
Abstract
This paper analyzes the spatiotemporal variation and the causes of seabird bycatch by the Uruguayan pelagic longline fleet in a region of the Atlantic Ocean where the world’s highest historical rates of seabird bycatch were recorded. The study is based on data obtained by the Uruguayan Observers Program in 29 trips, conducted from 1998 to 2004, totalling about 648,000 hooks. The bird capture per unit of effort (BCPUE) for the studied period was 0.42 birds/1,000 hooks. The highest BCPUE values were recorded in the period May–November. Three zones were identified, with BCPUEs of 2.50 birds/1,000 hooks (very high); 0.78 birds/1,000 hooks (high) and 0.04 birds/1,000 hooks (low). Though these BCPUE values are lower than those historically reported, some are still high in global terms. Night setting was found to be effective in reducing seabird bycatch, but it is necessary to implement additional measures as seabirds have access to bait also by night, especially during the more luminous moon phases.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Incidental capture in various fisheries is one of the main threats to the conservation of marine biodiversity and affects mammals, turtles, sharks, and birds (Brothers et al. 1999a; Oravetz 1999; Hall et al. 2000; Lewison et al. 2004; Carranza et al. 2006; Domingo et al. 2006a). At present, industrial longline fishing in areas of high biological productivity poses a big threat to seabirds occurring in those areas (Alexander et al. 1997; Brothers et al. 1999a). Many seabird species are attracted by baits deployed during line setting and waste from processed fish discarded into the sea. When seabirds attempt to feed from baited hooks, especially during setting operations, they often become entangled in the lines or hooked, and many drown. This results in the capture and death of tens of thousands of birds per year (Brothers 1991; Alexander et al. 1997; Gales et al. 1998; Brothers et al. 1999a). The most captured species throughout the world are albatrosses (Diomedeidae) and petrels of the Procellariidae family (Alexander et al. 1997; Brothers et al. 1999a).
Several pelagic longline fleets operate in the South Atlantic (Hazin et al. 2008; Domingo et al. 2006a) where they represent a large proportion of the fishing effort in the area (Tuck et al. 2003; Domingo et al. 2006b). This region is frequented by large numbers of seabirds of several species from breeding sites in the Southern Ocean, such as the Falkland/Malvinas, South Georgia and the sub Antarctic Islands of New Zealand (Tickell 1967; Escalante 1970; Prince et al. 1992; Robertson et al. 2003). Many of these species are globally threatened according to the IUCN (BirdLife International 2008). Species with a threatened conservation status that migrate from the south to Uruguayan waters and from breeding islands to the east include Black-browed albatross (Thalassarche melanophrys), Wandering albatross (Diomedea exulans), Southern Royal albatross (D. epomophora), Northern Royal albatross (D. sanfordi), White-chinned petrel (Procellaria aequinoctialis), Tristan albatross (D. dabbenena), and Atlantic yellow-nosed albatross (T. chlororhynchos).
An increasing amount of information on seabird bycatch in longline fishing in the Southwest Atlantic has become available in recent years (Vaske 1991; Neves and Olmos 1998; Schiavini et al. 1998; Stagi et al. 1998; Favero et al. 2003; Gandini and Frere 2006; Gómez-Laich et al. 2006; Seco Pon et al. 2007). However, data for pelagic longline fisheries, especially for those operating off Uruguay and adjacent waters are still scarce. Earlier papers (Vaske 1991; Alexander et al. 1997; Stagi et al. 1998) reported captures of 3.82, 4.7 and 5.03 birds/1,000 hooks in the area, the highest values reported at world level (Alexander et al. 1997). This region should therefore be taken into consideration in any effort to reduce seabird mortality in the southern hemisphere.
From 1997 to the present, no studies on bycatch with pelagic longline in this area were conducted. This paper attempts to fill this information gap. We analyse the spatial and temporal variation in seabird bycatch recorded on 29 commercial fishing trips between 1998 and 2004 by Uruguay’s National Observers Program on Board the Tuna Fleet (Programa Nacional de Observadores a bordo de la Flota Atunera Uruguaya—“PNOFA”) in a wide area of the Southwest Atlantic. We also analyze several environmental factors to determine their influence on the rate of seabird bycatch.
