
Abstract
The actual breeding population of the Southern Giant Petrel from the northern Patagonian colonies of Argentina on Isla Arce and Isla Gran Robredo was estimated to be 2,300 pairs. The available data sets are small, but showed a positive linear trend at Isla Gran Robredo over the last 14 years, but no trend was detected by a linear model at Isla Arce over a 17-year-period. Our study suggests higher population numbers in recent years. The close presence of increasing penguin, elephant seal and sea lion colonies, the low capture rates by longliners at the Patagonian shelf and/or the extensive use of waste from an increasing fleet of trawlers and jiggers as extra food supply might be some of the reasons for such a population increase.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Southern Giant Petrel (Macronectes giganteus) is considered a “vulnerable” species (IUCN 2004). Total world population has been estimated to be 31,360 pairs, and current information indicates a decline of over 17% with respect to that reported by Hunter (1985) (Patterson et al. 2006). Although general trends show a decline in total breeding population, some colonies have decreased as well as others have increased over the last decades (Patterson et al. 2006). In the southwest Atlantic, for example, a clear increase has been recently reported for colonies in the Malvinas (Falkland) Islands (Huin and Reid 2005). In general, population declines have been attributed to the effects of habitat devastation, human disturbance, introduced predators and fisheries (Rounsevell and Brothers 1984; Jouventin and Weimerskirch 1991; Croxall 1997; Woehler et al. 2003; Brooke 2004; Patterson et al. 2006).
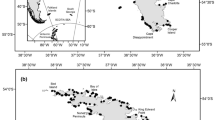
Southern Giant Petrels breed at several sub Antarctic islands, Antarctic Peninsula, southern Chile, Malvinas (Falkland) Islands and Patagonia, Argentina (Harrison 1983; Carboneras 1992; Brooke 2004). In Patagonia, Argentina, Southern Giant Petrels breed at four sites, with an estimated 2,300 breeding pairs in 1995 (Yorio et al. 1999). The northern colonies are located in Golfo San Jorge, found on two Islands separated by 47 km: Isla Arce (45°00′S, 65°29′W) and Isla Gran Robredo (45°08′S, 66°03′W). Southern Giant Petrels in northern Patagonia lay in late October, chicks hatch in late December and fledge between early March and early April (Punta and Saravia 1991; Quintana et al. 2005). Little information is available on the interannual variability in colony size and population trends. As reported by Patterson et al. (2006), current published information on giant petrel population trends is based on few counts at most breeding locations, and both breeding population updates and the development of monitoring programs are urgently needed. Moreover, the recent agreement for the conservation of albatross and petrels (ACAP) encourages the research on population trends of listed threatened species.
In this paper, we report on breeding population counts of Southern Giant Petrels from Isla Arce and Isla Gran Robredo at Golfo San Jorge, Chubut to evaluate population trends. Furthermore, we present data on the breeding performance during several breeding seasons.
Materials and methods
Census
We conducted aerial surveys of both colonies during November 1995–1997 and only the colony of Isla Gran Robredo in 2004. In addition, we visited Isla Arce to make ground counts of breeding pairs during 2001, 2002 and 2004 (see Table 1 for survey dates). Breeding seasons are named for the year during which the eggs were laid (i.e. 2004 breeding season started in October 2004 and lasted until May 2005).
To conduct aerial surveys, we used a two-engined aircraft (CASA 212) during counts in 1995 and 1996, a Puma SA-330L helicopter in 1997, and a Cessna aircraft in 2004. We flew at an altitude of 100–300 m and photographed each colony using 35 mm cameras with 75–205 and 28–105 mm lenses. We estimated colony sizes by counts from the photographs, considering as an Apparently Occupied Nest (following Creuwels et al. 2005), any bird sitting within the colony limits. We also conducted ground counts of the Active Nests (number of nests certainly containing an egg, following Creuwels et al. 2005) (see Census techniques) at Isla Arce from one vantage point (from where all adults sitting within the colony could be seen) located at 30 m from the colony, by using binoculars 8×20.
Population trends
Population trend analysis included our own data since the 1995 breeding season, supplemented with information from two previous published counts (Isla Arce during 1987, Punta 1989, and Isla Gran Robredo for 1990, Yorio and Harris 1997; see Table 1). To analyse and detect statistically significant trends in the data, we followed methods by Woehler et al. (2001) and Creuwels et al. (2005).
