanseni Just, 1980; c Pseudomesus brevicornis (Hansen, 1916). d Southern species with tendency to cross the Ridge northwards: Prochelator lateralis (Sars, 1899). e–h Southern species. e Chelator insignis (Hansen, 1916); f Desmosoma lineare Sars, 1864; g cf. Desmosoma tetarta (Hessler, 1970); h Echinopleura aculeata Sars, 1897

Distribution of species in the sampling area. a–h southern species. a Eugerdella polita (Hansen, 1916); b Exiliniscus clipeatus Siebenaller and Hessler, 1981; c Hebefustis cf. vafer Siebenaller and Hessler, 1977; d Mirabilicoxa acuminata Hessler, 1970; e Mirabilicoxa gracilipes (Hansen, 1916); f Nannoniscus cf. oblongus Sars, 1870; g Nannonisconus sp. A; h Rapaniscus crassipes (Hansen, 1916)

Distribution of species in the sampling area. a–b Southern species. a Thaumastosoma platycarpus Hessler, 1970; b Whoia angusta (Sars, 1899)
The area south of the Ridge is characterised by warm waters (e.g. MNAW; LSW), while the area north of the Ridge is characterised by cold waters (e.g. NSDW). Most of the shallower parts of the Ridge are influenced by warm waters (e.g. MNAW, MEIW, CW), while the deepest channels and some Ridge depressions hold cold waters, either constantly or intermittently (e.g. NSAIW). Cold waters from the north partly flow along the East Greenland current into the North Atlantic as surface waters (A/PW) or as overflow across the Denmark Strait and the Iceland–Scotland Ridge (e.g. NSAIW). The Iceland–Faeroe Ridge overflow may occur in five branches along depressions between Iceland and the Faeroes. In contrast to the eastern branches, the western branches of this overflow are presumably continuous, but the overflow water is highly variable, both spatially and temporally. A major amount of water flows through the Faeroe–Shetland channel. Accordingly, there are bands of warm and cold waters across the Ridge and in some places these bands vary in character. A detailed overview is given in Hansen and Østerhus (2000).
At each station, except stations taken on cruise with the RV Magnus Heinason, temperature and salinity were measured. Based on these data, stations were assigned to the water masses (Fig. 2).
Sampling and analysing
Samples were taken during the BIOICE project from 1991 to 2004. In total, 1,411 samples were collected with various instruments during the BIOICE cruises on the research vessels Bjarni Sæmundsson, Håkon Mosby and Magnus Heinason. The present study is based on 223 samples taken with a modified Rothlisberg–Pearcy epibenthic sledge (RP sled; Rothlisberg and Pearcy 1977; Brattegard and Fosså 1991) with 0.5 mm mesh in the cod end (Fig. 1; complete species by sample matrix is available as electronic supplement; ES1).
The samples were decanted through a series of sieves, the smallest mesh size being 0.5 mm. The specimens were then preserved in 10% buffered formalin and later transferred to 80% ethanol. Samples were sorted at the BIOICE facilities in Sandgerði where all specimens were sorted to family. The authors determined a total of 13.425 specimens of the Desmosomatidae and Nannoniscidae from 223 samples on species level. For species, which appeared to be similar, but not identical with the original description (holotype) of the species, a “cf.” was put in front of the species name. The term diversity refers here to ‘local diversity’ or ‘alpha diversity’.
Shipek grab samples were taken at some of the same station as the RP sled samples. These were analysed for sediment composition. The sediments were defined by the percentage of grain size (percentage of sand, muddy sand, sandy mud and gravel).
Maps were generated using ArcGIS 9.0. The Primer package (Clarke and Warwick 2001) version 6 was used for non-metric multidimensional scaling (nMDS) and cluster analysis to visualize faunal similarities. Abundance data were transformed by fourth root. Bray–Curtis similarity was used for the similarity matrix. Only species occurring at more than five stations were included in the data matrix. Four variables (Table 1) were analysed using BIOENV (rank correlation method: Spearman, resemblance measure: D1 Euclidean distance) to measure which abiotic factor has the strongest influence (Clarke and Ainsworth 1993). Differences between groups (defined a priori), using permutation/randomisation methods on resemblance matrix was tested with a one way designed “analysis of similarities” (ANOSIM, Clarke and Warwick 2001). To compare differences between groups, data were analyzed using the chi-square test with the statistical software package Sigma Stat version 3.5.
Results
In total, 13.425 specimens of desmosomatid and nannoniscid isopods were collected on the 223 analysed stations. These have been assigned to 20 genera and 34 species (Table 2), 12 desmosomatid (Chelator, Cryodesma, Desmosoma, Echinopleura, Eugerda, Eugerdella, Mirabilicoxa, Oecidiobrancus, Prochelator, Thaumastosoma, Torwolia and Whoia) and 8 nannoniscid (Austroniscus, Exiliniscus, Hebefustes, Nannoniscoides, Nannonisconus, Nannoniscus, Pseudomesus and Rapaniscus) genera.
