
Abstract
The parasite faunas of the Patagonian toothfish Dissostichus eleginoides from six locations around the Southern Ocean were studied and compared. Thirty-two parasite taxa were found. Ten parasite species are reported from D. eleginoides for the first time and some other previously reported species were new locality records. Sample size at Shag Rocks was sufficient to examine the effect of intrinsic host factors, including sex and length, on the parasite fauna and these results are discussed here. Some parasite species were found only in certain areas. Sørensen’s similarity index indicated that the parasite faunas at Heard, Maquarie and Prince Edward Islands were the most similar, while those from the Ross Sea was the most dissimilar. There may be a gradual decrease in parasite diversity the further east the samples were collected around the Southern Ocean.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Studies on Antarctic fish parasites are limited. Most studies deal with parasite taxonomy, and few deal with parasite life-cycles, biology and ecology (see Holloway and Spence 1980; Moser and Cowen 1991; Palm 1999; Palm et al. 1998; Wägele 1988). Most of these are restricted to the perciform suborder, the Notothenioidei from coastal or shelf waters (e.g. Zdzitowiecki 1991, 1997). Studies of fish parasites on the continental slope and deep-sea are limited to Macrourus whitsoni (Walter et al. 2002).
The Patagonian toothfish, Dissostichus eleginoides, is the largest member of the family Nototheniidae. Individuals of more than 2 m total length have been regularly recorded (Paul Brickle, personal observation; Sosinski and Kuranty 1979; Duhamel 1987). It has an elongate fusiform body with a large broad head which has dorsal naked areas, big eyes and large jaws with canine teeth, with robust canine teeth on each premaxilla (Norman 1937) and: “... its predatory appearance is enhanced by its projecting lower jaw” (Eastman 1993).
Dissostichus eleginoides is widely distributed in the Southern Ocean. Along the coasts of South America it probably extends as far north as any notothenioid (Eastman 1993). On the east coast it occurs near Uruguay (35°S) (Meissner and Zaostrovtsev 1983), but on the west coast it reaches north to 20°S (Oyarzun and Campos 1987). The cold, northwards flowing Peru Current, on the west coast, probably accounts for its extended range. It is also found at Subantarctic islands of South Georgia, Bouvet, Prince Edward, Crozet, Kerguelen, Macquarie, and Heard and around the Ob and Lena Banks (Norman 1938; Hart 1946; Andriashev 1964; Yukhov 1982; Meissner and Zaostrovtsev 1983; Duhamel and Pletikosic 1983; Kock et al. 1985). D. eleginoides also occurs at the Antarctic Penninsula, King George Island (61° 24’S) in the South Pacific (Arana and Vega 1999) and is regularly caught in the Ross Sea (Smith and McVeagh 2000).
The life-cycle of the Patagonian toothfish comprises several ontogenetic phases. Upon attaining 12–15 cm TL, semi-pelagic juveniles become demersal at 150–400 m depth, and after several years grow to 60–70 cm TL. Adult fish then migrate to deepwater meso- and bathypelagic habitats at depths greater than 1,000 m (Eastman 1993). Thus, D. eleginoides is an interesting species to study because early in life it is exposed to parasites that occur on the shelf and, when older to deep-water parasites.
Parukhin and Lyadov (1982) examined D. eleginoides from off Kerguelen and Crozet for parasites, while Gaevskaya et al. (1990) studied the parasite fauna of 24 specimens from South Georgia, and the Ob (26 host specimens) and Lena Banks (50 host specimens) below the Antarctic Convergence, and recovered 38 parasite species. Gaevskaya et al. (1990) referred to the data of Parukhin and Lyadov (1982) and suggested that the parasite fauna of D. eleginoides was richer in the western than in the eastern part of its range. They also suggested that D. eleginoides originated in the Falkland-Patagonian and South Georgia region and then spread eastwards through the Antarctic. This view is supported by Yukhov (1982). Rodriguez and George-Nascimento (1996) examined the parasite fauna of D. eleginoides off central Chile. The only other reports of parasites of D. eleginoides are those describing new species (Gaevskaya and Kovaleva 1976, 1978; Parukhin and Lyadov 1982; Rodyuk 1984, 1986; Brickle et al. 2001).
The aim of this study was to describe and compare the parasite faunas of D. eleginoides found in different parts of the Antarctic and to contribute to our knowledge of the biogeography and host specificity of Subantarctic parasites.