Census techniques and potential error source
Preliminary observations have shown that a certain proportion of non-incubating birds may spend time sitting within the colony limits without actually having a nest site (Apparently Occupied Nests, sensu Creuwels et al. 2005, F. Quintana and S. Copello, personal observation). To determine the proportion of non-incubating birds registered during the aerial counts, we gathered additional information at Isla Arce on 1 December 2004 and Isla Gran Robredo on 2 December 2004. From a vantage point (at a distance from the colony periphery to avoid disturbance to the birds), we took digital photographs of three randomly chosen sectors at each of the colonies. Immediately after taking the picture, one of us slowly approached the nests within the photographed sector. We assumed that birds that have no nest leave the area due to disturbance. We took a second picture of only the birds that were actually incubating (Active Nests of the ground counts, see above). We repeated the procedure in the same three sectors of the colonies at intervals of 2 h. At Isla Gran Robredo, five times (from 1000 to 1800 h) and at Isla Arce two times (1700 and 1900 h). We counted birds in each of the photographs, and we averaged numbers from the three sectors within each set of photographs.
Breeding performance
We estimated the breeding performance in both colonies counting sitting birds during incubation and chicks during the chick—rearing period (Isla Arce: during six breeding seasons and Isla Gran Robredo: during three breeding seasons; Table 1). Between 1995 and 1997, we conducted aerial surveys from late February to early April, using a CASA 212 aircraft and counting the number of chicks on the nests from aerial photographs. During this period, chicks can be easily recognized by their conspicuous dark plumage. In addition, they usually remain alone in their nests and less than 3% of the adults are usually seen attending the colony at this stage between approximately 1000 and 1600 h (F. Quintana and S. Copello, personal observation). Since the 2001 breeding season, we also counted the number of chicks present in the colonies through ground surveys made by three observers walking the whole colony area (see Table 1 for detail of years and colonies surveyed). We used log-likehood test (G-statistics) and χ 2 to compare the breeding performance between seasons and colonies.
Results
Colony sizes and trends
Aerial and ground surveys showed that Southern Giant Petrels bred at both islands during the sampled seasons. Colonies at both islands were located in open rocky substrate on the northern sector of Isla Arce and on the western sector of Isla Gran Robredo. The last count (2004) indicated a Southern Giant Petrel breeding population at northern Patagonia (both islands) of approximately 2,300 pairs, with more than 80% of pairs breeding at Isla Gran Robredo. No white morph birds were seen nesting on the islands.
Data sets spanning over 17 and 14 years were available for Isla Arce and Isla Gran Robredo, respectively. Population estimates on Isla Arce varied between 150 and 500 pairs and on Isla Gran Robredo between 1,078 and 1,883 (Table 1, Fig. 1).

Plots of breeding population (pairs) and trends fitted by linear models to both northern Patagonian colonies
On Isla Arce there was no linear trend between 1987 and 2004, based on seven counts (VE = 22.9%, P = 0.2). On Isla Gran Robredo, we found a significant increase between 1990 and 2004, based on five counts (VE = 68.7%, P = 0.01) (Fig. 1). A significant increase for the Gran Robredo colony was observed even when removing the 1990 count (P = 0.003).
Census techniques and potential error source
The percentage of non-incubating birds within the study area varied depending on the time of day, between 7 and 9.6% (1800 and 1400 h, respectively) at Isla Gran Robredo, and between 5.6 and 10.3% (1900 and 1700 h, respectively) at Isla Arce. On average, numbers obtained from the first photograph (undisturbed condition) were 8.7 ± 4.2% (n = 5) larger than the second (disturbed condition) at Isla Gran Robredo, and 8.0 ± 10.0% (n = 2) larger at Isla Arce.
Breeding performance
In both colonies, the breeding performance was variable and significant differences were found between seasons (G = 53.7 for Isla Arce and G = 68.4 for Isla Gran Robredo, both P < 0.001). The percent of nests with chicks during late season counts (late February to early April) varied between 54.3 and 98.9% at Isla Arce and between 63.9 and 91.7% at Isla Gran Robredo (Table 1). Difference in breeding performance between colonies were found during the 1996 (χ 2 = 17.9) and 2004 (χ 2 = 31.0) seasons (both P < 0.001), being 23.7% higher at Isla Gran Robredo in 1996 and 34.6 higher at Isla Arce in 2004 (Table 1). Such differences still persist when the aerial counts were corrected to ground counts according to an average estimated error for this colony.
