
Abstract
Drepanocladus longifolius (Mitt.) Paris is recorded for the first time from King George Island, South Shetland Islands, in the maritime Antarctic. It was collected in West Lake during the 23rd Chinese National Antarctic Research Expedition in 2006–2007. The moss was found at a depth of 5–6 m attached to the bed of the lake. The stems of the moss are about 1–1.5 m in length. The moss exhibits seasonal growth patterns, with shorter branch internodes, more widely spaced leaves and more branches in summer than in winter. Most of the branches are initiated in summer. The annual shoot extension is about 3–6 cm, which implies that the plants must be at least 15 years of age. The distribution of aquatic moss species and records in Antarctica is outlined and discussed and the nomenclature of previous reports clarified.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The Antarctic continent occupies about 14.4 million km2, more than 99% of which is covered by ice with an average thickness of around 1.8 km. Lichens and mosses are the dominant components of the terrestrial vegetation in Antarctica, which comprises a poorly developed tundra-like vegetation. 175 years of bryological exploration of the continent and adjacent areas, including the conterminous archipelagoes of the South Sandwich, South Orkney and South Shetland Islands, began with an account of Polytrichastrum alpinum (Hedw.) G.L.Sm. from King George Island, South Shetland Islands (Eights 1833). Although over 300 taxa of mosses have been reported from this continent, the recent comprehensive monographic study of the mosses of this biome has reduced this number to 111 species and two varieties belonging to 17 families and 55 genera (Ochyra et al. 2008).
The richest moss flora is in the South Shetland Islands where 87 species and one variety have been recorded. This is a large archipelago of about 15 islands situated about 160 km north of Trinity Peninsula, the northernmost tip of the Antarctic Peninsula, and approximately 770 km south-east of Cape Horn. The greatest diversity of mosses in this archipelago is on King George Island, the largest island in the group, with an area of ca. 1,300 km2 and with nearly 10% ice-free ground. 63 species have been recorded there, making it bryologically the richest locality within the Antarctic biome. The island’s bryoflora has been intensively investigated, and there is a relatively wide range of terrestrial and aquatic habitats. There are many streams, pools and lakes, particularly on Fildes Peninsula in the south-western part of the island. The lakes, some of which are up to 10 m in depth, have been rather poorly investigated, but Campylium polygamum (Schimp.) Lange et C.E.O.Jensen has been reported from Lake Kitezh in shallow water (Ochyra 1998). We now report the occurrence of the rare aquatic moss Drepanocladus longifolius (Mitt.) Paris from West Lake. It represents a remarkable addition to the moss flora of King George Island, which now totals 64 species. Other areas in the Antarctic biome with a large moss flora include Signy Island in the South Orkney Islands (59 species), and Livingston Island (56) and Deception Island (54), both in the South Shetland Islands.
Materials and methods
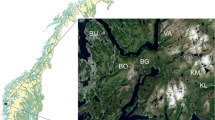
Specimens of Drepanocladus longifolius were collected during the 23rd Chinese National Antarctic Research Expedition in 2006–2007 at 5–6 m depth in West Lake (62°13′01(S, 58°57′57″W). We used a rubber boat and a rake with long handle to collect the specimens. West Lake is situated in the southern part of Fildes Peninsula on King George Island (Fig. 1), and it is the principal source of freshwater for China’s Great Wall Station (62°12′59″S, 58°57′52″W). The catchment area of West Lake is about 16,000 m2 and the maximum lake depth is about 10 m.

The location of sampling site in West Lake, Fildes Peninsula
The specimens, which are deposited in the Chinese National Herbarium, Institute of Botany, Chinese Academy of Sciences, Beijing (PE, identification number: 01745091) with a small duplicate preserved in the Bryological Herbarium of the Institute of Botany of the Polish Academy of Sciences in Kraków (KRAM, accession number: 179709), were examined with an SMZ1000 stereomicroscope, and the structures of their stems and leaves measured using a Leica DM2500 microscope by the first author.