Materials and methods
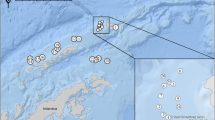
Figure 1 shows the sampling locations around the Southern Ocean for the present study. A total of 197 D. eleginoides were caught from various locations around the Antarctic and Subantarctic between, February 1997 and April, 2000 (Table 1).

Sampling locations of Dissostichus eleginoides samples collected for this study
Total length to the nearest centimetre below and the total weight (g) of each fish were recorded. To collect parasites the following procedure was adopted where only the viscera were retained. The head was removed and a cut around the exposed oesophagus was made, taking care not to damage the liver or the gallbladder. The oesophagus was then tied off to avoid loss of stomach contents. The abdominal cavity was cut from just anterior to the anus right up to the collar. The rectum was tied off and cut to the posterior of the tie. The viscera were removed carefully and placed in a plastic bag. The kidneys were scraped out with a spoon or similar shaped instrument and placed in a separate plastic bag, which was then placed in the bag containing the viscera. The gills were removed and placed into the bag containing the viscera and kidneys. The numbers of the parasitic copepod Eubrachiella antarctica were noted on waterproof paper along with the vessel’s callsign, the station and the host’s biometeric data and this too was placed in the bag with the viscera. The sample bags were then frozen within 1 h of processing. These samples were brought back to Port Stanley, Falkland Islands for examination. Those caught from Heard and Macquarie Islands and the Ross Sea were flown to the School of Biological Sciences of Aberdeen University. Samples from Prince Edward Island were shipped to the Institute of Marine and Coastal Management, Cape Town, and examined there.
Whole fish caught on the South Georgia shelf, were returned to Stanley, where they were defrosted individually in trays filled with warm water. The skin surface was examined in strong light, and squash preparations made of cysts or lesions were examined under a compound microscope at magnifications of up to 400X. The fish’s total length and weight were noted. The fins were then removed and examined microscopically as were lesions or cysts. The fish was washed in its tray of water, removed and the remaining water and mucus poured through a fine meshed sieve to collect any parasites that were not observed during the initial visual inspection. The fish were opened as described above and the viscera removed.
The internal organs (heart, liver, spleen, gall bladder, gonads, kidney, urinary bladder and digestive tract) were examined for parasites either free or encapsulated on the exterior surfaces, separated and then individually examined. The stomach, pyloric caeca and intestine were separated and opened longitudinally. In smaller fish the mucosa of the digestive tract was scraped with the handle of a scalpel to remove any parasites. Larger fish with substantial quantities of mucus in their digestive tracts were also opened longitudinally and their contents rinsed into beakers and mixed with sodium bicarbonate and settled to remove endoparasitic helminths (Arthur and Albert 1993). The walls of the stomach, pyloric caeca and intestine, and the whole liver, spleen, kidney and heart were compressed between glass plates and examined for parasites. The body-cavity was rinsed and the rinse collected and examined. Squash preparations were made from the liver, spleen, kidney, gonads, intestine and both muscle and brain tissue, and scrapings from the urinary and gallbladder were microscopically examined for protozoan and myxozoan parasites microscopically at 400X under bright field, phase contrast and Nomarski interference contrast illumination. Preparations not found to harbour parasites within 5 min were considered to be uninfected. The gills were removed, rinsed and the arches examined individually. The buccal cavity was rinsed and the opercula and eyes were rinsed separately. The body musculature was removed from the vertebral column, the skin removed from the fillets and the muscle flaps were thinly sliced and inspected for helminths and myxozoan cysts. All parasites were sorted into major taxonomic groups, cleaned and counted for each organ, and the data noted on an examination sheet and subsequently entered into a database.
Juvenile D. eleginoides caught around Shag Rocks provided a sufficient sample size (n=72) to examine the effect of intrinsic host factors, particularly length and sex, on the parasite fauna. However, only those parasites with prevalences greater than 5% were analysed. As many parasites had overdispersed frequency distributions (high variance to mean ratio), the abundance of infection was transformed as log (n+1) to reduce the variance (which helped to normalise the data; Elliot 1971) and was compared to intrinsic host factors using the Pearson’s correlation coefficient (r) and simple linear regression. The effect of fish sex on the parasite fauna was examined using the non-parameteric Mann–Whitney test. New host records and locality records were examined.