Materials and methods
Description of the fishery
The Uruguayan pelagic longline fleet has been operating uninterruptedly since 1981. Until 1991 there was a marked Asian participation in the fleet and the preferred gear was the Japanese type longline The main target species at that time were swordfish (Xiphias gladius) and bigeye tuna (Thunnus obesus) and albacore (T. alalunga) (Domingo et al. 2002). Since 1991 the main target species have been pelagic sharks, swordfish, and tuna (Domingo et al. 2002; Mora and Domingo 2006). The most commonly used fishing gear is the American-type monofilament longline (Domingo et al. 2002), but some freezer longliners have employed Spanish- type multifilament longline. Figure 1 shows a schematic representation of both types of gear.

Schematic representation of both types of gear used by the Uruguayan pelagic longline fishery. a American-type monofilament longline. Gangion lines have a section indicated with letter a, that usually consists of a polyamide monofilament (3.5–5.5 m) for swordfish and of a shorter steel or plaited monofilament for sharks. b Spanish-type multifilament longline. Each gangion line has three sections: a made of multifilament (0.9 m) fixed to the mother line; b made of multifilament (1.80 m) attached to the first by a snap; and c made of three twisted wire threads (5 m)
Typically, fishing operations are carried out as follows: The longline is set over the vessel’s stern, most times after sunset. The setting is generally completed before midnight. Hooks are set at 9–12-s intervals in the American longline and every 32 m in the Spanish longline. Early in the morning the gear is heaved at one side of the vessel. Heaving takes approximately 7 h, but this varies according to the number of hooks set, the volume of the capture and the sea conditions. The baits most commonly used are squid (Illex argentinus) and mackerel (Scomber spp., Trachurus spp.) between 20 and 35 cm in length, thawed a few hours before line setting (Domingo et al. 2002; Mora and Domingo 2006).
Data collection
From April 1998 to January 2004 we analysed data from 29 trips comprising a total of 450 sets and 647,722 hooks (Table 1). The main tasks of the observers were to record operational variables, record the catch, classify the capture, catch, discard (bycatch and released), and lost catch, along with the recording of biological information. With regard to seabirds, the work of observers was limited to recording the captures—which were classified as discard—and identifying specimens to the lowest possible taxonomic level. However, as some of the observers had little experience with seabirds, many specimens could not be identified. In addition, the species identification of great albatrosses (Diomedea spp.) by observers was not taken into consideration, given the difficulties in their classification. Species identification was made by using field guides (Harrison 1985; Vooren and Fernandes 1989; Narosky and Zurieta 1993; Onley and Bartle 1999). Bibliography (Cuthbert et al. 2003), photographs, skulls and bills collected by the observers were also used. Recapture of ringed birds was reported and was also used to confirm the species.
Settings were identified by date, position and starting time. The fishing effort was measured by the number of hooks. The sea surface temperature (SST) was measured with vessels’ instruments at a depth of 2–3 m.
Data analysis
The bird capture per unit of effort (BCPUE) was defined as the number of birds captured per thousand of hooks set. The spatial distribution of the fishing effort and bird catch was based on the position of the vessel at the starting of each set.
To analyze the spatial variation of the BCPUE we used a Classification and Regression Tree Analysis (CART) (Breiman et al. 1984). The latitude and longitude at the beginning of each set were used as continuous independent variables. CART was carried out on software R 2.6.1. (R Development Core Team 2007) using the tree package (Ripley 2007). BCPUE per area was defined as Very High (V), High (H) and Low (L), based on the values provided by Alexander et al. (1997) from 0.02 to 1.09 birds/1,000 hooks, with an estimate global average of 0.4 birds/1,000 hooks.
The temporal variation in seabird bycatch within one year was based on the average monthly BCPUE for the study period (with the exception of February). Temporal comparisons were made with the Mann-Whitney (M-W) test (Zar 1999).
The correlation between BCPUE and SST was estimated by means of the Spearman coefficient (R) (Zar 1999), using the average SST at the beginning and at the end of each set. In this analysis only sets with capture were considered. The effect of day setting on seabird bycatch was compared to the effect of night setting by means of the M-W test using the initial setting time and the BCPUE per set. All sets that started before sunset were considered as day sets. The rest were marked as night sets. To determine the relationship between BCPUE and moon phases, night sets were classified according to the moon phase under which they were conducted: first quarter, full moon, last quarter, and new moon. Kruskall–Wallis (KW) tests (Zar 1999) were performed to determine the difference in BCPUE among moon phases. Paired comparisons with the M-W test were also made.