The most common species was Chelator insignis comprising 6,100 specimens collected at 94 stations (i.e. 43% of the stations; Table 2). Eugerda reticulata was the second most abundant species occurring at 63 stations (28.3% of the stations) with the total number of 1,943 specimens. The third and the fourth most frequently occurring species were Eugerda cf. globiceps and Oecidiobranchus nanseni, occurring on 56 stations, respectively. Nine of the 34 species occurred at fewer than five stations, and three species occurred at a single station only. Most species were well represented and only five species had fewer than ten specimens each.
Nineteen species had a fairly wide depth range (>1,000 m, Table 2; Fig. 8). There was a tendency for a wide bathymetric range in species with a wide geographic range, i.e. seen in species found at several stations. Twelve of the 19 species with a depth range of >1,000 m were found at ten or more stations (Fig. 8; Table 2).

Bathymetric range of individual species. Filled bars southern species, open bars northern and southern circumicelandic species, gray bars northern species
Only a few ubiquitous species (occurring at ≥7 stations) were restricted to shallow water, i.e. Echinopleura aculeata, Eugerda tenuimana and Eugerdella cf. armata. In all, 13 species occurred in relatively shallow waters (≤300 m) towards considerable depths (>1,000 m). Eight species occurred only at depths greater than 840 m, deeper than the deepest sill depth of the Ridge.
Distributional patterns
In all, ten species were only found north of the Ridge. Some of the species, e.g. Austroniscus groenlandicus (Fig. 3a) and Austroniscus norbi (Fig. 3b) occurred only in fairly shallow waters (347–552 and 539–831 m, respectively) north of Iceland, both in water masses typical of the intermediate and deeper waters (NSAIW and NSDW), but A. groenlandicus also occurred in the A/PW. Two species, i.e. Hebefustis sp. A (Fig. 3f) and Nannoniscus cf. aequiremis (Fig. 4a) were only rarely observed north of the Ridge, but at intermediate depths. The remaining six species occurred either in shallow or deep waters of the Nordic Seas, i.e. at temperatures between <0 and <2.2°C and were found in at least two types of water masses; Cryodesma agnari (Fig. 3c), Desmosoma strombergi (Fig. 3d), Nannoniscoides angulatus (Fig. 3g) and Nannoniscus reticulatus (Fig. 3h), or were restricted to deeper cold waters (NSDW only; <0°C; Exiliniscus hanseni 1,130–1,648 m; Fig. 3e; and Torwolia creper, 1,642–2,069 m; Fig. 4b). Despite that E. hanseni and T. creper both occurred in deep waters and both at the same temperatures, their distributions differed considerably. E. hanseni occurred only to the north of Iceland, while T. creper occurred only in the deep waters northeast of Iceland, i.e. the former in the Iceland Sea, while the latter in the Norwegian Sea.
The largest group of species (14) was restricted to the south of the Ridge. The most common species, C. insignis (Fig. 5e), was found in shallow and deep waters southeast, south and southwest of Iceland (Table 2). This species was restricted to the warm waters masses found south of the Ridge, i.e. MNAW, LSW and ISOW and only found at temperatures from 2.07 to 7.6°C. Despite being present considerably shallower (about 40 localities more shallow than 600 m) than the sill depth (620 m) of the Greenland–Iceland Ridge, the species does not enter the cold waters north of the Ridge. Four species followed quite a similar pattern, i.e. occurring from quite shallow (upper ranges at 163–296 m) to deep waters, but not passing the channels of the Ridge, i.e. Nannoniscus cf. oblongus (Fig. 6f), Whoia angusta (Fig. 7b), Desmosoma lineare (Fig. 5f) and Rapaniscus crassipes (Fig. 6h), all occurring at temperature >2.3°C in MNAW, ISOW or LSW. E. aculeata (Fig. 5h) occurred only at shallow depths mainly southwest of Iceland between 200 and 300 m, with occasional findings from 136–778 m, at temperatures above 5.5°C and only in MNAW.
Several species occurred in deep water south and southwest of the Ridge and all but Mirabilicoxa gracilipes (upper limits 769 m; Fig. 6e) below the maximum sill depth of the GIF Ridge, i.e. 840 m), i.e. Eugerdella polita (Fig. 6a), Exiliniscus clipeatus (Fig. 6b), cf. Desmosoma tetarta (Fig. 5g), Mirabilicoxa acuminata (Fig. 6d), Nannonisconus sp. A (Fig. 6g), and Thaumastosoma platycarpus (Fig. 7a). Some of these species did not occur often in the samples, such as D. tetarta. M. gracilipes and E. polita occurred in all three water masses south of the Ridge (MNAW, LSW, ISOW), while the other species occurred in either ISOW or LSW.
Among the species occurring only to the south of the Ridge, patterns in the regional distribution differed. N. oblongus differed from other species in this group by occurring only to the west of the deep Reykjanes Ridge. On the other hand, M. acuminata and T. platycarpus occurred only to the east of the Reykjanes Ridge.
