
Abstract
The morphology, biology and geographic distribution of Hypsibius zetlandicus (Murray 1907) are considered. Scanning electron microscopy (SEM) and/or light microscopy (LM) analyses have been carried out on H. zetlandicus and the type species of Hypsibius: Hypsibius dujardini (Doyère 1840), with particular emphasis on the buccal–pharyngeal apparatuses. Some unusual characteristics of this apparatus in H. zetlandicus lead us to the erection of the new genus Borealibius, to which H. zetlandicus (Borealibius zetlandicus comb. n.) is transferred. In the light of new discoveries of this species from polar, sub-polar and alpine regions, and based on the available bibliographic references, we hypothesize a boreo–alpine distribution for this species. The presence of traits that are unusual and rare (in other tardigrade species) have been observed whilst analyzing the reproductive biology and ecology of B. zetlandicus (i.e., the presence of hermaphroditism, parental care and the colonization of very different substrates).
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The ability of most non-marine tardigrade species to survive in extreme environments and conditions using cryptobiosis, together with passive dispersal and the evolution of parthenogenesis and self-fertilization (this last a rare phenomenon in tardigrades), theoretically allows the colonization of new and remote territories from a single individual (Bertolani 2001). Despite this theoretical possibility, a very high number of tardigrade species have limited geographical distributions and are adapted to restricted microenvironmental conditions. Pilato (1979) emphasized this apparent disparity based on the available literature and suggested an interpretation related to the biology of tardigrades. Based on these statements, several authors have continued to point out the biogeographic importance of tardigrades (e.g., Binda and Pilato 1973; Pilato 1979; Bertolani et al. 1993; McInnes 1994; Pugh and McInnes 1998; Pilato and Binda 2001). To improve the knowledge on this topic, our samplings and materials furnished by colleagues (see Acknowledgements) gave us the opportunity to examine several specimens of Hypsibius zetlandicus (Murray 1907) coming from different regions of the world. Material from Greenland, Faroe Islands and Alps (Italy), together with Russian material from Biserov’s collection, allowed us to study in more depth the geographic distribution, biology and morphology of this species. Little had been written on the biology of this species, with only scant information on the possibility of eggs developing within the adult animals (Murray 1907a; Biserov 1988).
Materials and methods
Materials examined
We examined specimens of H. zetlandicus from Greenland (pond sediment at Disko Island) and from Italian Alps (sediment of Nambino Lake, 1,800 m a.s.l., Madonna di Campiglio, Trento; submerged moss of a spring, 2,270 m a.s.l., Valle Pian Venezia, Trento, collected in March, July and November 2004). Moreover, we examined the specimens from a small lake at Naryan-Mar, Karelia, European Russia (from Biserov’s collection held by the Museum of Natural History of Verona) and specimens collected by Birna V. Trygvadottir from three localities of Faroe Island: Sornfelli locality (pond sediment) and Saksun locality (colony of cyanobacteria—Nostoc sp.) both on Streymoy Island and Fuglafjorour locality (sediment of temporary pond) on Eysturoy Island.
For taxonomic comparison, we examined specimens of Hypsibius dujardini (Doyère 1840), the type species of the genus Hypsibius, from a cultured strain present in the laboratory of the University of Modena and Reggio Emilia (the cultured strain derived from animals given by Prof. Bob Goldstein of the Department of Biology, University of North Carolina, USA), and specimens of the same species from a Sicilian stream (Torrente Cutò, Nebrods Mts., Italy).
Specimens, mounted in polyvinyl lactophenol or in Faure-Berlese fluid, of the two species considered and in vivo specimens of H. zetlandicus have been observed by light microscopy (LM) up to maximum magnification.
For scanning electron microscopy (SEM) observations on H. zetlandicus, specimens were obtained from the population of Valle Pian Venezia; buccal–pharyngeal apparatuses of H. dujardini were obtained from specimens of the cultured strain cited above. A Philips SEM XL 40 was used, available at the ‘Centro Interdipartimentale Grandi Strumenti’ of the University of Modena and Reggio Emilia.
We examined buccal–pharyngeal apparatuses of H. zetlandicus and H. dujardini prepared for SEM, according to the method recently suggested by Eibye-Jacobsen (2001).