Hydrological data of West Lake are showed in Table 1, which is supplied by Dr Juan Wang, College of Environmental Science and Engineering, Tongji University, China. The water temperature, pH, ORP, SpCond, Sal, Depth and DO values are detected by HydroLab DS 5X water quality multiprobes in the lake. The surface water of 250 ml for each sample was collected from the lake by plexiglass water sampler, and then the samples were examined in the laboratory and the NH4 +-N, NO3 −-N, NO2 −-N, TP values were determined by HACH DR 2800 portable spectrophotometer.
Results and discussion
Specimens descriptions
Drepanocladus longifolius is a very distinct and characteristic species in Antarctica (Figs. 2, 3). It is the only pleurocarpous moss in this biome having lanceolate and long acuminate leaves with a fairly strong, long-excurrent costa which may occasionally be percurrent on branch leaves. Because of deep submergence, the Antarctic plants are morphologically highly modified in comparison with typical phenotypes from the South American mainland and Tierra del Fuego, where the plants grow in swampy habitats but are seldom submerged. The species is clearly differentiated from other pleurocarpous mosses in the region by its alar cells which form small auricles in the basal angles of the leaves.

Sketch of morphology and anatomy of Drepanocladus longifolius. 1 portion of plant; 2 cross section of stem; 3 portion of the cross section of stem; 4, 5 leaves; 6 apical leaf cells; 7 median marginal leaf cells; 8 alar cells (drawn from C.·S.Li 0701)

Pictures showing the morphological differences of Drepanocladus longifolius in consecutive winters and summers (arrows indicate the growth in different summers)
The King George Island plants of D. longifolius are slender, yellowish-green, and float freely in water, forming fairly dense mats. The stems are irregularly branched, rounded to elliptical in cross section, with a small central strand. Cortical cells of the stems are incrassate, in 2–3 layers. Medullary cells are large, 40–50 μm long and 4–6 μm wide, hyaline, with thin walls. Leaves are 1.5–4 mm long and 0.2–0.4 mm wide, erect-spreading, shortly and narrowly decurrent, flattened or slightly convolute, narrowly lanceolate, gradually narrowed into a long acumen, with entire margins. The costa is single and fairly strong, 60–70 μm wide at the base, long-excurrent or seemingly percurrent because of imperceptible merging with the cells in the acumen. Mid-leaf cells are elongate-rhomboidal to linear, thin-walled, 70–130 μm long, 4–8 μm wide, becoming shorter (40–60 μm long) and wider (8–10 to 8–12 μm wide) towards the base. Alar cells are rectangular, with thin- to somewhat thickened and yellowish walls, forming small, convex, rounded to oval groups occupying about 1/3 or less the leaf base. Branch leaves are similar to stem leaves but somewhat smaller.
Drepanocladus longifolius is a very distinctive moss which is unlikely to be mistaken for any other moss in Antarctica, although some aquatic expressions of two other amblystegialean mosses, Campylium polygamum and Warnstorfia sarmentosa (Wahlenb.) Hedenäs may be externally similar to it. Campylium polygamum is widespread around China’s Great Wall Station on Fildes Peninsula and some plants have been found floating in intricate mats in lakes (Chen et al. 1994; Ochyra 1998). Additionally, this species has been found in the South Orkney Islands, Livingston Island in the South Shetland Islands and on Alexander Island in the maritime Antarctic (Ochyra et al. 2008). The main differences between these two species relate to the leaf shape, costa and alar cells. The leaves in C. polygamum are ovate-lanceolate, gradually tapering from an ovate to rounded-triangular base to a long, slender and channeled acumen, the costa is single and extends to three quarters the leaf length or is shorter and forked, and the alar cells are enlarged and form transversely triangular to broadly ovate, somewhat convex groups, extending from the margin to two-thirds of the leaf insertion or to the costa. Conversely, D. longifolius has narrowly lanceolate, gradually long acuminate leaves, with a single, long-excurrent costa and alar cells which form a small, rounded to oval group occupying one-third or less the width of the leaf base.