Only three species, i.e. Eugerda cf. globiceps (Fig. 4g), E. reticulata (Fig. 4h) and O. nanseni (Fig. 5b) were commonly found both north and south of the GIF Ridge. All of them had a wide bathymetrical distribution both north and south of the Ridge and occurred accordingly at a wide range of temperatures (Eugerda cf. globiceps, north: 97–2,613 m, south: 120–2,270 m, −0.86 to 8.04°C; O. nanseni, north: 209–1,558 m, south: 317–2,215 m, −0.86 to 7.12°C; E. reticulata, north: 227–3,003 m, south: 544–2,295 m, −0.9 to 4.57°C; Fig. 8) and in all seven water masses (i.e. MNAW, LSW, ISOW MEIW, A/PW, NSAIW and NSDW). No significant differences (p > 0.05) were found in the depth occurrences of these common species (occurrences at stations shallower than 350 m compared to occurrences at stations >350 m). In addition, Nannoniscus profundus (Fig. 5a) and Pseudomesus brevicornis (Fig. 5c) were found at both sides of the Ridge, but only at a few or several deep locations (Fig. 8).
A few species were found on both sides of the GIF Ridge, but in different patterns. Desmosoma latipes (Fig. 4c), Eugerdella armata (Fig. 4d) and Eugerdella hessleri (Fig. 4f) had most of their sites north of the Ridge. All these species were found from shallow to intermediate or deep waters, i.e. Eugerdella cf. armata at 149–910 m, D. latipes at 157–1,727 m and E. hessleri at 300–1,727 m and occurred accordingly in several water masses and at considerable temperature range (<0 to ≥6°C). E. tenuimana (Fig. 4e) was mainly found at the northern side of the Greenland–Iceland Ridge, but with additional localities south of the Iceland–Faeroe Ridge. This is a species occurring in quite shallow waters (104–558 m), but at a fairly wide temperature range (−0.47 to 5.4°C) and in four water masses. Prochelator lateralis (Fig. 5d) was mainly found south of the Ridge, but stretching into cold waters having been found at one location northwest of Iceland and to the north of the Greenland–Iceland Ridge.
Temperature ranges and water masses
Most of the species occurred in more than a single water mass. Only 11 species were restricted to single water mass (Table 2). Several of these were, however, found at only a few locations. Only four of the species found at more than four stations were restricted to single water mass, i.e. T. creper and E. hanseni in NSDW, M. acuminata in ISOW and E. aculeata in MNAW.
Some species occurred in several water masses. Most often, individual species occurred in water masses that were similar in temperature and salinity. In all, 15 species were “warm water species” occurring either in a single warm water mass or combinations of either one or two warm water masses (i.e. MNAW, LSW or ISOW) with temperatures above 2°C, some at a narrow range (range about 2°C), while others at had a fairly wide range (range about 5°C). The occurrence of species depending on temperature (°C) in water mass was tested for warm (>3°C) and cold (<3°). The test showed that the two groups were not significantly related (p = 0.647).
Despite the fact that the ISOW is formed north of the Ridge, its temperatures are above zero. Four species occured in this water mass only (E. clipeatus, M. acuminata, Nannonisconus sp. and T. platycarpus), while seven species occurred in combinations of MNAW, LSW and ISOW (temperatures >2.07°C), and five species occurred in a ISOW as well as warm and cold water masses.
Only eight species were confined to purely arctic water masses (A/PW, NSAIW or NSDW) ranging in temperatures from <0°C to maximally 1.41°C. Of these only four species were restricted to temperatures below 0°C (E. hanseni, Hebefustis sp. A, Nannoniscus cf. aequiremis and T. creper). All four species occurred in the deep Nordic Seas.
Patterns of species composition
The MDS plot distinguished the northern stations and the southern stations (Fig. 9a) The MDS plot did further distinguish the stations with warm water masses from the stations with cold water masses (Fig. 9b). The ANOSIM analysis revealed that the regions north and south of the Ridge were different from each other (Global R = 0.44, p < 0.001) and that stations with cold and warm temperatures were different from each other (Global R = 0.474, p < 0.001). Individual water masses were, however, poorly distinguished from each other in the MDS plot. Depth and sediment types were not found to be important in explaining observed patterns. A few stations located both north and south of the Ridge were distinguished in both plots from the bulk of the stations (Fig. 9a, b). These stations are located at various depths and with different water masses, i.e. NSDW, MNAW or CW. Four stations were northern stations below 1,844 m and the deepest one being at 2,069 m in NSDW. Stations with MNAW were at depths of 425 and 558 m, and the station with CW was at 104 m depth.

Multivariate analyses of the species composition of all sites: Two-dimensional MDS configuration based on Bray–Curtis similarities. a Species abundance plotted with the factor of the region; b species abundance plotted with the factor water mass
Species richness patterns
The number of desmosomatid and nannoniscid species collected varied considerably (by an order of magnitude) between samples, ranging from one to 12 species per sample. The patterns in number of species per sled with increasing depth differed considerably between the areas north and south of the Ridge (Fig. 10). North of the Ridge the number of species ranged from one to 12, with several stations having ≥6 species at the depth range from about 300 to about 1,700 m. Below 1,800 m only a single species occurred at each of the stations. South of the Ridge, species numbers were similar at all depths, while the two stations with the highest species richness occurred at depths below 2,000 m.