The pt index used is the percentage ratio between the length of a structure and the length of the buccal tube measured from the anterior margin of the dorsal transversal ridge of the buccal armature to the base of the pharyngeal apophyses (Pilato 1981).
For remarks on the reproductive biology, specimens of H. zetlandicus from Valle Pian Venezia population have been fixed in Carnoy (methanol/acetic acid 3:1), stained with acetic carmine, subsequently mounted in Faure-Berlese’s mounting medium and observed by LM. In vivo observations have been carried out on Italian material from Valle Pian Venezia and on Greenlander material.
Results and discussion
Morphological and taxonomic results
H. zetlandicus
The cuticle is wrinkled (Fig. 1a); eyespot is present. The buccal armature has three dorsal and three ventral transversal ridges: the dorso-lateral and the ventro-lateral ridges at each extremity are prolonged into an acuminate tooth. The buccal tube (Figs. 1b–d, 2a, b, 4a, b) is narrow and rigid without ventral lamina. The walls of the anterior portion of the buccal tube are thick, and their dorsal and ventral caudal margins form a well-developed transversal crest (Figs. 1c, d, 2a–d, 4a, b) with very pronounced dorsal and ventral apophyses for the insertion of the stylet muscles (AISMs); dimensions and shape of these structures represent the main peculiarity of this species. They are almost cylindrical in shape with a large rounded or bilobated projection pointing caudally and diverging from the buccal tube walls (Figs. 1b, c, 2a–d, 4a, b). The anterior portions of both apophyses are joined to the buccal tube wall by two cuticular laminae that, extending laterally, participate to form the stylet sheaths (i.e., the canals where the terminal portion of the stylets settle) (Fig. 2a, c, d, 4a, b). Peribuccal lamellae and peribuccal papulae are absent; pharyngeal bulb has two macroplacoids, both with a marked central narrowing (Figs. 1c, d, 2a, b, 4a, b); microplacoid is absent.

H. zetlandicus (LM). a Detail of the cuticular sculpture. b Buccal–pharyngeal apparatus in dorsal view. c Buccal–pharyngeal apparatus in lateral view. d Buccal armature in which are visible some teeth. e Claws of the second and third pairs of legs. f Claws of the fourth pair of legs; a short cuticular bar is visible between the claws of this pair of legs. Scale bar=20 μm

H. zetlandicus (SEM). a, b Buccal–pharyngeal apparatuses in dorsal (a) and in lateral view (b). c, d Anterior portions of the buccal tube (detail of Fig. 2a, b, respectively), black and white arrows indicate the stylet sheath. E. Stylet furca. Scale bars=5 μm
The stylet furcae (Figs. 1c, d, 2a, b, e, 4a, b) have an unusual aspect, due to the fact that the postero-lateral processes are more robust than they are in other genera of Hypsibiidae (Fig. 3a, c). The extremity of each process is thickened and swollen.

H. dujardini (SEM). a Buccal–pharyngeal apparatus (dorsal view). b Anterior portion of the buccal tube (detail of Fig. 3a); note that the postero-lateral processes of the AISMs extend laterally and participate to form the stylet sheaths (arrow). c Stylet furca. Scale bars=2 μm
Claws are of Hypsibius type and well developed (Fig. 1e, f) with accessory points on the main branches; lunules are absent; there are no cuticular bars on the first three pairs of legs; a short cuticular bar is present between the claws of the fourth pair of legs (Fig. 1f).
In a specimen from Nambino Lake (Madonna di Campiglio, Italy), the body length is 360 μm. The buccal tube is 52.3 μm long and 4.1 μm wide (pt=7.8); stylet supports are inserted on the buccal tube wall at 63.9 % of its length (pt=63.9); The entire macroplacoid row is 19.1 μm long (pt=36.5), first macroplacoid 10.2 μm (pt=19.5), second macroplacoid 8.5 μm long (pt=16.2); external claws on the second pair of legs are 22.1 μm long (pt=42.2), internal claws 17.2 μm long (pt=32.9).
H. dujardini
The specimens of H. dujardini show both AISMs that, in lateral view, appear as “semilunar hooks” (Fig. 4d), and in dorsal or ventral view (Fig. 3a, b, 4c) appear as longitudinal processes that run downwards from the anterior extremity of the buccal tube and end with two postero-lateral processes that extend laterally and participate to form the stylet sheaths. The stylet furca (Fig. 3c) has thinner and longer postero-lateral processes.