Distribution
Drepanocladus longifolius is a pan-south-temperate species with its main center of distribution in southern South America, including western and southern Patagonia, Tierra del Fuego and the Falkland Islands (Ochyra and Matteri 2001), extending along the Andean chain at high elevations from Bolivia to Colombia (Hedenäs 2003). It also occurs on some subantarctic islands including South Georgia (Ochyra et al. 2002), Îles Crozet (Ochyra, personal observations) and Îles Kerguelen (Hedenäs 1997; Ochyra, personal observations) and extends to south-eastern Australia and the northern maritime Antarctic. In the Antarctic, the species is known from Signy Island, South Orkney Islands, and Livingston Island, South Shetland Islands, as well as from James Ross Island and Vega Island on the eastern coast of the Antarctic Peninsula (Fig. 4). At all Antarctic sites the species grows in very similar habitats, being deeply submerged in water of lakes. Generally, the habitats of D. longifolius are more or less mineral- and nutrient-rich, the plants are often submerged in lakes, pools and swamps, and sometimes they thrive in periodically dry pools, small streams and springs (Hedenäs 1997; Ochyra and Matteri 2001).

Current distribution of Drepanocladus longifolius in the Antarctic. New locality on King George Island marked with the filled triangle
Habitat descriptions
West Lake is located in the western part of Great Wall Station. The climate parameters around the station include the mean annual temperature of −2.1°C, the maximum temperature of +11.7°C in January, the minimum temperature of −27.7°C in September. The mean temperature from November to March, which is considered as summer period, is +0.8°C; the mean temperature from April to October considered as the winter period, is −4.3°C. The mean annual precipitation is 517.2 mm, falling mostly as snow (based on data from Great Wall Station, China from 1985 to 2000). The thickness of the ice covering the lake in winter is ca. 1–1.2 m. The mean annual temperature of the lake water is about 4.5°C, with the mean temperature of 5.04°C during the summer period and 4.18°C during the winter period (based on data from Great Wall Station, China in 2007). Hydrological data for West Lake are summarized in Table 1, and the water conductivity is low (0.1 ms/cm), with pH ranging between 7.2 and 8.2.
Annual shoot extensions
Mosses have been well documented growing submerged in shallow areas of lakes and even in deep-water in Antarctica (e.g. Light and Heywood 1973; Kaspar et al. 1982; Seppelt 1983; Pickard and Seppelt 1984; Imura et al. 2003; Wagner and Seppelt 2006). When submerged, mosses may change their form considerably (Priddle 1979; Seppelt 1983; Seppelt and Selkirk 1984; Kanda and Ohtani 1991), with stems from aquatic forms being much more elongated than those of terrestrial plants. Plants of D. longifolius were exceptionally long and measured 1–1.5 m. This is the largest size recorded for this species and is amongst the longest stems ever reported in mosses. Fontinalis dalecarlica Schimp. from Amchitka Island in the Aleutian Islands, have been measured at up to 166 cm in length (Takaki 1985).
Priddle (1979) showed that the annual extensions of moss shoots in aquatic habitats could be estimated using innate growth markers. The moss plants produced leaves of different sizes and the branch internodes distances differed with season. This phenomenon was reported also in Lake Grane Langsø, Denmark (56°01′05(N, 9°27′06″E), where Sphagnum produces longer (and more) side branches during summer than winter, and Drepanocladus produces a higher density of side branches along the main shoot axis in summer than in winter (Riis and Sand-Jensen 1997). The specimens of D. longifolius from Fildes Peninsula also displayed a distinct pattern of seasonal growth (Fig. 3). The stems have shorter internodes and more densely arranged leaves in summer. Summer leaves are about 3 mm long, while winter leaves are 1.5 mm in length. The leaf color in winter is slightly paler than in summer. Most of the side branches are produced in summer. The annual extension of shoots was measured to be about 3–6 cm (4.35 cm on average) based on ten specimens studied. The apparently active living part of the stem was measured to be about 20 cm long. We inferred from our measurements that stems which are about 1–1.5 m in length have been submerged in water for more than 15 years. The extension of the shoot in summer is about 2.71 cm and 1.64 cm in winter on average, so the mean daily rate of shoot extension is about 0.3 mm in summer and 0.06 mm in winter.