Results
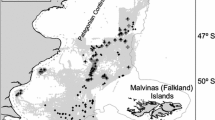
From April 1998 to January 2004 the monitored vessels fished in Uruguayan jurisdictional waters on the continental shelf and slope and in adjacent international waters extending to the northeast up to 26ºS and 21ºW (Fig. 2). The effort per set ranged from 200 to 3,360 hooks (median = 1,000 hooks).

Distribution of the observed fishing effort during the study period (fishing effort is represented in number of hooks per set)
In total, 272 seabirds of at least seven species were captured in the 6-year period covered by this study. These included Wandering, Black-browed, Yellow-nosed, and Tristan albatrosses; White-chinned and Cape (Daption capense) petrels and Great shearwater (Puffinus gravis). A total of 128 out of the 272 specimens were identified to species level. Black-browed albatrosses were the most numerous (67%, n = 86), followed by White-chinned petrels (16%, n = 20). Great albatrosses (Diomedea spp.) represented 13% of the seabird bycatch (n = 17) and included one specimen of Tristan albatross and three Wandering albatrosses. Single specimens of the Great shearwater and of Cape petrel were recorded (Table 2).
The BCPUE in the Uruguayan pelagic longline fleet from 1998 to 2004 was 0.42. Birds were captured in 21.3% (n = 96) of the sets studied (Fig. 3).

Frequency distribution of the BCPUE (birds/1,000 hooks) calculated for each observed set (n = 450) of the 29 monitored trips for the period 1998–2004
Spatial and temporal variation
Bird bycatch was not homogeneous within the study area. Three different zones were identified, longitude being the split criterion (Fig. 4). Zone V comprised the external shelf and the Uruguayan slope (34º30′–37ºS and 51º30′–53ºW). The BCPUE averaged 2.50 birds/1,000 hooks (Fig. 5a). Within the zone, the highest BCPUE values (25 birds/1,000 hooks) were recorded on the continental slope with isobaths of 200–2,000 m (Fig. 5b). Zone H comprised oceanic Uruguayan waters, international waters adjacent to Uruguayan and South Brazilian waters (32º30′–37ºS and 47º30′–51º30′W). Here the mean BCPUE was 0.78 (Fig. 5a). In zone L, located in international waters, East of 47º30′W (Fig. 5a), the mean BCPUE was 0.04 birds/1,000 hooks.

Results of the regression analysis utilized to differentiate zones of incidental capture. In each terminal node the mean BCPUE (birds/1,000 hooks) is shown for the identified zones and the number of data. In each node the criteria for the decision are shown. Data with values of less than the splitting point go to the left daughter node. The (−) sign in the longitude values within the nodes correspond to western coordinates and are in a decimal scale [−51.5 = 51º30′W, −47.5 = 47º30′W]

a Spatial variation of the BCPUE (birds/1,000 hooks) in the three identified zones (V = very high BCPUE, H = high BCPUE and L = low BCPUE). b Distribution of the BCPUE in the zone V
Seabird bycatch varied considerably between months with average BCPUE values ranging from 0 to 1.80 birds/1,000 hooks (Table 3). In December no bird captures were recorded. In summer and early fall, the months of December–April, the average BCPUE was 0.09 birds/1,000 hooks (SE ± 0.05) with seven seabirds captured. In late fall, winter and late spring, the months of May–November, the BCPUE was 0.98 birds/1,000 hooks (SE ± 0.16) which was significantly higher (M-W: U 1, 450 = 14,679.0, P < 0.01) than the summer-early fall season. In fact, 97.4% of the bird captures and the highest monthly averages occurred from May to November. The highest mean monthly BCPUE occurred in September.
Factors affecting mortality
Considering the marked variation in seabird bycatch rates among zones V, H and L, we analysed independently for each zone the effect of environmental (SST, moon phase) and operational variability on BCPUE. In zone V, where the highest BCPUE was recorded for the period May–November, a negative correlation was observed between SST and BCPUE (R = −0.59; P < 0.01; n = 33). For this zone and period, BCPUE tended to be higher in day sets; however, the difference with night sets was not significant (M-W: U 1,83 = 671.5; P = 0.35) (Fig. 6). On the other hand, significant variability was observed with changes in the moon phase (K-W: H 3,55 = 7.995; P < 0.05), with higher mortality rates observed under full and first quarter moon (Fig. 7).