Discussion
Colony sizes and population trend
The colonies in northern Patagonia (with 2,300 breeding pairs in 2004) represent over 80% of the total breeding population of Southern Giant Petrels in Patagonia. Only two other Southern Giant Petrel breeding sites have been reported in Argentina, both of them located in Isla de los Estados, Tierra del Fuego (Schiavini et al. 1999). The number of breeding pairs were estimated at 500 and less than 30 in Isla Observatorio and Península López, respectively (Quintana et al. 2005).
Previous records of colony sizes in northern Patagonia are scarce. Southern Giant Petrels were first reported breeding at Isla Arce in 1982 (Humphrey and Livesey 1983). They estimated between 75 and 100 nests based on ground counts during chick stage. Humphrey and Livesey (1983) also mentioned the existence of a Southern Giant Petrel colony at Isla Gran Robredo, although they did not visit it. The first published record of the Isla Gran Robredo colony is from 1988 (Punta and Saravia 1991). This colony was visited on February 8, and the number of nests, mostly with chicks, was estimated through ground counts at 450 ± 50. The last estimated numbers of breeding pairs obtained at both islands in 2004 were 190 and 75% larger than those estimated using the same census methodology in 1987 (Punta 1989) and 1990 (Yorio and Harris 1997) at Isla Arce and Isla Gran Robredo, respectively. These counts and those reported in the literature (see above) seem to give a clear evidence of increase in the size of both colonies. Previous counts reported by Humphrey and Livesey (1983) and Punta and Saravia (1991) were made during the chick stage, underestimating the total breeding population due to nest losses. Despite different census methods and different timing of the census, the large differences between these early counts and our results suggest that the breeding population of northern Patagonia has increased during the last 17 years. However, given that significant interannual variation in the number of breeding individuals has been reported for this species (Hunter 1984; Voisin 1988; Woehler et al. 2003; this study), care must be taken when interpreting census results to determine the real magnitude of population changes and the definition of population trends.
Finally, the two trend analysis made for the colony at Isla Gran Robredo (with and without counts before 1994) showed a linear increase, but no linear trend was detected for the one at Isla Arce (trend did not change when the four ground counts were corrected to aerial counts). However, in both colonies we found a higher number of breeding Southern Giant Petrels after 1995. More counts over a longer time period are needed to infer real population trends. We are aware that our analysis derived from a relatively small series of counts should be considered with caution.
Census techniques and potential error source
Monitoring of breeding Southern Giant Petrels requires the standardization of census methods, considering both the timing and counting techniques, in order to correctly interpret long-term population trends (Creuwels et al. 2005). Aerial photographs proved to be adequate for colony size estimations in our study area. Counts of Apparently Occupied Nests (sitting adults) (see Creuwels et al. 2005) from aerial photographs may include adult birds which are loafing within the colony limits. Our data shows that this count may overestimate the number of nests that really contain an egg (Active Nests, following Creuwels et al. 2005) by less than 10%, which is an acceptable error for the purposes of long-term monitoring. Differences between aerial and ground counts of less than 10% were also found in surveys of Southern Giant Petrels in the Malvinas (Falkland) Islands (Huin and Reid 2005). Creuwels et al. (2005) found that counts of Apparently Occupied Nests estimate the number of Active Nests with ca 10%. However, even though aerial surveys are a convenient method for monitoring colony sizes due to the difficulties of accessing the islands by boat, future census work should consider applying a correction factor to compensate for the presence of non or failed breeding individuals within the colony limits at the time of the census.