Moss collected in water at 6–10 m depth in Lake Grane Langsø (56°01′05″N, 9°27′06″E) in Denmark had a growth rate of 9–25 cm of shoot per year, but a relatively short longevity of 0.7–2.9 years (Riis and Sand-Jensen 1997). Mosses from West Lake have a slower growth rate and greater longevity compared with the mosses in Lake Grane Langsø. It is apparent that at the same depth, the temperature and light intensity are lower in West Lake than in Lake Grane Langsø. Annual growth rate and longevity of aquatic mosses are related to water temperature and light intensity. With the decrease of the water temperature and light intensity, the annual growth rate and longevity will decrease simultaneously.
Possibilities of the origin of the aquatic mosses
Drepanocladus longifolius is the only aquatic moss known in West Lake and it has not been observed in the terrestrial habitats around the lake. Elsewhere in Antarctica aquatic mosses are also generally found on the surrounding land. The only apparent exceptions are Pohlia wilsonii (Mitt.) Ochyra in the Skarvsnes region of the Sôya Coast, Enderby Land (Imura et al. 2003, as Leptobryum sp.) and Plagiothecium orthocarpum Mitt. from the Schirmacher Oasis, Dronning Maud Land (Bednarek-Ochyra et al. 1999, as P. georgicoantarcticum (Müll.Hal.) Kindb.).
There are three possible explanations for the presence of these aquatic mosses in Antarctic lakes. Firstly, D. longifolius may have been accidentally introduced by human occupation of the area on King George Island. Secondly, it may have been carried to Antarctica by air currents from conterminous regions of southern South America or subantarctic South Georgia, as Kanda and Mochida (1992) suggested for Pohlia wilsonii. Thirdly, the population of this species may represent a relict which could have persisted from earlier times only in the lake but has disappeared from the surrounding land. The latter hypothesis seems to be reinforced by the discovery of D. longifolius in the mid-Miocene sediments from the Olympus Range in the McMurdo Dry Valley in Victoria Land (Lewis et al. 2008).
Light intensity is attenuated at the lake bottom by the depth of the water column, turbidity, surface ice and snow, and may be significantly lower than the level of irradiance at the adjacent land surface (Kudoh et al. 2003a). In Kuwai Ike, Sôya Coast (69°28′36″S, 39°38′48″E), many aquatic moss pillars form on lake bottoms at 2–5 m depth, where the light intensity was measured to be 50% of that at the ground level during the ice-free season (Imura et al. 2003). During the ice-covered season, the light intensity is further reduced to <10% (Kudoh et al. 2003b). Light and Heywood (1973) indicated that less than 1% of the total amount of visible incident light may reach the ice-water interface during the 6 months of winter. Because the sunshine duration in West Lake is low, about 499 h per year, the total irradiance per year is calculated at 1,932 MJ/m2. Maximum solar radiation at Great Wall Station is from November to January, which is 53% of the total solar radiation (Bian et al. 1992). We calculated that the lowest radiation levels on King George Island, from February to October, are about 3.588 and 36.8 KJ/m2 day from November to January. Thus, D. longifolius survives here with a solar radiation level as low as 3.588 KJ/m2 day.
A review of the aquatic mosses in Antarctica
Although Antarctica is commonly perceived as a land dominated by ice and snow, there are various types of wet and moist habitats, in which various types of wetland vegetation dominated by bryophytes has developed. Mosses occur in freshwater habitats ranging from mires, to streams, pools and lakes. Mosses growing permanently submerged in freshwater to slightly brackish lakes are a seldom documented but common feature of polar areas. They are especially abundant under oligotrophic conditions where little incoming radiation is absorbed by phytoplankton in the water column. Mosses are rare in the water with a depth shallower than 2 m, due to spring and summer ice-scour, but they have been recorded at depths greater than 30 m, and cover up to 40% of some lake bottoms. Phytoplankton are sparse in the clear lakes fed mainly by melt-water in Antarctica, and there are no emergent macrophytes. Benthic algae and mosses are the major primary producers in such lakes (Priddle 1980). Moss stems are invariably mixed with dense growths of algae and cyanobacteria.