Number of species with depth (m). a South of the GIF Ridge; b north of the GIF Ridge
Discussion
The Nordic Seas and the Arctic Ocean are in a way semi-isolated from other oceans. Only the shallow Bering Strait connects the Arctic Ocean and the North Pacific, while the shallow GIF Ridge separates the deep basins of the Nordic Seas and the Arctic Ocean from the North Atlantic proper. The topographical evolution of the GIF Ridge is related to the development of the oceanic circulation of the North Atlantic (Bott 1983). Today, the GIF Ridge plays a decisive role in separating water masses of different climatological origin: the Atlantic waters with high salinities and high temperatures and the Polar waters with low temperatures and low salinities (Meincke 1983; Hansen and Østerhus 2000). The GIF Ridge was formed in the time interval 57–16 m.y. ago (Larsen 1983), the youngest part being at the spreading axis through Iceland (Bott 1983). In times of a lower sea level, the western part of the GIF Ridge was apparently a continuous land bridge and has probably interrupted the water exchange between the Atlantic proper and the Nordic Seas (Thiede 1983; Larsen 1983). The Iceland–Faeroe Ridge has probably subsided below sea level 24–14 m.y. ago (Berggren and Schnitker 1983).
The Nordic Seas and the Arctic Ocean have undergone large changes in the temperatures, changing from fairly high temperatures in the past to cold waters at the beginning of the glaciations 2.5 m.y. ago. The structure and the development of the GIF Ridge may explain the patterns observed today for the isopod fauna around Iceland. The pattern seen may partly be historical and partly due to present ecological conditions. The northernmost part of the North Atlantic and the Nordic Seas, where the Atlantic proper meets the Nordic Seas and the Arctic Ocean, holds a diverse component of the desmosomatid and nannoniscid isopods, with an even dispersal of the species. This may indicate that the distributional pattern of the species around Iceland is more due to present ecological conditions rather than historical events. The deep water basins of the Nordic Seas and the Arctic Ocean may, have been strongly influenced by glaciations during the last few million years (Dahl 1972). Indeed one of the strongest signals in these and other data is the considerable drop in diversity with increasing depth north of the GIF Ridge, whereas south of it and elsewhere in the open Atlantic (Svavarsson 1997; Wilson 1998). Bouchet and Warén (1979) concluded though that a part of the molluscs fauna of the Norwegian Sea was a reminiscence of the old Pacific fauna that lived in the Polar Basin before it was disconnected from the North Pacific by the formation of the Bering Strait.
Pronounced differences were seen in the distributions of the isopods in the area, and most of the species were restricted either to the cold waters north of the Ridge or to the warm waters south of the GIF Ridge. This indicates that the distributions are not only shaped by the temperature (ANOSIM: R 0.474, p 0.001), also by the actual presence of the Ridge itself (ANOSIM: Global R 0.44, p 0.001). This is supported by the fact that several of the species occur shallower than the actual sill depths of the Ridge and could pass the Ridge if the temperatures were suitable. Therefore, the physical presence of the Ridge itself may act secondarily as barrier, having though in mind that the Ridge shapes the temperatures of the area. Similar argumentation has been provided for the isopod Astacilla boreaphilis Stransky and Svavarsson 2006, which occurs only in the waters to the south of the Ridge, but the species occurs considerably shallower than the sill depths of the Ridge (Stransky and Svavarsson 2006). Numerous other studies have shown pronounced limits in species distributions at the Ridge (e.g. Negoescu and Svavarsson 1997; Gudmundsson 1998; Weisshappel 2000, 2001; Dijkstra et al. 2009).
Most of the isopod species occurred in several water masses, and only a few common species were restricted to single water mass. The physical differences between several of the water masses are quite small and even though occurring in more than one water mass, a species may be adapted to a narrow range of temperatures as the small differences are not sufficient to limit distributions. Most species were, however, restricted to either cold or warm water masses, while a few species, i.e. the species occurring on both sides of the Ridge, where able to deal with a wide range of temperatures. The presence of cryptic species cannot be excluded for such species occurring within so wide ranges of temperatures, e.g. Eugerda cf. globiceps and O. nanseni.
Differences were observed in the distribution of several species within the Nordic Seas (e.g. E. hanseni vs. T. creper) and for some species in the Atlantic proper (e.g. M. acuminata vs. Nannoniscus cf. oblongus). These differences are most like due to ecological conditions, other than the temperature. Sediment had not much influence on the distributions (BIOENV analysis). Weisshappel and Svavarsson (1998) showed that the sediment structure shaped the distribution of amphipods only when the temperature was excluded. Other plausible explanations for these differences are differences in the food supply, competition, disturbance, etc., which can act at local scales (Gray 1974; Pearson and Rosenberg 1987; Thrush et al. 2005).