a, bH.zetlandicus. A, Buccal–pharyngeal apparatus in dorsal (a) and lateral (b) views. c, dH. dujardini. Buccal–pharyngeal apparatus in dorsal (c) and lateral (d) views. Scale bar=20 μm
Phylogenetic and taxonomic evaluations
As represented by Marcus (1929), the anterior portions of the stylet protractor muscles are attached to the AISMs while the posterior portions are attached to the stylet furcae. Therefore, these two sclerified structures are related to each other because they are connected by the same muscular bundles. Likewise, H. zetlandicus (Hypsibiidae), the Macrobiotidae Adorybiotus granulatus (Richters 1903) and Richtersius coronifer (Richters 1903) show wide AISMs and wide stylet furcae. It is reasonable to suppose that the larger sizes of these structures in these species are related to a larger muscular structure associated to them. The adaptive gain of these structures can be related to the higher power of penetration of the stylets during the phase of food assumption.
As reported by Pilato (1987) and Pilato and Binda (1987), the shape of the AISMs and the shape of the stylet furcae are constant within the genera (sometimes also in different genera), showing in general a reduced tendency for variation; therefore, a marked difference affecting these structures can be considered a significant indicator of phylogenetic divergence (Pilato and Binda 1997).
The AISMs of H. zetlandicus are considerably different from those of H. dujardini and from any other species of the genus Hypsibius and also of the entire family Hypsibiidae. The only other known case with large AISMs within the Hypsibiidae is represented by Isohypsibius malawiensis. Jørgensen (2001) described as having a drop-shaped cuticular swelling. Nonetheless, these AISMs are quite different from those of H. zetlandicus. It is interesting to note that Murray described H. zetlandicus in 1907, when not only the shape, but also the existence of the AISMs was not considered as taxonomic detail; however, Murray (1907a) in Fig. 24a of Plate IV drew one large apophysis with a rounded extremity exactly as it appears in our specimens.
According to a criterion of in-group commonality: “a plesiomorphous character state is more likely to be widespread within a monophyletic taxon than is any one apomorphous character state” (de Jong 1980). Therefore, the different shape of AISMs with respect to H. zetlandicus (i.e., without a large lobed projection), and in particular the absence of a transversal crest in the caudal portion of the external walls of the buccal cavity in Hypsibius and in all other genera of Hypsiibiidae, leads to suppose that this last situation represents the plesiomorphic condition. For a parsimonious criterion, it is to hypothesize that the large AISMs with a large rounded or lobed projection and the presence of a dorsal and a ventral transversal crest in the caudal portion of the buccal cavity may have been formed once, and not lost independently many times in different evolutionary lines of the family. Therefore, these characters present in H. zetlandicus can be considered autoapomorphies, and they justify, in our opinion, the transfer of this species to a new genus we named Borealibius. This has to be considered closely related to Hypsibius, due to the very similar claw morphology.
Borealibius, gen. n
Diagnosis: Claws of Hypsibius type; buccal–pharyngeal apparatus with rigid buccal tube without ventral lamina; in their caudal margins, the walls of the anterior portion of the buccal tube form well-developed transversal crests with very pronounced AISMs, almost cylindrical in shape with a large rounded or bilobed projection pointing caudally and diverging from the buccal tube walls; peribuccal lamellae and peribuccal papulae absent; pharyngeal apophyses and placoids present.
Etymology: The name Borealibius refers to the distribution of the species only in localities of the Boreal hemisphere.
Type species: Macrobiotus zetlandicus (Murray 1907).
Composition: Borealibius zetlandicus, comb. n. is the only known species of the genus.
Three specimens of B. zetlandicus from Disko Island (Greenland), eight specimens from Nambino Lake and 112 specimens from Valle Pian Venezia are deposited in Bertolani’s collection (Dipartimento di Biologia Animale, Università di Modena e Reggio Emilia, Modena, Italy); 15 specimens from Karelia are deposited in Biserov’s collection (Museo Civico di Storia Naturale, Verona, Italy); one specimen from Karelia and five specimens from Italy (Nambino Lake, Madonna di Campiglio, Trento) are deposited in the collection of Binda and Pilato (Museo del Dipartimento di Biologia Animale “Marcello La Greca”, Università di Catania, Italy); six specimens from three localities of Faroe Island (Sornfelli , Saksun and Fuglafjorour) collected by Birna V. Trygvadottir are deposited in the collection of the Department of Invertebrate Zoology, Zoological Museum, University of Copenhagen (Copenhagen, Denmark).