BCPUE (birds/1,000 hooks) for day (D) and night (N) sets in the three zones (V = very high BCPUE, H = high BCPUE and L = low BCPUE) for the period May–November

BCPUE (birds/1,000 hooks) according to the moon phase for night sets in the three zones (V, H and L as in Fig. 6) for the period May–November. Significant differences (paired Mann-Whitney comparisons P < 0.05) are shown with asterisk
In zone H the BCPUE showed a significant relation with the setting time in the period May–November. Higher BCPUE values were observed in day sets (M-W: U 1,132 = 1,335; P < 0.05; Fig. 6). Similar to zone V, BCPUE varied according to the moon phase (K-W: H 3,94 = 10.02; P < 0.05), with higher values under full and first quarter moon (Fig. 7).
In zone (L) the statistical tests disclosed no relations between seabird bycatch and the environmental factors for the May–November period. No relation between BCPUE and the studied factors was observed in the three zones for the period December–April.
Discussion
Bycatch rates
The overall BCPUE of 0.42 birds/1,000 hooks recorded in the observed Uruguayan pelagic longline fleet for the period 1998–2004 was similar to the BCPUE (0.4 birds/1,000 hooks) reported for other longline fisheries that operate in the Southern Hemisphere in the 1990s (Brothers 1991; Alexander et al. 1997; Barnes et al. 1997). The first studies on seabird bycatch in the pelagic longline fisheries in the Southwest Atlantic showed that these fisheries were an important cause of seabird mortality (Vaske 1991; Stagi et al. 1998). The bird bycatch rates observed for Uruguayan and Southern Brazilian waters 4.7–5.3 birds/1,000 hooks and 3.82 birds/1,000 hooks, respectively, were exceptionally high (Alexander et al. 1997; Barnes et al. 1997). These values were obtained mainly on the continental slope off Uruguay and South of Brazil (Vaske 1991; Stagi et al. 1998). Our study revealed a significant spatial (zones V, H and L, Fig. 5a) and temporal (highest BCPUE in May–November) variability in seabird bycatch. The highest BCPUE was recorded on the Uruguayan continental slope (zone V), with a mean BCPUE of 2.50. Though this value is lower than those reported in earlier studies, it is still very high at world level.
Oceanographic factors affecting mortality
In Brazil, with a well developed commercial pelagic longline fishery, the highest bird captures have been recorded towards the south (Vaske 1991; Neves and Olmos 1998), mainly on the continental slope. Similarly, in a systematic study with bottom longline conducted on the continental shelf and slope south and southeast off Brazil (22º–34º40′S), it was observed that bird capture was concentrated in 30º–35ºS, adjacent to Uruguayan waters (Vooren and Coelho 2004). Currently, Argentine does not have a commercial pelagic longline fleet. However, it is known that bycatch in the commercial bottom longline fishery is higher on the slope, with an estimated bird capture of thousands of birds a year (Favero et al. 2003).
One of the most important oceanographic features in these waters under study is the sub-tropical convergence that is the confluence of the Brazil subtropical current and the Falklands current (Seeliger et al. 1998). In general, the region of influence of the sub-tropical convergence has been highlighted as an area of great richness, abundance and biomass of seabirds (Rumboll and Jehl 1977; Veit 1995; Vooren and Coelho 2004; Favero and Silva Rodriguez 2005). Within this area, the continental slope is considered one of the main frontal zones of South America (Acha et al. 2004). The subtropical shelf front (a subsurface thermohaline front), formed by an extension of the convergence, reaches the slope in Uruguayan waters (Piola et al. 2000), while the Argentine shelfbreak front (a thermohaline front) formed by sub Antarctic waters of the Argentinean shelf and waters of the Falkland current, extends along the continental slope into Uruguayan waters (Acha et al. 2004). This highly productive area may provide an important feeding habitat for seabirds (mainly albatrosses and petrels) (Thurston 1982; Favero and Silva Rodriguez 2005). As various fisheries also concentrate in this area, the interaction between longline fisheries that operate in the region, particularly near the slope, and seabirds is very frequent and may be the cause of increased mortality rates (Vaske 1991; Stagi et al. 1998).
Other factors affecting mortality
The bird capture rates varied from month to month with more than 97% of the BCPUE occurring between May and November. This is very similar to the information reported for the Brazilian tuna fleet where 94% of the seabird bycatch occurred between May and October (Neves and Olmos 1998). This period (mid-fall to mid-spring) is characterized by the lowest SST and the prevalence of the Falkland current. The highest BCPUE values per set were recorded in the coldest waters. It is also the time of the year when migrating albatrosses and petrels from the South, especially the most commonly captured species such as Black-browed albatross and White-chinned petrel, are most abundant (Escalante 1970; Vooren and Fernandez 1989; Olmos 2002).