Breeding performance
During March counts, most Southern Giant Petrel chicks were older than 2 months, an age after which mortality has been considered to be minimal in the study area (Quintana et al. 2005) and at sub Antarctic breeding sites (Hunter 1984). Therefore, chick counts during this period may be good indicators of overall breeding performance. The proportion of successful nest at this time of the season in both colonies ranged between 54 and 99%. These figures are similar to those reported for the breeding performance of Southern Giant Petrels at sub Antarctic islands (Warham 1962; Conroy 1972; Voisin 1988) except for the 1997 and 2004 breeding seasons at both colonies and Isla Arce, respectively, when the proportion of surviving chicks was very high. As we counted after the beginning of the egg-laying period, we missed the early egg-losses, which can be considerable in fulmarine petrels (Warham 1990). Therefore we cannot calculate the overall breeding success, based on the number of chicks fledged out of all eggs laid. Due to differences in the timing and methods of censuses, published values for the breeding performance are difficult to compare. Although our breeding performance seems to be high, they are not very different from the values from South Georgia (Hunter 1984), Frazier Islands (Creuwels et al. 2005) and Malvinas/Falklands (Reid and Huin 2005), but probably higher than Marion Island (Cooper et al. 2001).
The northern Patagonian population in the Patagonian shelf
Our results suggest that the populations of Southern Giant Petrels in northern Patagonia, Argentina, have increased or reached, at least, higher stable levels. Similarly, the Malvinas (Falkland) Islands population has shown a much larger population than the previously estimated (Reid and Huin 2005). These two southwest Atlantic population of the Southern Giant Petrel inhabit the large Patagonian shelf, a highly productive ocean ecosystems in the southern hemisphere (Brandini et al. 2000; Croxall and Wood 2002; Acha et al. 2004).
Even though Southern Giant Petrel mortality due to fisheries has been noted in the Patagonian shelf waters, the capture rates are too low to significantly affect their population status (Sullivan and Reid 2002; Favero et al. 2003; Sullivan and Reid 2003, 2004). Despite the low fisheries-related mortality, trawlers are an important source of food for Southern Giant Petrels through the extensive use of fishery waste (Copello and Quintana 2003). On the Patagonian shelf, Argentine fisheries (trawlers and jiggers) have increased to 60% in the last decade (SAGPyA 2005) and flocks of up to 500 Southern Giant Petrels (T. Reid, personal observation) frequently occur around trawlers (see also Yorio and Caille 1999; González Zevallos and Yorio 2006).
The Southern Giant Petrel is an opportunistic scavenger (Hunter 1983, 1985; Brooke 2004) and the availability of penguin and pinniped carcasses might be another reason for higher numbers. The northern Patagonian Southern Giant Petrels breed in proximity of large colonies of these species. Even though Magellanic Penguin, Spheniscus magellanicus, colonies and South American Sea Lion, Otaria flavescens, rookeries occur all along the Patagonian coast, the highest abundances are concentrated at the San Jorge Gulf and its vicinities to less than 100 km from the Southern Giant Petrel colonies (S. Copello and F. Quintana, unpublished data). Satellite tracking studies of Southern Giant Petrels breeding at Isla Arce and Isla Gran Robredo revealed frequent visits to nearby large colonies of penguin and sea lion (F. Quintana and S. Copello, unpublished data). These species show different population trends, and while South American Sea Lions have shown a population recovery in the last decades (Reyes et al. 1999; Dans et al. 2004), Magellanic penguin numbers have increased or declined depending on the colony considered (Schiavini et al. 2005). Both species constitute important prey items in the diet of adult birds from both islands during the chick rearing period (88 and 42% of dietary occurrence for penguins and sea lions, respectively) (Quintana et al. 2005; S. Copello and F. Quintana, unpublished data).
Both northern Patagonian colonies could also benefit by the relatively low human disturbance at their breeding sites. However, despite their importance to the overall Patagonian population, these colonies currently lack any legal protection. Particular attention should be paid to visiting regulations of the islands, given the high sensitivity of breeding Giant Petrels to human disturbance (Warham 1990; Woehler et al. 2003; Brooke 2004; Patterson et al. 2006) and the increasing interest in visiting wildlife areas by cruise vessels (Yorio et al. 2001).