Benthic moss was first reported from the Antarctic by Savicz-Lyubitskaya and Smirnova (1959) who described Bryum korotkevicziae L.I.Savicz et Smirnova dredged from depths of 33–36 m in Lake Figurnoye, Bunger Hills, Queen Mary Land. This species was subsequently redetermined as the common Antarctic terrestrial moss B. pseudotriquetrum (Hedw.) P. Gaertn., B. Mey. et Scherb., (Seppelt 1983, as B. algens Cardot). Since then, a number of aquatic mosses were reported from many lakes and water bodies in West and East Antarctica (Savicz-Lyubitskaya and Smirnova 1960, 1964; Light and Heywood 1973, 1975; Nakanishi 1977; Priddle and Dartnall 1978; Ochi 1979; Priddle 1979; Kaspar et al. 1982; Seppelt 1983; Pickard and Seppelt 1984; Kanda and Iwatsuki 1989; Kanda and Ohtani 1991; Imura et al. 1992; Kanda and Mochida 1992; Tewari and Pant 1996; Imura et al. 1999; Imura et al. 2003; Wagner and Seppelt 2006). They have often been identified only to genus or misidentified, but revision of most voucher collections has clarified their correct identities (Ochyra et al. 2008) (Table 2). Three of these mosses are now known to be exclusively aquatic in Antarctica, although they exist as terrestrial plants elsewhere in the world. These include Plagiothecium orthocarpum, which has its only known continental Antarctic records in several lakes in the Schirmachar Oasis, Dronning Maud Land, where it grows to 32.5 m depth (Savicz-Lyubitskaya and Smirnova 1964, as P. simonovii L.I.Savicz et Smirnova; Bednarek-Ochyra et al. 1999, as P. georgico-antarcticum). Pohlia wilsonii is another exclusively benthic moss reported from lakes near the Japanese Syowa station, Enderby Land, where it forms unique “towers” up to 60 cm high and 40 cm wide at depths of 3–5 m (Kanda and Mochida 1992; Imura et al. 1999, as Leptobryum pyriforme (Hedw.) Wilson). The third exclusively benthic moss in the Antarctic is D. longifolius, which is discussed in detail in the present account.
Twelve species of totally submerged mosses, belonging to 11 genera, are known in Antarctica. They have been found at many locations (Fig. 5). Of these, nine species have been recorded in West Antarctica, and four species in East Antarctica, with Bryum pseudotriquetrum being the only species known to occur in water bodies throughout the Antarctic. Five species (Cratoneuropsis relaxa (Hook.f. et Wilson) Broth. subsp. minor (Wilson et Hook.f.) Ochyra., Warnstorfia sarmentosa, Sanionia uncinata (Hedw.) Loeske, Drepanocladus longifolius and Pohlia nutans (Hedw.) Lindb.) have been found growing in lakes on Signy Island and four species are known from lakes on Alexander Island (Bryum pseudotriquetrum, Campylium polygamum, Dicranella campylophylla (Taylor) A.Jaeger and Distichium capillaceum (Hedw.) Bruch et Schimp.). In continental lakes the dominant moss is B. pseudotriquetrum which usually forms monospecific aggregations, but in the maritime Antarctic is sometimes accompanied by P. nutans, P. wilsonii and Hennediella heimii (Hedw.) R.H. Zander. Plagiothecium orthocarpum is the only verified benthic species in Schirmacher Oasis, and we have not been able to authenticate the record of P. wilsonii (Tewari and Pant 1996). B. pseudotriquetrum was found in Lake Vanda (77°32′S, 161°35′E), Victoria Land (Kaspar et al. 1982) which represents the southernmost distribution of an aquatic moss in this biome.