Most birds that interact with longline fisheries are “visual” predators that feed by day (Vaske 1991; Weimerskirch and Wilson 1992; Brothers et al. 1999a), though there are those that also feed at night (e.g., Procellaria spp.) (Weimerskirch et al. 1999). The highest BCPUE values were recorded in day sets, as observed in other studies (Gales et al. 1998; Brothers et al. 1999b; Belda and Sanchez 2001; Sullivan et al. 2004). Higher BCPUE values in night sets with first quarter and full moon appear to confirm that the intensity of light is one of the main factors that affect seabird bycatch. However, Goméz-Laich and Favero (2007) found no relation between the moon phase and captures of White-chinned petrels. Though night setting diminishes the capture of birds and is one of the mitigation measures in practice in various longline fleets, it should be taken into account that its effectiveness is lower during the moon’s more luminous phases.
Implications for seabird conservation in the Southern Ocean
Specimens of at least seven species were captured, five of which are globally threatened. The most captured species was the Black-browed albatross, which is the most abundant albatross in Uruguayan waters (Escalante 1970; Stagi et al. 1998). Other species include the white-chinned petrel and albatrosses of the Diomedea genus. Black-browed albatrosses and White-chinned petrels are the most affected by longline fisheries at a regional level (Vaske 1991; Barnes et al. 1997; Neves and Olmos 1998; Stagi et al. 1998; Brothers et al. 1999a; Gómez Laich et al. 2006; Sullivan et al. 2004) and also in Uruguay. Stagi et al. (1998) reported a high proportion of Black-browed albatrosses captured in nine pelagic longline fishing trips in Uruguayan waters between 1993 and 1994. Vaske (1991) reported a majority of White-chinned petrels captured in 52 fishing days with pelagic longline between 1987 and 1990 off Southern Brazil. The largest populations of Black-browed albatrosses and White-chinned petrels, located in the Falkland/Malvinas and South Georgia, respectively (Coxall and Gales 1998; Berrow et al. 2000a), have declined (Berrow et al. 2000a; Huin 2001; Sullivan et al. 2004) and longline fishing appears to be the main cause. It is very likely that the Black-browed albatrosses captured during the period of our study breed at the Falkland/Malvinas Islands, but with a small proportion of wintering birds from South Georgia (Phillips et al. 2005). In contrast, most of the White-chinned petrels are likely to be wintering birds originating from the South Georgia population, and only a tiny proportion from the small breeding population in the Falkland/Malvinas (Phillips et al. 2006; Reid et al. 2007).
Regarding the provenance of Wandering albatrosses, various studies have shown that the specimens of this species come from Bird Island, South Georgia (Vooren and Fernandes 1989; Croxall and Prince 1990; Prince et al. 1992; Stagi et al. 1998; Olmos 2002). The decline of this population over the last decades (Croxall 1979; Croxall et al. 1998; Poncet et al. 2006) has been largely ascribed to longline fishing. Croxall and Prince (1990) showed that in the period 1960–1974, 34% of Wandering albatross’ rings from Bird Island were reported from South America, and the proportion grew up to 65% for the period 1975–1988. Most reports from this last period corresponded to birds captured in fisheries (mainly longline) in Uruguayan and Southern Brazilian waters. The increasing number of reports was in line with the development and expansion of the longline fishery in this area and with the decline in the above-mentioned populations (Croxall and Prince 1990). The strongest impact of longline fisheries on seabirds in the region has been recorded on the continental slope. The area comprises Argentinean, Uruguayan and Brazilian waters where only the respective fleets are allowed to operate. Several foreign longline fleets also operate or have historically operated in adjacent international waters, where we recorded a high BCPUE (0.78 in zone H). A detailed description of the effort of the principal fleets was provided by Tuck et al. (2003). The Atlantic Ocean area adjacent to Uruguayan waters has been one of the places where the effort of the various fleets has concentrated over the last decades (Hazin et al. 2008). This high concentration of longline vessels in the area could be causing significant impact on seabirds, but no data are publicly available on bycatch by these foreign fleets. In spite of this, published information (e.g., Vaske 1991; Neves and Olmos 1998; Favero et al. 2003) plus the data provided in this study suggest that the incidental mortality of some species mentioned above could be the main cause of their population declines. In the 29 trips analyzed in this study no mitigation measures were used though night setting for operational reasons had a mitigating effect. Our results confirm that seabird mortality in the region can be partially avoided with night setting. Additional measures should be adopted on an urgent basis as albatrosses and petrels have also access to bait at night, especially during the more luminous moon phases. It is necessary to continue monitoring seabird bycatch to assess capture trends, especially in Uruguayan and adjacent international waters. Monitoring this area is of paramount importance for the adoption of national and international conservation measures and for the management of the longline fishery.