References
Acha EM, Mianzan H, Guerrero R, Favero M, Bava J (2004) Marine fronts at the continental shelves of austral South America: physical and ecological processes. J Marine Syst 44:83–105
Brandini F, Boltovskoy D, Piola AR, Kocmur S, Rottgers R, Abreu P, Mendes Lopes R (2000) Multi-annual trends in fronts and distribution of nutrients and chlorophyll in the southwestern Atlantic (30–62°S). Deep-Sea Res I 47:1015–1033
Brooke M (2004) Albatrosses and petrels of the world. Oxford University Press, Oxford
Carboneras C (1992) Handbook of the birds of the world, Barcelona
Conroy JWH (1972) Ecological aspects of the biology of the giant petrel Macronectes giganteus (Gmelin) in the maritime Antarctic. British Antarctic Survey. Sci Rep 75:1–74
Cooper J, Brooke ML, Burger AE, Crawford RJM, Hunter S, Williams T (2001) Aspects of the breeding biology of the Northern giant petrel (Macronectes halli) and the Southern giant petrel (M. giganteus) at sub-Antarctic Marion Island. Int J Ornithol 4:53–68
Copello S, Quintana F (2003) Marine debris ingestion by Southern giant petrels and its potential relationships with fisheries in the Southern Atlantic Ocean. Mar Pollut Bull 46:1513–1515
Creuwels JCS, Stark JS, Woehler EJ, Van Franeker JA, Ribic CA (2005) Monitoring of a Southern giant petrel Macronectes giganteus population on the Frazier Islands, Wilkes Land, Antarctica. Polar Biol 28:483–493
Croxall JP (1997) Research and conservation: a future for albatrosses? In: Robertson G, Gales R (eds) Albatross biology and conservation. Surrey Beatty & Sons, Chipping Norton, pp 267–288
Croxall JP, Wood AG (2002) The importance of the Patagonian Shelf for top predator species breeding at South Georgia. Aquat Conserv 12:101–118
Dans SL, Crespo E, Pedraza SN, Koen Alonso M (2004) Recovery of the South American sea lion (Otaria flavescens) population in northern Patagonia. Can J Fish Aquat Sci 61:1681–1690
Favero M, Khatchikian C, Arias A, Silva Rodriguez MP, Cañete G, Mariano-Jelicich R (2003) Seabird by-catch along the Patagonian Shelf by Argentine Longline Fishing Vessels: estimations based on data from the Observers National Program, period 1999–2001. Bird Conserv Int 13:273–281
González Zevallos D, Yorio P (2006) Seabird use of discards and incidental captures at the Argentine hake trawl fishery in Golfo San Jorge, Argentina. Mar Ecol Prog Ser (in press)
Harrison P (1983) Seabirds: an identification guide. Beckenham-UK-Croom-Helm
Huin N, Reid T (2005) Census of the giant-petrel population of the Falkland Island. Falkland Conservation, Falkland Island
Humphrey PS, Livesey BC (1983) Giant petrels (Macronectes giganteus) nesting in Chubut, Argentina. Le Gerfaut 73:3–8
Hunter S (1983) The food and feeding ecology of the giant petrels Macronectes halli and M. giganteus at South Georgia. J Zool 200:521–538
Hunter S (1984) Breeding biology and population dynamics of giant petrel Macronectes at South Georgia (Aves: Procellariiformes). J Zool Lond 203:441–460
Hunter S (1985) The role of giant petrels in the Southern Ocean ecosystem. In: Siegfried WR, Condy PR, Laws RM (eds) Antarctic nutrient cycles and food webs, pp 534–542
IUCN (2004) 2004 IUCN Red list of threatened species. http://www.iucnredlist.org
Jouventin P, Weimerskirch H (1991) Changes in the population size and demography of southern seabirds: management implications. In: Perris CM, Leberton JD, Hirons GJM (eds) Bird population studies: their relevance to conservation and management. Oxford University Press, Oxford, pp 297–314
Patterson DL, Woehler EJ, Croxall JP, Cooper J, Poncet S, Freser WR (2006) Breeding distribution and population status of the northern giant petrel Macronectes halli and the southern giant petrel M. giganteus. Mar Ornithol 28 (in press)
Punta G (1989) Guaneras de la Provincia del Chubut. Potencialidad productiva y fundamentos para su manejo racional. Dirección de Intereses Marítimos y Pesca Continental de la Provincia de Chubut
Punta G, Saravia J (1991) A newly discovered colony of southern giant petrels Macronectes gigantes on isla Gran Rob redo, Chubut Province, Argentina. Mar Ornithol 19:131–133
Quintana F, Schiavini A, Copello S (2005) Estado poblacional, ecología y conservación del Petrel Gigante del Sur (Macronectes giganteus) en Argentina. El Hornero 20:25–34