Reproductive biology of B. zetlandicus
Most specimens from Valle Pian Venezia exhibited a small gonad with a single lateral gonoduct connected to the rectum (as in eutardigrade gonochoristic females and in hermaphrodites). In all samples (collected in March, July and November), specimens with oocytes in advanced stage of maturation were never observed. Within the gonad, we observed spermatids and few spermatozoa mixed with oocytes in pre-vitellogenesis, which were never separated by morphological barriers (Fig. 5). This type of gonad was also observed in animals carrying the exuvium containing eggs or newborns (see below; Fig. 5c–e). In most animals, a cluster of spermatozoa was present in the posterior part of the gonad, close to the beginning of the gonoduct (Fig. 5e).

B. zetlandicus comb. n. (LM). a Ovotestis with evident spermatids (arrow head) and spermatozoa (full arrow). b Ovotestis with previtellogenetic oocyte (empty arrow) and spermatozoa. c Old exuvium containing an animal with small gonad (plus), an embryonated egg (filled star) and a newborn (asterisk). d Detail of C, small ovotestis with spermatozoa and previtellogenetic oocytes. e Ovotestis with previtellogenetic oocytes and spermatozoa, to note the group of sperms in the posterior portion of the gonad (within the circle). Scale bars: A, B=10 μm, C=45 μm, D, E=15 μm
Animals laid from 3 to 12 smooth eggs within the exuvia. Although most observations of exuvia containing eggs were not associated with adult animals, occasionally adult animals were found “carrying” exuvia with eggs (even near full-term, embryonate eggs and hatchings) (a specimen from Greenland and several specimens from Valle Pian Venezia). Under laboratory conditions (at 13°C over several weeks) a number of adult animals were observed to molt but remained within an old cuticle along side embryonated eggs and/or 1–3 newborns (Fig. 5c).
B. zetlandicus, or at least the population of Valle Pian Venezia (Italy), is to be considered hermaphroditic for the presence of an ovotestis similar to that of other hermaphroditic tardigrade species (Rebecchi et al. 2000). Hermaphroditic species and/or populations are not common in tardigrades, indicating that hermaphroditism is the less frequent sexual condition. The new finding of a hermaphroditic species (or population) confirms that in tardigrades this condition developed independently in different evolutionary lines. Within the phylum, only one species of the class Heterotardigrada (family Halechiniscidae) is hermaphroditic (R.M. Kristensen, personal communication in Bertolani 1987), whereas in Eutardigrada hermaphroditism is a little more frequent, though sporadic. With B. zetlandicus, nine other tardigrade species are hermaphroditic and belong to five different families: Macrobiotidae, Eohypsibiidae, Hypsibiidae, Calohypsibiidae and Necopinatidae (see Bertolani 1987, 2001; Rebecchi et al. 2000).
In two tardigrade species, Isohypsibius monoicus (Bertolani 1981) and Macrobiotus joannae (Pilato and Binda 1983), self-fertilization certainly occurs (Bertolani and Rebecchi 1999). We cannot confirm whether this is true for B. zetlandicus, even though spermatozoa are never separated from the oocytes by morphological barriers.
The exuvia containing eggs at the end of the development carried by the animals indicate the presence of a form of parental care by B. zetlandicus. This phenomenon is also uncommon in tardigrade species; with only Isohypsibius annulatus (Murray 1905) and in two species of the genus Pseudobiotus having been observed showing similar, but not an identical, behavior (see Bertolani and Rebecchi 1999). In the original description of B. zetlandicus, Murray (1907a) reports: “Living youngs have been seen in the body of the parent ... some malformation has prevented the deposition of the eggs, which were retained till hatched”, whereas Biserov (1988) reports that, within the ovary of some females, embryonated eggs were present due to an abnormal development or to viviparity. Retention of fully developed embryos within the ovary has been recently reported for one specimen of Macrobiotus richtersi (Murray 1911) of a cultured population (Hohberg and Greven 2005), but we do not have sufficient data to decide if this phenomenon is physiological or not.