References
Acha EM, Mianzan HW, Guerrero RA, Favero M, Bava J (2004) Marine fronts at the continental shelves of austral South America: Physical and ecological processes. J Mar Syst 44:83–105
Alexander K, Robertson G, Gales R (1997) The incidental mortality of Albatrosses in longline fisheries. Australian Antarctic Division, Tasmania
Barnes KN, Ryan PG, Boix-Hinzen C (1997) The impact of the hake Merluccius spp. Longline fishery off South Africa on Procellariiform seabirds. Biol Conserv 82:227–234
Belda EJ, Sánchez A (2001) Seabirds mortality on longline fisheries in the western Mediterranean: factors affecting bycatch and proposed mitigating measures. Bio Conserv 98:357–363
Berrow SD, Croxall JP, Grant SD (2000) Status of white-chinned petrels Procellaria aequinoctialis Linnaeus 1758, at Bird Island, South Georgia. Antarct Sci 12:399–405
BirdLife International (2008) Species factsheet. Available at: http://www.birdlife.org. Accessed 26 Jul 2008
Breiman L, Friedman JH, Olshen RA, Stone CJ (1984) Classification and regression trees. Chapman and Hall, New York
Brothers N (1991) Albatross mortality and associated bait loss in the Japanese fishery in the southern Ocean. Biol Conserv 55:255–268
Brothers NP, Cooper J, Løkkeborg S (1999a) The incidental catch of seabirds by longline fisheries: worldwide review and technical guidelines for mitigation. FAO Fisheries Circular No. 937, Food and Agriculture Organization of the United Nations, Rome
Brothers NP, Gales R, Reid T (1999b) The influence of environmental variables and mitigation measures on seabird catch rates in the Japanese tuna longline fishery within the Australian Fishing Zone, 1991–1995. Biol Conserv 88:85–101
Carranza A, Domingo A, Estrades A (2006) Pelagic longlines: a threat to sea turtles in the Ecuatorial Eastern Atlantic. Biol Conserv 131:52–57
Croxall JP (1979) Distribution and population changes in the Wandering Albatross at South Georgia. Ardea 67:15–21
Croxall JP, Prince PA (1990) Recoveries of Wandering Albatross Diomedea exulans ringed at South Georgia 1958–1986. Ringing Migr 11:43–51
Croxall JP, Prince PA, Rothery P, Wood AG (1998) Populations changes in albatrosses at South Georgia. In: Robertson G, Gales R (eds) Albatross biology and conservation. Surrey Beatty, Chipping Norton, pp 69–83
Cuthbert R, Phillips RA, Ryan PG (2003) Separating the Tristan Albatross and the Wandering Albatross using morphometric measurements. Waterbirds 26:338–344
Domingo A, Mora O, Cornes M (2002) Evolución de las capturas de elasmobranquios pelágicos en la pesquería de atunes de Uruguay, con énfasis en los tiburones Azul (Prionace glauca), Moro (Isurus oxyrinchus) y Porbeagle (Lamna nasus). Col Vol Sci Pap ICCAT 54:1406–1420
Domingo A, Bugoni L, Prosdocimi L, Miller P, Laporta M, Monteiro DS, Estrades A, Alvareda D (2006a) El impacto generado por las pesquerías en las tortugas marinas en el Océano Atlántico sud occidental. WWF Programa Marino para Latino América y el caribe, San José, Costa Rica
Domingo A, Sales G, Giffoni B, Miller F, Laporta M, Maurutto G (2006b) Captura incidental de tortugas marinas con palangre pelágico en el Atlántico Sur por las flotas de Brasil y Uruguay. Col Vol Sci Pap ICCAT 59:992–1002
Escalante R (1970) Aves marinas del Río de la Plata y aguas vecinas de Océano Atlántico. Barreiro y Ramos S. A, Montevideo
Favero M, Silva Rodríguez MP (2005) Estado actual y conservación de aves pelágicas que utilizan la plataforma continental argentina como área de alimentación. Hornero 20:95–110
Favero M, Khatchikian CE, Arias A, Silva Rodríguez MP, Cañete G, Mariano-Jelicich R (2003) Estimates of seabirds by-catch along the Patagonian shelf by Argentine longline fishing vessels, 1999–2001. Bird Conserv Int 13:273–281
Gales R, Brothers N, Reid T (1998) Seabirds mortality in the Japanese tuna longline fishery around Australia, 1988–1995. Biol Conserv 86:37–56