Distributions of aquatic mosses in Antarctic lakes. Star previous studies on aquatic mosses in Antarctic. Filled triangle this work
In contrast to mosses, benthic liverworts are generally less frequent and in the Antarctic only once has a hepatic been found submerged in a lake. This is Pachyglossa dissitifolia Herzog et Grolle which was collected with D. longifolius and Warnstorfia sarmentosa in Midge Lake on Livingston Island at about 1–2 m depth (Bednarek-Ochyra et al. 2000). A related species, Pachyglossa tenacifolia (Hook.f. and Taylor) Herzog et Grolle, has similarly been collected from relatively shallow waters at 1–2 m depth in Prion Lake, subantarctic Macquarie Island (Seppelt, unpublished collections).
Aquatic mosses have been present in continental Antarctic regions for thousands of years. Pickard and Seppelt (1984) discussed the presence of aquatic B. pseudotriquetrum (as B. algens) in the Vestfold Hills (68°30′S, 78°E) and dated subfossil deposits at between 7,000 and 8,000 years BP. These leafy stems were mixed with algae and cyanobacteria, as they do in present day living aquatic associations. The Vestfold Hills were completely overridden by the terminal Pleistocene advance of the Antarctic ice sheet and were subsequently exposed by its Holocene retreat (Adamson and Pickard 1983). The moss was presumed to have recolonized the region from elsewhere following the ice retreat. Ingólfsson et al. (1992) also reported a layer of subfossil of D. longifolius (as D. cf. aduncus) near Cape Lachman, James Ross Island, which is 9,525 years B·P. In the Larsemann Hills (69°30′S, 76°20′E), there is evidence of subfossil moss in lake sediments dating from at least 24,950 years BP (Burgess et al. 1994), providing an indication that at least some areas of present day coastal continental Antarctic regions may not have been fully overrun by ice during the Wisconsin period.
Changes in the growth and habit of benthic moss plants may provide evidence of changes in lake water conditions. As sensitive indicators of environmental conditions, mosses play an important role in the monitoring of environmental change in Antarctica (Rao 1982; Proctor 1984; Bates 1992). A detailed study of the geochemistry of lake waters, coupled with morphometrics of the benthic mosses, molecular systematic analysis of the relationships between aquatic and terrestrial forms of the same species seems warranted. Dunn and Robinson (2006) examined the effects of UV-B radiation on pigment composition in terrestrial forms of Ceratodon purpureus (Hedw.) Brid., B. pseudotriquetrum and Schistidium antarctici (Cardot) L.I. Savicz et Smirnova, from the Windmill Islands region (66°22′S, 110°30′E), concluding that B. pseudotriquetrum was best equipped to deal with the negative effects of increased exposure to UV radiation. Antarctic aquatic mosses may provide a useful surrogate measure for monitoring past changes in the levels of UV radiation through analysis of UV absorbing compounds coupled with a molecular genetic analysis of potential DNA change initiated by increased levels of UV radiation.