With most of the species occurring to the south of the Ridge, the desmosomatids and nannoniscids are similar to most other studied groups in the area, showing the same pattern, e.g. anthurid isopods (Negoescu and Svavarsson 1997) and eusirid amphipods (Weisshappel 2000). Calliopiid amphipods have, however, a different pattern with most of their species occurring to the north of the Ridge (Weisshappel 2001).
A low regional diversity has been demonstrated for several groups of invertebrates, e.g. echinoderms (Sibuet 1979) and molluscs (Bouchet and Warén 1979), of the Arctic deep water. A low regional diversity has previously been shown for the asellote isopods of the deep Arctic Ocean (Gurjanova 1938) and the deep Nordic Seas (Svavarsson et al. 1990; Svavarsson 1997). The low diversity of the deep waters of the Nordic Seas has been partly explained due to the presence of the Ridge itself, i.e. isolation of the deep basins of the Nordic Seas and the Arctic Ocean from the basins of the North Atlantic (e.g. Svavarsson et al. 1993; Rex et al. 1993, 2000; Wilson 1998).
This is based on the argument that various groups of invertebrates are differently represented in the Nordic Seas/Arctic Ocean and the North Atlantic south of the Ridge (Svavarsson et al. 1993). Despite being a deep-sea family the Desmosomatidae has a fair amount of their known species (46) occurring in the Nordic Seas. However, some of the other deep-sea asellote isopod families seem to have much lesser representation in the Nordic Seas, despite being species rich just south of the Ridge. Only two species of the Haploniscidae, a family with over 100 known species worldwide, occur in the Nordic Seas (Svavarsson et al. 1993) while several species of this family were collected at the RP sledge stations on the GIF Ridge during the BIOICE project. The highest number of haploniscid specimens per station collected was 4,259 specimens of Haploniscus bicuspis (Sars 1877) (BIOICE station 2014, North of Iceland). However, this may not fully explain the low diversity and a lack of adaptation towards subzero temperatures needs to be addressed.
A decline in diversity with depth has been shown for asellote isopods in the eastern and western part of the Nordic Seas (Svavarsson et al. 1990; Svavarsson 1997). In the present study, the pattern seen in species richness differed considerably between the areas south and north of the Ridge. North of the Ridge the highest numbers of species per station (5–12 species/sled) were at shallow and intermediate depths (around 300 to >1,600 m) and the species richest stations were either in shallow warm (>0°C) or intermediate cold (−0.51 to −0.76°C) waters. Below depths of around 1,700 m only a single species occurred per station north of the Ridge. Similar pattern (high diversity at cold temperatures, but low diversity in the basins of the Arctic Ocean and the Nordic Seas) has also been shown for the amphipods (Weisshappel 2000, 2001). Renaud et al. (2006) found, however, no changes in nematode diversity with depth in the central Arctic Ocean. As fairly high diversity occurs at those cold temperatures, this implies that the low regional diversity of the deep Arctic Ocean and the Nordic Seas may not caused by a lack of adaptations to subzero temperatures. Extinction events in the past may play a role and other environmental factors, such as food availability, may shape the diversity patterns.
Interestingly, few species that occur on the Norwegian coast, e.g. N. oblongus and R. crassipes, are found only on the Atlantic side of Iceland. These species are shallow water species which are also recorded from the Greenland shelf. In the case of N. oblongus, we speak of a species with a wide distribution from Greenland over the south of Iceland, the Faroes, the southern part of Norway to the northern part of Norway. Most probably temperature is the limit for distribution at the GIF Ridge while the coastal waters in Norway offer the species good conditions.
In all, 13 species occurred between relatively shallow waters (≤300 m) and considerable depths (>1,000 m). Only a few ubiquitous species, i.e. E. aculeata, E. tenuimana and Eugerdella cf. armata, were restricted to shallow water while eight species occurred only at depths greater than 840 m. While shallow waters were more frequently sampled than the deep waters and thus shallow water species seem to be ubiquitous, they are actually no more common than species in deep waters.
Some desmosomatid species occur in bathyal waters south-east of the Greenland–Iceland–Faeroe Ridge although being usually found in shallower waters in other subarctic regions. The circumicelandic species Eugerda cf. globiceps extends deeper north of the Ridge as well as E. reticulata and O. nanseni. For these three circumicelandic species (showing a wide bathymetric range and occurring at more than 50 stations) it was tested if there is no difference of occurrence in depth (“shallow” <350 m; “deep” >350 m). No difference was found. Although bathymetrical distribution of water masses and hence temperature may differ considerably within the Arctic Ocean and the Nordic Seas and hence lead to shallow occurrences of a species in some areas but deeper occurrences of the same species in other areas, this is not the case for Eugerda cf. globiceps, E. reticulata and O. nanseni.