Reid T, Huin N (2005) Census of the Southern giant-petrel population of the Falkland Islands 2004/2005. Project funded by Overseas Territories Environmental Programme and Falkland Islands Government
Reyes LM, Crespo E, Szapkievich V (1999) Distribution and population size of the Southern Sea Lion (Otaria flavescens) in central and southern Chubut, Patagonia, Argentina. Mar Mamm Sci 15:478–493
Rounsevell DE, Brothers NP (1984) The status ans conservation of seabirds at Macquarie Island. In: Croxall JP, Evans PG, Schreiber RW (eds) Status and conservation of the world’s seabirds. ICBP Technical Publication no 2, Cambridge, pp 587–592
SAGPyA (2005) Secretaría de Agricultura, Ganadería, Pesca y Alimentos. República Argentina. http://www.sagpya.mecon.gov.ar/
Schiavini ACM, Frere E, Yorio P, Parera A (1999) Las aves marinas de la Isla de los Estados, Tierra del Fuego, Argentina: revisión histórica, estado poblacional y problemas de conservación. Anales del Instituto de la Patagonia, Serie Ciencias Naturales (Chile) 27:25–40
Schiavini A, Yorio P, Gandini P, Raya Rey A, Boersma D (2005) Los pingüinos de las costas argentinas: estado poblacional y conservación. El Hornero 20:5–23
Sullivan BJ, Reid T (2002) Seabid interactions/mortality with longliners and trawlers in Falkland Island waters 2001/02. Falkland Conservation, Stanley
Sullivan BJ, Reid T (2003) Seabird mortality in longliners and trawlers in Falkland Island waters 2002/03. Falkland Conservation, Stanley
Sullivan BJ, Reid T (2004) Seabid mortality in fisheries and mitigation techniques in Falkland Island waters 2003/04. Falkland Conservation, Stanley
Voisin JF (1988) Breeding biology of the Northern giant petrel Macronectes halli and the Southern giant petrel M. giganteus at ile de la Possession, Iles Crozet 1966–1980. Cormorant 16:65–97
Warham J (1962) The biology of the giant petrel Macronectes giganteus. Auk 79:139–160
Warham J (1990) The petrels, their ecology and breeding systems. Academic Press, London
Woehler EJ, Cooper J, Croxall JP, Fraser WR, Kooyman GL, Miller GD, Nel DC, Patterson DL, Peter H-U KM, Ribic CA, Salwicka K, Trivelpice WZ, Weimerskirch H (2001) A statistical assessment of the status and trens of Antarctic and sub Antarctic seabirds. SCAR
Woehler EJ, Riddle MJ, Ribic CA (2003) Long-term population trends in the southern giant petrels in East Antarctica. In: Huiskes AHL, Gieskes WWC, Rozema J, Schorno RML, van der Vies SM, Wolff WJ (eds) Antartic biology in a global context. Backhuys Publishers, Leiden, pp 290–295
Yorio P, Caille G (1999) Seabird interactions with coastal fisheries in northern Patagonia: use of discards and incidental captures in nets. Waterbirds 22:201–216
Yorio P, Harris G (1997) Distribución reproductiva de aves marinas y costeras coloniales en Patagonia. Relevamiento aéreo Bahía Blanca-Cabo Vírgenes, noviembre 1990. Fundación Patagonia Natural, 29, Puerto Madryn, Argentina
Yorio P, Frere E, Gandini P, Conway W (1999) Status and conservation of seabirds breeding in Argentina. Bird Conserv Int 9:299–314
Yorio P, Frere E, Gandini P, Schiavini A (2001) Tourism and recreation at seabird breeding sites in Patagonia, Argentina: current concerns and future prospects. Bird Conserv Int 11:231–245
Acknowledgments
We thank Gonzalo Herrera and Ricardo Vera for assistance and data gathering. We are grateful to Dr. J. C. Creuwels for his dedicated and constructive comments during the revision of the manuscript. We also thank the Dirección General de Intereses Marítimos y Pesca Continental of the Chubut Province for institutional support and Prefectura Naval Argentina (PNA) for logistical support. We specially thank Maximiliano Cosentino, helicopter pilot of the PNA, who died during a rescue operation at sea shortly after conducting our survey.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Quintana, F., Punta, G., Copello, S. et al. Population status and trends of Southern Giant Petrels (Macronectes giganteus) breeding in North Patagonia, Argentina. Polar Biol 30, 53–59 (2006). https://doi.org/10.1007/s00300-006-0159-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-006-0159-9