The terms prevalence, mean intensity and mean abundance follow the recommendations of Bush et al. (1997). The degree of similarity of the parasite fauna of the different populations of D. eleginodes was analysed using the Sørensen similarity index (Sørensen 1948).
Results
A total of 32 parasite taxa were recorded from D. eleginoides in this study (Table 2 and 3). Ten species were the first records and some parasites previously reported from D. eleginoides were new locality records.
Shag Rocks
Dissostichus eleginoides caught in the Shag Rocks area ranged between 16 cm and 68 cm total length and their sex ratio was 1:1. All fish were immature (maturity stage 1) with an exception of a 63 cm male at maturity stage 2 (mature resting).
Seventeen metazoan parasite taxa and 2,076 individual parasites were recovered from 72 D. eleginoides from the Shag Rocks area (Table 2). The parasite fauna consisted of two monogeneans, four digeneans, two cestodes, five nematodes, two acanthocephalans, one copepod and gill cysts of unknown aetiology (CUEs). Since there was no significant difference in the mean abundance of parasites with host sex (P>0.05, Mann–Whitney U=856.5) samples from both sexes were combined in subsequent analyses.
The abundances of some parasite taxa were either positively or negatively correlated with fish TL and there were notable changes in prevalence with fish TL. Out of ten parasite taxa with prevalences of greater that 5%, the abundance of seven of these were significantly correlated with fish TL. Tetraphyllidean plerocercoids (=Scolex pleuronectis and S. polymorphus) were significantly negatively correlated (P<0.01, r= −0.988) with host length, and decreased in prevalence (Fig. 2). Abundances of the larval acanthocephalan Corynosoma bullosum, the copepod Eubranchiella antarctica, the nematodes Contracaecum sp. and Hysterothylacium sp., the monogenean Pseudobenedenia dissostichi and CUEs all showed significant positive correlations and increasing prevalences with increasing host length (Fig. 2). The abundances of three taxa, the digenean Gonocerca phycidis, the monogenean Neopavlovskioides geogianus and the nematode Anisakis sp. showed no correlation with host length. However, N. georgianus and Anisakis sp. both showed an increase in prevalence with increasing host length.

Correlation analysis between host length and abundance of parasites and the relationship between increasing host length and parasite prevalence. a Tetraphyllidean cercoids. b Corynosoma bullosum, c Eubrachiella antarctica, d Contracaecum sp., e Hysterothylacium sp., f Pseudobenedenia dissostichi, and g CUEs
South Georgia
Twenty-one adult D. eleginoides, ranging between 68 cm and 130 cm total length, were examined from around South Georgia. All of the fish were mature (gonad maturities of stage two to or greater).
Twenty-two parasite taxa, including a total of 7,251 metazoan individuals, were recovered from South Georgia toothfish (Table 3), comprising one microsporidean, one myxosporean, one monogenean, four digeneans, three cestodes, seven nematodes, three acanthocephalans, one copepod, and CUEs in the gills. Two species, the microsporidean sp. 1 and the myxosporean Alataspora sp., represent new host records for D. eleginoides and, because they were considered oioxenic, they also represented new locality records. The nematode Capillaria sp. found in this study was the second record for this genus in the Antarctic and a new host record for D. eleginoides.
Prince Edward Island
A total of 31 D. eleginoides were examined ranging in size from 61 cm to 114 cm total length, of which 9 (29 %) were considered to be sexually immature.
Twenty-three parasite taxa, including a total of 4,163 metazoan individuals, were recovered (Table 3), comprising one myxosporean, one monogenean, seven digeneans, two cestodes, six nematodes, four acanthocephalans, one copepod and CUEs in the gills. New host records for D. eleginoides were the digeneans Stenakron sp. and Lecithophyllum champsocephali and the acanthocephalan Aspersentis megarhynchus. New locality records included the myxosporean Alataspora sp., the monogenean N. georgianus, the nematode Capillaria sp., the larval acanthocephalans C. bullosum, C. arctocephali and Echinorhynchus petroschenkoi, the copepod E. antarctica, the digeneans Neolepidapedon magnatestis, Derogenes varicus, Elytrophalloides oatesi and Lecithophyllum champsocephali, and the larval cestodes Hepatoxylon trichiuri and Tetraphyllidean plerocercoids.
Heard Island
Twenty-six fish were examined ranging with a total length of 40–84 cm of which 22 (84%) were sexually immature.