In the deep-sea basins south of Iceland, i.e. the North Atlantic proper, species were found that were recorded by Hessler (1970a) and Siebenaller and Hessler (1977, 1981) from the Bermuda Transect (in the Irminger Basin D. tetarta and Hebefustis cf. vafer; in the Iceland Basin E. clipeatus, M. acuminata, Nannonisconus cf. sp. nov. and T. platycarpus), indicating that some of the species having their northern limits of distribution at the GIF Ridge may extend their distribution far south. Meland and Brattegard (2007) found mysid species south of Iceland previously reported from the Antarctic and close resemblance of Antarctic and North Atlantic species. Such wide distribution raises a question of presence of cryptic species. Brökeland and Raupach (2008) showed for species of Haploniscidae that genetic distances in mitochondrial markers go along with morphological characters in a complex of sibling species.
In particular, the observation above suggests that the ecological differences between the Atlantic and Arctic sectors may be influencing the distributions. Answering the questions from the beginning, we can conclude:
Most isopod species around Iceland are restricted to water masses of a certain temperature and salinity. The topography of the GIF Ridge does not shape the distribution of asellote isopods as much as the abiotic factors temperature and salinity. Sediment or depth were not found to limit the species distribution in the area.
References
Berggren WA, Schnitker D (1983) Cenozoic marine environments in the North Atlantic and Norwegian–Greenland Sea. In: Bott MHP, Saxov S, Talwani M, Thiede J (eds) Structure and development of the Greenland–Scotland Ridge new methods and concepts. Springer, US, pp 495–548
Bott MHP (1983) Deep structure and geodynamics of the Greenland–Scotland Ridge: an introductory review. In: Bott MHP, Saxov S, Talwani M, Thiede J (eds) Structure and development of the Greenland–Scotland Ridge new methods and concepts. Springer, US, pp 3–10
Bouchet P, Warén A (1979) The abyssal molluscan fauna of the Norwegian Sea and its relation to other faunas. Sarsia 64:211–243
Brandt A, Brökeland W, Brix S, Malyutina M (2004) Diversity of Southern Ocean deep-sea Isopoda (Crustacea, Malacostraca)—a comparison with shelf data. Deep-Sea Res II 51:1753–1768
Brandt A, Brix S, Brökeland W, Cedhagen T, Choudury M, Cornelius N, Danis B, De Mesel I, Diaz RJ, Gillan DC, Hilbig B, Howe J, Janussen D, Kaiser S, Linse K, Malyutina M, Brandao S, Pawlowski J, Raupach M, Gooday AJ (2007) The Southern Ocean deep sea: first insights into biodiversity and biogeography. Nature 447:307–311
Brattegard T, Fosså JH (1991) Replicability of an epibenthic sampler. J Mar Biol Ass UK 71:153–166
Brix S (2006a) A new genus and new species of Desmosomatidae (Crustacea: Isopoda: Asellota) from the deep sea of south-eastern Australia. Mem Mus Vic 63:175–205
Brix S (2006b) A new species of Desmosomatidae (Isopoda: Crustacea) from the deep Southern Ocean: Eugerdella serrata sp. nov. including remarks to the morphological variability within Eugerdella Hessler, 1970. Mitt Zool Mus Inst 103:69–84
Brix S (2007) Four new species of Desmosomatidae Sars, 1897 (Crustacea: Isopoda) from the deep sea of the Angola Basin. Mar Biol Res 3:205–230
Brökeland W, Raupach M (2008) A species complex within the isopod genus Haploniscus (Crustacea: Malacostraca: Peracarida) from the Southern Ocean deep sea: a morphological and molecular approach. Zool J Linn Soc 152:655–706
Clarke KR, Ainsworth M (1993) A method of linking multivariate community structure to environmental variables. Mar Ecol Prog Ser 92:205–219
Clarke KR, Warwick RM (2001) Change in marine communities: an approach to statistical analysis and interpretation. Primer-E Ltd., Plymouth
Dahl E (1972) The Norwegian Sea deep water fauna and its derivation. Ambio Spec Rep 2:19–24
Dijkstra HH, Warén A, Gudmundsson G (2009) Pectinoidea (Mollusca: Bivalvia) from Iceland. Mar Biol Res 5:207–243
Gray JS (1974) Animal-sediment relationships. Oceanogr Mar Biol Ann Rev 12:707–722
Gudmundsson G (1998) Distributional limits of Pyrgo species at the biogeographic boundaries of the Arctic and the North-Atlantic Boreal regions. J Foraminifer Res 28:240–256
Gurjanova E (1938) On the question of the composition and origin of the abyssal fauna of the Polar Basin. Comp Rend (Doklady) l’Acad Sci l’URSS 20:333–336