Gandini PA, Frere E (2006) Spatial and temporal patterns in the bycatch of seabirds in the Argentinean longline fishery. Fish Bull 104:482–485
Gómez-Laich A, Favero M (2007) Spatio-temporal variation in mortality rates of White-chinned Petrels Procellaria aequinoctialis interacting with longliners in the south-west Atlantic. Bird Conserv Int 17:359–366
Gómez-Laich A, Favero M, Mariano-Jelicich R, Blanco G, Cañete G, Arias A, Silva Rodriguez P, Brachetta H (2006) Environmental and operational variability affecting the mortality of black-browed Albatrosses associated with long-liners in Argentina. Emu 106:21–28
Hall M, Alverson DL, Metuzals KI (2000) By-Catch: problems and Solutions. Mar Pollut Bull 41:204–219
Harrison P (1985) Seabirds, an identification guide. Houghston Mifflin Company, Boston
Hazin FHV, Broadhurst MK, Amorim AF, Arfelli CA, Domingo A (2008) Catches of pelagic sharks by subsurface longline fisheries in the South Atlantic Ocean during the last century: a review of available data with emphasis on Uruguay and Brazil. In: Camhi M, Pikitch E, Babcock E (eds) Sharks of the Open Ocean. Blackwell, New York
Huin N (2001) Census of the Black-browed albatross population of the Falkland Islands. Falklands Conservation, Falkland Islands
Lewison RL, Crowder LB, Read AJ, Freeman SA (2004) Understanding impacts of fisheries bycatch on marine megafauna. Trends Ecol Evol 19:598–604
Mora O, Domingo A (2006) La flota atunera uruguaya: evolución y tendencias (1981–2004). Col Vol Sci Pap ICCAT 59:608–614
Narosky T, Yzurieta D (1993) Guía para la identificación de las aves de Argentina y Uruguay. Asociación Ornitológica del Plata, Buenos Aires
Neves T, Olmos F (1998) Albatross mortality in fisheries off the coast of Brazil. In: Robertson G, Gales R (eds) Albatross biology and conservation. Surrey Beatty, Chipping Norton, pp 214–219
Olmos F (2002) Non-breeding seabirds in Brazil: a review of bands recoveries. Ararajuba 10:31–42
Onley D, Bartle S (1999) Identificación de aves marinas de los océanos del sur. Una guía para observadores científicos a bordo de buques pesqueros. Te Papa Press, Wellington, New Zealand
Oravetz CA (1999) Reducción de la Captura Incidental en Pesquerías. In: Eckert KL, Bjorndal KA, Abreu-Grobois FA, Donnelly M (eds) Research and management techniques for the conservation of sea turtles. IUCN/SSC Marine Turtle Specialist Group Publication No 4, pp 217–222
Phillips RA, Silk JRD, Croxall JP, Afanasyev V, Benett VJ (2005) Summer distribution and migration of nonbreeding albatrosses: individual consistencies and implications for conservation. Ecology 81:2386–2396
Phillips RA, Silk JRD, Croxall JP, Afanasyev V (2006) Year-round distribution of white-chinned petrels from South Georgia: Relationships with oceanography and fisheries. Biol Conserv 129:336–347
Piola AR, Campos EJD, Moller OO Jr, Charo M, Martinez C (2000) Subtropical Shelf Front off eastern South America. J Geophys Res 105:6565–6578
Prince PA, Wood AG, Barton T, Croxall JP (1992) Satellite tracking of wandering albatrosses (Diomedea exulans) in the South Atlantic. Antarct Sci 4:31–36
Poncet S, Robertson G, Phillips RA, Lawton K, Phalan B, Trathan PN, Croxall JP (2006) Status and distribution of wandering, black-browed and grey-headed albatrosses breeding at South Georgia. Polar Biol 29:772–781
R Development Core Team (2007) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. Available at: http://www.r-project.org/. Accessed 22 Sep 2008
Reid TA, Lecoq M, Catry P (2007) The White-chinned Petrel Procellaria aequinoctialis population of the Falkland Island. Mar Ornithol 35:57–60
Ripley B (2007) Tree: classification and regression trees. [R package version 1.0-26]. Available at: http://cran.r-project.org/web/packages/tree/. Accessed 22 Sep 2008