Twenty-two parasite taxa, including 1,390 metazoan individuals, were recovered (Table 3), comprising one myxosporean, one monogenean, seven digeneans, two cestodes, six nematodes, three acanthocephalans, one copepod and CUEs in the gills. In the Heard Island area, the only new host record is represented by the digenean species 1. New locality records for Heard Island included the digeneans Lecithaster macrocotyle and Lecithophyllum champsocephali, and the acanthocephalans C. arctocephali and E. petroschenkoi.
Macquarie Island
Twenty-nine fish had a total length of 54–110 cm, of which 3 (10%) were considered to be immature.
Twenty-one parasite taxa, including 2,995 metazoan individuals, were recovered (Table 3), comprising one myxosporean, one monogenean, five digeneans, four cestodes, six nematodes, two acanthocephalans, one copepod and CUEs on the gills. Lecithochirium sp. was a new host record for D. eleginoides. This may be a new species but requires further study. New locality records were the myxosporean Alataspora sp., the monogenean N. georgianus, the digeneans G. physidis, G. macrouri and L. champsocephali, the nematode Capillaria sp., and the acanthocephalan C. arctocephali.
Ross Sea
Eighteen D. eleginoides were examined with a total length 81–126 cm. Of these 3 (16%) were immature.
The sample of Ross Sea D. eleginoides had the lowest diversity of parasites, yielding 13 taxa and 1,102 metazoan individuals (Table 3). The parasite fauna consisted of one myxosporean, three digeneans, one cestode, three nematodes and three acanthocephalans. Of these only the digenean Helicometra antarctica was a new host record for D. eleginoides. New locality records were N. georgianus, N. magnatestis, G. macrouri, Tetraphyllidean plerocercoids and C. bullosum.
Parasitological similarity and zoogeography.
There seems to be a small reduction in parasite diversity from west to east around the SubAntarctic, with 22 and 23 species in South Georgia and Prince Edward Island, respectively, then a decline to 21 species for Heard and Macquarie Islands and, a large decrease to 13 species for the Ross Sea.
Sørensen’s similarity index (Table 4) illustrated that the parasite faunas of D. eleginoides were similar around the SubAntarctic but those from the Ross were least similar. The parasite faunas of Prince Edward, Heard, and Macquarie Islands were the most similar.
Discussion
The diverse parasite fauna of D. eleginoides is comparable to that of other predatory fish in the Subantarctic and Antarctic. Palm et al. (1998) recorded 37 species from the Antarctic rock cod Notothenia coriiceps and Walter et al. (2002) reported 22 species from the Antarctic grenadier Macrourus whitsoni. Gaevskaya et al. (1990) reported 23 species from D. eleginoides in Antarctic areas. In the present study, most of the parasites were adult, using toothfish as a final host. However, several larval forms were also recovered, suggesting that toothfish may serve as important intermediate or paratenic hosts. The recovery of only three species of ectoparasites, supports Rohde’s (1999) suggestion that in high latitudes ectoparasite diversity is lower but it is higher for endoparasites.
Immature D. eleginoides from the Shag Rocks area had a lower parasite diversity compared with the other areas in this study except for the Ross Sea. This may be because larger fish provide more internal and external space and offer a larger area for parasite establishment and can support higher infection rates, because they eat more infected prey (Poulin 2000). des Clers (1991) developed simple mathematical models to describe this. In the case of the Ross Sea, D. eleginoides is at the southern edge of its geographical range and thus, has a reduced parasite fauna. The sample size of immature D. eleginoides from the Shag Rocks area was sufficient to examine the effect of host sex and host length on the parasite fauna. Host sex was not significant. Out of ten parasite species that had prevalences of greater than 5%, seven showed statistically significant correlations with fish size. Immature fish are known to feed on krill (Duhamel 1991) and they become more piscivorous as they increase in age/size (Arkhipkin et al. 2003). Tetraphyllidean plerocercoids use copepods and euphausiids as intermediate hosts (Hochberg 1990), that are then ingested by juvenile D. eleginoides, which serves as a further intermediate or paratenic host. As D. eleginoides increase in size/age, they prey less on copepods and euphausiids, which could explain the pattern of decreasing abundance and prevalence of Tetraphyllidean plerocercoids with host length.