Gurjanova E (1946) Novye vidy Isopoda i Amphipoda iz Severnogo Ledovitogo okeana. Trudy Dreifuiushchaia ekspeditsii Glavsevmorputi na ledokol‘nomparakhode “G. Sedov” 1937–1940 3:272–297
Hansen HJ (1916) Crustacea Malacostraca III, Isopoda. Dan Ingolf Exped 3:1–262
Hansen B, Østerhus S (2000) North Atlantic-Nordic Seas exchanges. Prog Oceanogr 45:109–208
Hessler RR (1970a) The Desmosomatidae (Isopoda, Asellota) of the Gay Head-Bermuda transect. Bull Scripps Inst Oceanogr 15:1–185
Hessler RR (1970b) High-latitude emergence of deep-sea isopods. USA Antarctic Res J 5:133–134
Hessler RR, Strömberg J-O (1989) Behavior of janiroidean isopods (Asellota), with special reference to deep-sea genera. Sarsia 74:145–159
Hult J (1941) On the soft-bottom isopods of the Skagerrak. Zool bidr Uppsala 21:1–234
Just J (1970) Decapoda, Mysidacea, Isopoda, and Tanaidacea from Jørgen Brønlund Fjord, North Greenland. BioScience 84:1–32
Just J (1980) Polar Sea abyssal and deep bathyal Isopoda (Crustacea). Steenstrupia 6:197–230
Kaiser S, Brix S (2005) A new isopod species from the Southern Ocean: Disparella maiuscula sp nov. (Isopoda: Asellota: Desmosomatidae). Mitt Hamb zool Mus Inst 102:153–165
Kaiser S, Brix S (2007) Two new species of Pseudomesus Hansen, 1916 (Isopoda: Asellota: Desmosomatidae) Pseudomesus pitombo sp nov. and Pseudomesus satanus sp. nov. Zootaxa 1658:21–38
Larsen B (1983) Geology of the Greenland-Iceland Ridge in the Denmark Strait. In: Bott MHP, Saxov S, Talwani M, Thiede J (eds) Structure and development of the Greenland-Scotland Ridge new methods and concepts. Springer, US, pp 425–444
Malmberg S-A, Valdimarsson H (2003) Hydrographic conditions in Icelandic waters, 1990–1999. ICES Mar Sci Symposia 219:50–60
Meincke J (1983) The modern current regime across the Greenland-Scotland Ridge. In: Bott MHP, Saxov S, Talwani M, Thiede J (eds) Structure and development of the Greenland-Scotland Ridge new methods and concepts. Springer, US, pp 637–652
Meland K, Brattegard T (2007) New Mysida (Crustacea) in the genera Amblyops and Pseudomma from the Iceland Basin. Zootaxa 1628:43–58
Menzies RJ, Mohr JL (1962) Benthic Tanaidacea and Isopoda from the Alaskan Arctic and the polar Basin. Crustaceana 3(3):192–202
Negoescu I, Svavarsson J (1997) Anthurideans (Crustacea, Isopoda) from the North Atlantic and the Arctic Ocean. Sarsia 82:159–202
Paul AZ, George RY (1975) High Arctic benthic isopods from Fletchers’s Ice Island, T–3, with a description of one new species, Mirabilicoxa fletcheri n sp. Crustaceana 29:166–168
Pearson TH, Rosenberg R (1987) Feast and famine: structuring factors in marine benthic communities. In: Gee JHR, Giller PS (eds) Organisation of communities past and present. Blackwell Scientific Publications, Oxford, pp 373–395
Poore GCB, Just J, Cohen BF (1994) Composition and diversity of Crustacea (Isopoda) of the Southern Australian Continental Slope. Deep Sea Res Part I 41:677–693
Renaud PE, Ambrose WG Jr, Vanreuseld A, Cloughe LM (2006) Nematode and macrofaunal diversity in central Arctic Ocean benthos. J Exp Mar Biol Ecol 330:297–306
Rex MA, Stuart LT, Hessler RH, Allen JA, Sanders HL, Wilson GDF (1993) Global-scale patterns of species diversity in the deep sea. Nature 365:636–639
Rex MA, Stuart CT, Coyne G (2000) Latitudinal gradients of species richness in the deep-sea benthos of the North Atlantic. Proc Natl Acad Sci USA 97:4082–4085
Rothlisberg PC, Pearcy WG (1977) An epibenthic sampler used to study the ontogeny of vertical migration of Pandalus jordani (Decapoda, Caridea). Fish Bull 74:994–997
Sars GO (1899) Isopoda, an account of the Crustacea of Norway, Bergen Museum. Bergen 2:1–270
Schotte M, Kensley BF, Shilling S (2009) World list of marine, freshwater and terrestrial Crustacea Isopoda. National Museum of Natural History Smithsonian Institution: Washington, DC, USA http://www.nmnh.si.edu/iz/isopod/. Accessed 1 July 2009
Schuchert P (2000) Hydrozoa (Cnidaria) of Iceland collected by the BIOICE programme. Sarsia 85:411–438
Sibuet M (1979) Distribution and diversity of asteroids in Atlantic abyssal basins. Sarsia 64:85–91
Siebenaller JF, Hessler RR (1977) The Nannoniscidae (Isopoda, Asellota): Hebefustis n gen. and Nannoniscoides Hansen. Trans San Diego Soc Nat Hist 19:17–44