Robertson CJR, Bell EA, Sinclair N, Bell BD (2003) Distribution of seabirds from New Zealand that overlap with fisheries worldwide. DOC Science for Conservation 233, Departament of Conservation, Wellington, New Zealand
Rumboll MAE, Jehl JR Jr (1977) Observations on pelagic birds in the South Atlantic Ocean in the Austral Spring. San Diego Soc Nat Hist Trans 19:1–16
Schiavini A, Frere E, Gandini P, García N, Crespo E (1998) Albatross-fisheries interactions in Patagonian shelf waters. In: Robertson G, Gales R (eds) Albatross biology and conservation. Surrey Beatty, Chipping Norton, pp 208–213
Seco Pon JP, Gandini PA, Favero M (2007) Effect of longline configuration on seabird mortality in the Argentine semi-pelagic Kingclip Genypterus blacodes fishery. Fish Res 85:101–105
Seeliger U, Odebrecht C, Castello JP (1998) Os ecossitemas costeiro e marinho do extremo Sul do Brasil. Ecoscientia, Rio Grande
Stagi A, Vaz-Ferreira R, Marin Y, Joseph L (1998) The conservation of albatrosses in Uruguayan waters. In: Robertson G, Gales R (eds) Albatross biology and conservation. Surrey Beatty, Chipping Norton, pp 220–224
Sullivan BJ, Reid TA, Pompert J, Enticott JW, Black AD (2004) Seabird mortality associated with Patagonian Toothfish (Dissostichus eleginoides) longliners in Falkland Island waters. Emu 104:317–325
Tickell WLN (1967) Movements of Black-browed and Grey-headed Albatrosses in the South Atlantic. Emu 66:357–367
Thurston MH (1982) Ornithological observations in the South Atlantic Ocean and Weddell Sea, 1959–64. Br Antarct Survey Bull 55:77–103
Tuck GN, Polacheck T, Bulman CM (2003) Spatio-temporal trends of longline fishing effort in the southern Ocean and implications for seabirds bycatch. Biol Conserv 114:1–27
Vaske T (1991) Seabirds mortality on longline fishing for tuna in the southern Brazil. Ciencia e Cultura 43:388–390
Veit RR (1995) Pelagic communities of seabirds in the South Atlantic Ocean. Ibis 137:1–10
Vooren CM, Coelho L (2004) Captura incidental de aves oceânicas na pesca com espinhel de fundo. In: Haimovici M, Ávila da Silva AO, Rossi–Wongstschowski CLDB (eds) Prospecção pesqueira de espécies demersais com espinhel de fundo na Zona Econômica Exclusiva da Região Sudeste–Sul do Brasil. Serie Documentos Revizee–Score Sul, São Paulo, pp 85–90
Vooren CM, Fernandes AC (1989) Guia de albatrozes y petréis do sul do Brasil. Sagra, Porto Alegre
Weimerskirch H, Wilson RP (1992) When do wandering albatrosses Diomedea exulans forage? Mar Ecol Prog Ser 86:297–300
Weimerskirch H, Catard A, Prince PA, Cherel Y, Croxall JP (1999) Foraging white-chinned petrels Procellaria aequinoctialis at risk from the tropics to Antarctica. Biol Conserv 87:273–275
Zar JH (1999) Biostatistical analysis. Prentice Hall, New Jersey
Acknowledgments
This work was made possible by the Programa Nacional de Observadores de la Flota Atunera Uruguaya (PNOFA), Departamento de Recursos Pelágicos, Dirección Nacional de Recursos Acuáticos. We thank all the scientific observers who collected the data—especially Alvar Carranza, Marcos Cornes, Alfredo Díaz and Marcel Rodríguez—and skippers, crew and vessel owners of the Uruguayan tuna fleet. Special thanks go to Graham Robertson, Ed Melvin and Leandro Bugoni for their valuable suggestions and review of the manuscript, to Philip Miller for his cooperation with maps, to Maite Pons for her assistance in the statistical analyses and to Stella Weng for the translation and suggestions. Our recognition to Marco Favero and the anonymous referees whose comments and recommendations helped us to improve this work.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Jiménez, S., Domingo, A. & Brazeiro, A. Seabird bycatch in the Southwest Atlantic: interaction with the Uruguayan pelagic longline fishery. Polar Biol 32, 187–196 (2009). https://doi.org/10.1007/s00300-008-0519-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-008-0519-8