References
Adamson DA, Pickard J (1983) Late Quaternary ice movement across the Vestfold Hills, East Antarctica. In: Oliver RL, James PR, Jago JB (eds) Antarctic earth science. Cambridge University Press, Cambridge, pp 465–469
Bates JW (1992) Mineral nutrient acquisition and retention by bryophytes. J Bryol 17:223–240
Bednarek-Ochyra H, Lewis Smith RI, Ochyra R (1999) The genus Plagiothecium (Plagiotheciaceae, Bryopsida) in Antarctica. Bryobrothera 5:55–64
Bednarek-Ochyra H, Váňa J, Ochyra R et al (2000) The liverwort Flora of Antarctica. Institute of Botany, Polish Academy of Sciences, Cracow
Bian LG, Lu LH, Lu CG et al (1992) A study of radiative features at the Great Wall and Zhongshan Station of Antarctica. Q J Appl Metab 9:160–168 (in Chinese with English abstract)
Burgess JS, Spate AP, Shevlin J (1994) The onset of deglaciation in the Larsemann Hills, Eastern Antarctica. Antarct Sci 6:491–495. doi:10.1017/S095410209400074X
Chen FD, Li XD, Liu JX (1994) Handbook of Fildes Peninsula Antarctica mosses. Ocean Press, Beijing (in Chinese)
Dunn JL, Robinson SA (2006) UV-B screening potential is higher in two cosmopolitan moss species than in a co-occurring Antarctic endemic moss—implications of continuing ozone depletion. Gl Change B 12:2282–2296. doi:10.1111/j.1365-2486.2006.01283.x
Eights J (1833) Description of a new crustaceous animal found on the shores of the South Shetland, with remarks on their natural history. Trans Alb Ins 2:53–69 pls 1–2
Hedenäs L (1997) The Drepanocladus s str. species with excurrent costae (Amblystegiaceae). Nova Hedwig 64:535–547
Hedenäs L (2003) Amblystegiaceae (Musci). Flora Neotropica. Monograph 89. The New York Botanical Garden, New York
Imura S, Higuchi M, Kanda H et al (1992) Culture of rhizoidal tubers on an aquatic moss in the lakes near the Syowa Station area, Antarctica. Proc NIPR Symp Polar Biol 5:114–117
Imura S, Bando T, Saito S et al (1999) Benthic moss pillars in Antarctic lakes. Polar Biol 22:137–140. doi:10.1007/s003000050401
Imura S, Bando T, Seto K et al (2003) Distribution of aquatic mosses in the Sôya Coast region, East Antarctica. Polar Biosci 16:1–10
Ingólfsson Ó, Hjort C, Björck S et al (1992) Lake Pleistocene and Holocene history of James Ross Island, Antarctic Peninsula. Boreas 21:209–222
Kanda H, Iwatsuki Z (1989) Two aquatic mosses in the lakes near Syowa Station, Continental Antarctica. Hikobia 10:293–297
Kanda H, Mochida Y (1992) Aquatic mosses found in lakes of the Skarvsnes region, Syowa Station area, Antarctica (extended abstract). Proc NIPR Symp on Polar Biol 5:177–179
Kanda H, Ohtani S (1991) Morphology of the aquatic mosses collected in lake Yukidori, Langhovde, Antarctica (twelfth symposium on polar biology). Proc NIPR Symp Polar Biol 4:114–122
Kaspar M, Simmons GM Jr, Parker BC et al (1982) Bryum Hedw. collected from Lake Vanda, Antarctica. Bryologist 85:424–430. doi:10.2307/3242912
Kudoh S, Watanabe K, Imura S (2003a) Ecological studies of aquatic moss pillars in Antarctic lakes 2. Temperature and light environment at the moss habitat. Polar Biosci 16:23–32
Kudoh S, Kashino Y, Imura S (2003b) Ecological studies of aquatic moss pillars in Antarctic lakes 3. Light response and chilling and heat sensitivity of photosynthesis. Polar Biosci 16:33–42
Lewis AR, Marchant DR, Ashworth AC et al (2008) Mid-Miocene cooling and the extinction of tundra in continental Antarctica. Proc Natl Acad Sci USA 105:10676–10680. doi:10.1073/pnas.0802501105
Light JJ, Heywood RB (1973) Deep-water mosses in Antarctic lakes. Nature 242:535–536. doi:10.1038/242535a0
Light JJ, Heywood RB (1975) Is the vegetation of continental Antarctica predominantly aquatic? Nature 256:199–200. doi:10.1038/256199a0