Siebenaller JF, Hessler RR (1981) The genera of the Nannoniscidae (Isopoda, Asellota). Trans San Diego Soc Nat Hist 19:227–250
Stefánsson U (1962) North Icelandic waters. Rit Fiskideildar 3:1–269
Stephensen K (1937) Marine Isopoda and Tanaidacea. Zool Iceland 3:1–26
Stephensen K (1943) The zoology of East Greenland—Leptostraca, Mysidacea, Cumacea, Tanaidacea, Isopoda and Euphausiacea. Medd Grønland 121:1–82
Stransky B, Svavarsson J (2006) Astacilla boreaphilis sp nov. (Crustacea: Isopoda: Valvifera) from shallow and deep North Atlantic waters. Zootaxa 1259:1–23
Svavarsson J (1982) Nannoniscus profundus sp n. and Austroniscus norbi sp. n. (Isopoda, Asellota, Nannoniscidae) from the deep Norwegian Sea. Sarsia 67:179–186
Svavarsson J (1984) Description of the male of Pseudomesus brevicornis Hansen, 1916 (Isopoda, Asellota, Desmosomatidae) and rejection of the family Pseudomesidae. Sarsia 69:37–44
Svavarsson J (1988) Desmosomatidae (Isopoda, Asellota) from bathyal and abyssal depths in the Norwegian, Greenland, and North Polar Seas. Sarsia 73:1–32
Svavarsson J (1997) Diversity of isopods (Crustacea): new data from the Arctic and Atlantic Oceans. Biodiversity Conserv 6:1571–1579
Svavarsson J, Brattegard T, Strömberg J-O (1990) Distribution and diversity patterns of asellote isopods (Crustacea) in the deep Norwegian and Greenland Seas. Prog Oceanogr 24:297–310
Svavarsson J, Strömberg J-O, Brattegard T (1993) The deep-sea asellote (Isopoda, Crustacea) fauna of the Northern Seas: species composition, distributional patterns and origin. J Biogeogr 20:537–555
Thiede J (1983) Outstanding geological problems of the Greenland–Scotland ridge: an introduction. In: Bott MHP, Saxov S, Talwani M, Thiede J (eds) Structure and development of the Greenland–Scotland Ridge new methods and concepts. Springer, US, pp 313–318
Thrush SF, Hewitt J, Herman PMJ, Ysebaert T (2005) Multi-scale analysis of species–environment relationships. Mar Ecol Prog Ser 302:13–26
Valsangkar AB (2004) Clay as an indicator of sediment plume movement in deep-sea environment. Curr Sci 87:1747–1751
Weisshappel JBF (2000) Distribution and diversity of the hyperbenthic amphipod family Eusiridae in the different seas around the Greenland–Iceland–Faeroe-Ridge. Sarsia 85:227–236
Weisshappel JBF (2001) Distribution and diversity of the hyperbenthic amphipod family Calliopiidae in the different seas around the Greenland–Iceland–Faeroe-Ridge. Sarsia 86:143–151
Weisshappel JBF, Svavarsson J (1998) Benthic amphipods (Crustacea: Malacostraca) in Icelandic waters: diversity in relation to faunal patterns from shallow to intermediate deep Arctic and North Atlantic Oceans. Mar Biol 131:133–143
Wilson GDF (1998) Historical influences on deep-sea isopod diversity in the Atlantic Ocean. Deep Sea Res II 45:279–301
Wilson GDF (2008a) A review of taxonomic concepts in the Nannoniscidae (Isopoda, Asellota), with a key to the genera and a description of Nannoniscus oblongus Sars. Zootaxa 1680:1–24
Wilson GDF (2008b) Local and regional species diversity of benthic Isopoda (Crustacea) in the deep Gulf of Mexico. Deep-Sea Res Part II 55:2634–2649
Wolff T (1962) The systematics and biology of bathyal and abyssal Isopoda Asellota. Galathea Rep 6:1–320
Acknowledgments
We would like to thank Torleiv Brattegard for his hospitality on board RV Håkon Mosby, MRI, Reykjavík, for the use of RV Bjarni Sæmundsson and the Fiskirannsóknastova, Torshavn, for the use of RV Magnus Heinason. We would like to thank Sigrún Haraldsdóttir and the staff of the Sandgerði Marine Centre for sorting the isopods. Ólafía Lárusdóttir kindly helped in making the maps and in designating the water masses. Stefanie Kaiser, Niel Bruce, Christoffer Schander and an anonymous reviewer improved the manuscript considerably with their constructive comments. Bente Stransky helped with the Primer analysis. Eric de Santos improved the English. We acknowledge grants received from the Icelandic Centre for Research, the University of Iceland Research Fund and the German Science Foundation (DFG) under contract No. Br 1121/28-1.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Brix, S., Svavarsson, J. Distribution and diversity of desmosomatid and nannoniscid isopods (Crustacea) on the Greenland–Iceland–Faeroe Ridge. Polar Biol 33, 515–530 (2010). https://doi.org/10.1007/s00300-009-0729-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-009-0729-8