The abundance of the larval acanthocephalan C. bullosum had a significant positive correlation and showed an increase in prevalence with host length, suggesting that this parasite accumulates with host length/age. Corynosoma spp. use amphipods as their first intermediate hosts and fish of different species play a major role as paratenic hosts. Zdzitowiecki (1991) found that predatory fish species were the most heavily infected with Corynosoma spp. around South Georgia. During the present study one D. eleginoides was infected with over 500 C. bullosum (Table 2). Juvenile Corynosoma spp. infected the mesenteries, abdominal wall, stomach and intestinal wall of toothfish. There were dead encapsulated cystacanths in some individuals. C. bullosum is considered to be part of the permanent fauna of D. eleginoides and accumulates as the fish grow. Increasing intensity of Corynosoma sp. with increasing host length has been observed in the Chilean hake Merluccius gayi (George-Nascimento 1996). This phenomenon was also evident in larval Contracaecum sp. infecting immature D. eleginoides and its accumulation with host length could have similar patterns to juvenile Corynosoma sp. Larval Contracaecum sp. also had a positive correlation with host length and intensity in the Chilean hake (George-Nascimento 1996).
The abundance of the copepod E. antarctica showed a significant positive correlation and an increased prevalence with increasing host length. This could be due to larger toothfish having a larger attachment area for the copepods. The monogenean Pseudobenedenia dissostichi also showed a significant positive correlation (P<0.01, r=0.931) between host length and abundance. The prevalence also increased with increasing length up to 40 cm total length but decreased after this. The increase in available surface area in larger fish would also explain the increase in abundance with host length.
The nematode Hysterothylacium sp. increased in abundance and prevalence with increasing host length, and this pattern could also be explained by an increase in host internal surface area for the parasites. The nematode Anisakis sp. did not show a significant correlation of abundance with increasing fish length but did show a pattern of increasing prevalence with increasing fish length. Anisakis spp. use euphausiids as the first intermediate hosts and squid and fish as second intermediate hosts. Their larvae can be passed onto other fish and squids without further moults; these squid and fish, therefore, act as paratenic hosts. Adult Anisakis spp. are parasites of pinnipeds and cetaceans (Anderson 2000). D. eleginoides will accumulate increasing numbers of Anisakis sp. through feeding on infected euphausiids and, later, by feeding on fish. Increasing intensity of Anisakis sp. with increasing host length was also noted in the Chilean hake (Carvajal et al. 1979). The digenean G. physidis and the monogenean N. georgianus did not show significant correlations of abundance with increasing host length. Although N. georgianus showed a pattern of increasing prevalence with length, it is likely that older/larger toothfish are more suitable hosts for this parasite and this is highlighted by the high range of intensities encountered in adult fish around the Subantarctic (Table 2).
Cysts of unknown aetiology found on the gill filaments of D. eleginoides also showed a significant positive correlation of abundance and an increase in prevalence with increasing host length. CUEs are quite common in D. eleginoides throughout its range and may be the so-called “tumour of attachment” of N. georgianus on the gill filaments. There was a positive correlation (P<0.01, r=0.626) between CUE numbers and N. georgianus numbers in the Prince Edward Island sample.
A number of parasite species seemed to be specific to certain Antarctic areas. The heaviest infections of Tetraphyllidean plerocercoids occur in immature toothfish around Shag Rocks, which may be related to their diet. Microsporidean species 1 and Neolebouria antarctica infected adult toothfish only at South Georgia. Gaevskaya et al. (1990) found the latter species only around South Georgia and not around the Ob and Lena Banks. Stenakron sp. and A. megarhynchus were found only around Prince Edward Island at relatively high prevalences of 26 and 45%, respectively. Lecithophyllum champsocephali was restricted to Heard, Macquarie and Prince Edward Islands. Sørensen’s similarity index illustrated that the parasite faunas of D. eleginoides were similar around the Subantarctic but showed greater distances in the Ross Sea. The greatest similarities were between Prince Edward, Heard, and Macquarie Islands. Smith (1986) concluded that no index based on presence–absence data was entirely satisfactory, but Sørensen’s was the best of those considered.
There may a gradual decrease in diversity eastwards around the Antarctic. However, larger sample sizes will be required to examine this in greater detail.