Nakanishi S (1977) Ecological studies of the moss and lichen communities in the ice-free areas near Syowa Station, Antarctica. Antarct Rec 59:68–96
Ochi H (1979) A taxonomic review of the genus Bryum, musci in Antarctica. Mem Natl Inst Polar Res 11:70–80
Ochyra R (1998). The moss flora of King George Island, Antarctica. Polish Academy of Sciences, W. Szafer Institute of Botany, Cracow
Ochyra R, Matteri CM (2001) Amblystegiaceae. Flora Criptogámica de Tierra del Fuego. In: Guerrera SA, Gamundi de Amos IJ, Matteri CM (eds) Buenos Aires: Consejo Nacional de Investigaciones Clientíficasy Técnicas de la Republica Argentina, pp 1–96
Ochyra R, Bednarek-Ochyra H, Lewis Smith RI (2002) New and rare moss species from subantarctic South Georgia. Nova Hedwig 74:121–147. doi:10.1127/0029-5035/2002/0074-0121
Ochyra R, Lewis Smith RI, Bednarek-Ochyra H (2008) The illustrated moss flora of Antarctica. Cambridge University Press, Cambridge
Pickard J, Seppelt RD (1984) Holocene occurrence of the moss Bryum algens Card in the Vestfold Hills, Antarctica. J Bryol 13:209–217
Priddle J (1979) Morphology and adaptation of aquatic mosses in an Antarctic lake. J Bryol 10:517–529
Priddle J (1980) The production ecology of benthic plants in some Antarctic lakes: I. In situ production studies. J Ecol 68:141–153. doi:10.2307/2259248
Priddle J, Dartnall HJG (1978) The biology of an Antarctic aquatic moss community. Freshw Biol 8:469–480. doi:10.1111/j.1365-2427.1978.tb01469.x
Proctor MCF (1984) Structure and ecological adaptation. In: Dyer AFD, Duckett JG (eds) The experimental biology of bryophytes. Academic Press, London, pp 9–37
Rao DN (1982) Responses of bryophytes to air pollution. In: Smith AJE (ed) Bryophyte ecology. Chapman and Hall, London, pp 445–471
Riis T, Sand-Jensen K (1997) Growth reconstruction and photosynthesis of aquatic mosses: influence of light, temperature and carbon dioxide at depth. J Ecol 85:359–372. doi:10.2307/2960508
Savicz-Lyubitskaya LI, Smirnova ZN (1959) A new species of the genus Bryum Hedw. from Bunger’s Oasis. Inf Byull Sov Antarkt Eksped 7:34–39 (in Russian)
Savicz-Lyubitskaya LI, Smirnova ZN (1960) New variety of Bryum korotkevicziae Sav.-Ljub. et Z. Smirn. Inf Byull Sov Antarkt Eksped 17:25–27 (in Russian)
Savicz-Lyubitskaya LI, Smirnova ZN (1964) A deep-water member of the genus Plagiothecium Br. et Sch. in Antarctica. Inf Byull Sov Antarkt Eksped 49:33–39 (in Russian)
Seppelt RD (1983) The status of the Antarctic moss Bryum korotkevicziae. Lindbergia 9:21–26
Seppelt RD, Selkirk PM (1984) Effects of submersion on morphology and the implications of induced environmental modification on the taxonomic interpretation of selected Antarctic moss species. J Hattori B 55:273–279
Takaki N (1985) The longest moss. Bryol Times 30:4
Tewari SD, Pant G (1996) Some moss collections from Dakshin Gangotri, Antarctica. Bryol Times 91:7
Wagner B, Seppelt R (2006) Deep-water occurrence of the moss Bryum pseudotriquetrum in Radok Lake, Amery Oasis, East Antarctica. Polar Biol 29:791–795. doi:10.1007/s00300-006-0116-7
Acknowledgments
We wish to express our sincere gratitude to Chinese Arctic and Antarctic Administration, the State Oceanic Administration of China. Also, many thanks to the members of the 23rd Chinese National Antarctic Research Expedition teams for their help during the field work. This research was supported by the Strategic Research Foundation on Polar Sciences of China (No. 20080220).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Li, SP., Ochyra, R., Wu, PC. et al. Drepanocladus longifolius (Amblystegiaceae), an addition to the moss flora of King George Island, South Shetland Islands, with a review of Antarctic benthic mosses. Polar Biol 32, 1415–1425 (2009). https://doi.org/10.1007/s00300-009-0636-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-009-0636-z