References
Anderson RC (2000) Nematode parasites of vertebrates: their development and transmission. CAB International, Wallingford
Andriashev AP (1964) On the composition and origin of the Antarctic pelagic fish fauna. In: Carrick R, Holdgate M, Prevost J (eds) Biologie Antarctique. Hermann, Paris, pp 271–272
Arana P, Vega R (1999) Exploratory fishing for Dissostichus spp. in the Antarctic region (subareas 48.1, 48.2 and 88.3). CCAMLR Sci 6:1–7
Arkhipkin A, Brickle P, Laptikhovsky V (2003) Variations in the diet of the Patagonian toothfish with size, depth and season around the Falkland Islands. J Fish Biol 63:428–441
Arthur JR, Albert E (1993) Use of parasites for separating stocks of Greenland halibut (Reinhardtius hippoglossoides) in the Canadian northwest Atlantic. Can J Fish Aquat Sci 50:2175–2181
Brickle P, Kalavati C, MacKenzie K (2001) Two new species of myxozoan parasites (Myxosporea, Bivalvulida) from toothfish Dissostichus eleginoides Smitt, 1898 (Pisces, Nototheniidae). Acta Parasitol 46:250–253
Bush AO, Lafferty KD, Lotz JM, Shostak AW (1997) Parasitology meets ecology on its own terms: Margolis et al. revisited. J Parasitol 83:575–583
Carvajal J, Cattan PE, Castillo C, Schatte P (1979) Larval anisakids and other helminths in the hake Merluccius gayi (Guichenot) from Chile. J Fish Biol 15:671–677
des Clers S (1991) Functional relationship between sealworm (Pseudoterranova decipiens, Nematoda, Ascaridoidea) burden and host size in Atlantic cod (Gadus morhua). P Roy Soc Lon Ser B 245:85–89
Duhamel G (1987) Distribution and abundance of fish on the Kerguelen Islands shelf. In: Kullander OS, Fernholm B (eds) Swedish Museum of Natural History, Stockholm, pp 397–403
Duhamel G (1991) Biologie et exploitation de Dissostichus eleginoides autour des iles Kerguelen (Division 58.5.1). (SC-CAMLR-SSP/8), 85–106. Hobart, Australia, CCAMLR
Duhamel G, Pletikosic M (1983) Donnes biologiques sur les Nototheniidae des Îles Crozet. Cybium 7:43–57
Eastman JT (1993) Antarctic fish biology: evolution in a unique environment. Academic, London
Elliot JM (1971) Some methods for the statistical analysis of samples of benthic invertebrates. Freshwater Biol Ass Sci Publ 25:1–144
Gaevskaya AV, Kovaleva AA (1976) Trematode fauna of certain mass species of fish from the south-west Atlantic (In Russian). Trudy AtlantNIRO 60:3–14
Gaevskaya AV, Kovaleva AA (1978) Materials on fish trematodes in the South-Western Atlantic (In Russian). Vestnik Zoologii 3:60–66
Gaevskaya AV, Kovaleva AA, Parukhin AM (1990) Peculiarities and formation of the parasitofauna of the Patagonian toothfish Dissostichus eleginoides (In Russian). Biol Morya 4:23–28
George-Nascimento M (1996) Populations and assembalges of parasites in hake, Merluccius gayi, from the southern Pacific Ocean: stock implications. J Fish Biol 48:557–568
Hart TJ (1946) Report on trawling surveys on the Patagonian continental shelf. Dis Rep 23:223–408
Hochberg FG (1990) Diseases of mollusca: Cephalopoda. In: Kinne O (eds) Diseases of marine animals, vol III. Cephalopoda to Urochordata. Biologisches Anstalt Helgoland, Hamburg, pp 47–227
Holloway HL, Spence JA (1980) Ecology of animal parasites in McMurdo sound, Antarctica. Com Physiol Ecol 5:262–284
Kock K-H, Duhamel G, Hureau JC (1985) Biology and status of exploited Antarctic fish stocks. A review. Biomass Sci Ser 6:1–143
Meissner EE, Zaostrovtsev AV (1983) New data on the distribution of Dissostichus eleginoides Smitt. Biol Morya 4:66–67
Moser M, Cowen R K (1991) The effects of periodic europhication on parasitism and stock identification of Trematomus bernacchii (Pisces: Nototheniidae) in McMurdo Sound, Antaractic. J Parasitol 77:551–556
Norman JR (1937) Coast fishes Part II. The Patagonian region. Dis Rep 16:1–150
Norman JR (1938) Coast fishes Part III. The Antarctic zone. Dis Rep 19:1–104
Oyarzun C, Campos PW (1987) Dissostichus eleginoides Smitt, 1898; concideraciones sobre su determinacion taxonomica e implicancias biogeographicas (Pisces, Perciformes, Nototheniidae). Rev Biol Mar 23:173- 192
Palm HW, Reimann N, Spindler M, Poltz J (1998) The role of the rock cod Notothenia coriiceps Richardson, 1844 in the life-cycle of Antarctic parasites. Polar Biol 19:399–406
Palm H (1999) Ecology of Pseudoterranova decipiens (Krabbe, 1878) (Nematoda: Anisakidae) from Antarctic waters. Parasitol Res 85:638–646
Parukhin AM, Lyadov VN (1982) Helminth fauna of food Nototheniidae fishes from Kerguelen region (In Russian). Ekologya Morya Kiev 10:49–57
Poulin R (2000) Variation in intraspecific relationship between fish length and intensity of parasitic infection: biological and statistical causes. J Fish Biol 56:123–137
Rodjuk GN (1984) New representatives of the genus Metechinorhynchus (Acanthocephala), parasites of fishes of the western Atlantic (In Russian). Zoologicheskiy Zhurnal 63:1893–1896
Rodjuk GN (1986) New species of acanthocephala of the genus Echinorhynchus (Echinorhynchidae) from the southwestern Atlantic (In Russian). Parazitologiya 20:224–227
Rodriguez L, George-Nascimento M (1996) The metazoan parasite fauna of the Patagonia toothfish Dissostichus eleginoides Smitt, 1898 (Pisces: Nototheniidae) off central Chile: taxonomic, ecological and zoogeographical aspects. Rev Chil Hist Nat 69:21–33
Rohde K (1999) Latitudinal gradients in species diversity and Papoprt’s rule revisited: a review of recent work and what can parasites teach us about the causes of gradients? Ecography 22:593–613
Smith B (1986) Evaluation of different similarity indices applied to data from the Rothamsted insect survey. University of York, York
Smith P, McVeagh M (2000) Allozyme and microsatellite DNA markers of toothfish population structure in the Southern Ocean. J Fish Biol 57:72–83
Sørensen T (1948) A method of establishing groups of equal amplitude in plant sociology based on similarity of species content and its application to analyses of the vegetation on Danish commons. Biol Skr (K Danske Vidensk Selsk NS) 5:1–34
Sosinski J, Kuranty J (1979) The Scotia Sea—a new fishing ground for the Polish Fishery. Technika Gospodarka Morska 29:12–15
Wägele JW (1988) Apects of the life cycle of the Antarctic fish parasite Gnathia-calva Vanhoffen (Crustacea, Isopoda). Polar Biol 8:287–291
Walter T, Palm HW, Piepiorka S, Rückert S (2002) Parasites of the Antarctic rattail Macrourus whitsoni (Regan, 1913) (Macrouridae, Gadiformes). Polar Biol 25:633–641
Yukhov VL (1982) Antarctic toothfish (In Russian). Nauka, Moscow
Zdzitowiecki K (1991) Antarctic Acanthocephala. Koeltz Scientific Books, Königstein, Germany
Zdzitowiecki K (1997) Antarctic Digenea, parasites of fishes. Koeltz Scientific Books, Königstein, Germany
Acknowledgements
We thank the Government of South Georgia and the South Sandwich Islands, British Antarctic Survey and the Marine Resources Assessment Group for providing samples from Shag Rock and South Georgia. We are also indebted to Dr. Dick Williams of the Australian Antarctic Division for providing samples from off Heard and Macquarie Islands and to Dr. Peter Smith of NIWA for providing samples from the Ross Sea. We thank Martin Purvis and Linda Staverees of Marine and Coastal Management, Cape Town for providing samples from Prince Edward Island and for providing laboratory space. The Shackleton Scholarship fund provided funds for a flight to Cape Town, for PB for which we are grateful. Consolidated Fisheries Ltd was the driving force in the initial stages of this project. We are grateful to the Director of Fisheries (Falkland Islands Government), John Barton, for supporting this work and to Professor Chris Secombes for providing laboratory facilities at the School of Biological Sciences, University of Aberdeen. We also thank Dr. Les Chappell for his help in Aberdeen. Finally, we thank the three anonymous referees that helped to improve the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Brickle, P., MacKenzie, K. & Pike, A. Parasites of the Patagonian toothfish, Dissostichus eleginoides Smitt 1898, in different parts of the Subantarctic. Polar Biol 28, 663–671 (2005). https://doi.org/10.1007/s00300-005-0737-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-005-0737-2