Our findings on B. zetlandicus, in particular, the newborns still within the old cuticle of the parent indicate that some “parental care” is possible, even though this is limited and not always present. Moreover, these new data suggest that Murray (1907a) and Biserov (1988) misinterpreted their observations of this condition in B. zetlandicus, though this is a condition not constant, and uncommon or absent in other tardigrades.
Ecology and biogeography
B. zetlandicus has been found in marine samples from the Barents Sea (Biserov 1999), freshwater sediments (Murray 1907a; Biserov 1996, 1999, present study), submerged mosses (present study) and Sphagnum sp. (Biserov 1991), terrestrial substrates, such as moss (Rudescu 1964; Morgan 1980), mosses on soil (Biserov 1991), soil (animals and cysts; Stark and Kristensen 1999) and on colony of cyanobacteria (present study). The wide range of habitats available for this species represents an exception for a tardigrade species, whose occurrence is more usually restricted to only one or a few substrates.
The species was originally described as M. zetlandicus by Murray (1907a) with specimens from Fort Augustus (Scotland), Shetland Islands (Zetland being an alternative spelling and the etymology of the species name) and Spitsbergen and later attributed to the genus Hypsibius by Thulin (1928). Van der Land (1966) defined a lectotype from the original slides and correctly indicated Fort Augustus (bog pool) as the Locus typicus because, according to Murray (1907a), that locality represented the oldest site from which this species had been found.
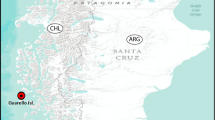
In addition to the L. typicus, B. zetlandicus has also been found in Arctic regions of Russia (Malyi Vudjavr in Murmansk district, Naryan-Mar, Karelia, Vorkuta, Novaya Zemlya and Taimyr peninsula) by Biserov (1988, 1991, 1996, 1999), in Spitsbergen (Svalbard) by Murray (1907a, b) and Greenland (Disko Island) by Stark and Kristensen (1999) and Hansen and Katholm (2002), and in sub-polar areas such as the Shetland Islands by Murray (1907a) and Iceland by Morgan (1980). Under the 60° parallel, this species has been found in the European states of Finland (Nederström 1919), Scotland (Murray 1907a; Morgan 1974; van der Land 1966), Slovakia (Bartoš 1935, 1939, in a mountain region between 1,800 and 2,400 m a.s.l.) and Romania (Rudescu 1964, in mountain region) (Fig. 6). The discovery of B. zetlandicus from the Faroe Islands and Italy (in mountain regions between 1,800 and 2,300 m a.s.l.) are new records for these countries. Morgan (1980) reported an occurrence of B. zetlandicus in Greenland but did not report who found it, or where and when the species was found.

Distribution of B. zetlandicus, comb. n.: plus = Murray (1907a, b), cross = Nederström (1919), filled diamond = Bartoš (1935, 1939), filled half circle = Rudescu (1964), inverted triangle = van der Land (1966), filled star = Morgan (1974, 1980), filled circle = Biserov (1988, 1991, 1996, 1999), filled triangle = Hansen and Katholm (2002) and filled square = present study
The rich tardigrade biodiversity in Disko Island (more than 100 species) (Heide-Jørgensen and Kristensen 1998, 1999; Stark and Kristensen 1999; Hansen and Katholm 2002) emphasizes the accumulating evidence that these extreme environments can be colonized by a very high numbers of well-adapted species.
The discontinuous distribution of this species, present in polar, sub-polar and alpine (mountain above 1,800 m a.s.l.) regions, clearly indicates a boreo–alpine distribution. This distribution reflects the palaeo-climatic events and is related to ecological needs of B. zetlandicus, particularly cold climates. In non-polar regions this species can be considered a “glacial relict”.
References
Bartoš E (1935) Tardigrada Vysokýchtater. Příroda 28:1–4
Bartoš E (1939) Die tardigraden der Tschechoslowakischen Republik. Zool Anz 125:138–142
Bertolani R (1987) Sexuality, reproduction and propagation in tardigrades. In: Bertolani R (ed) Biology of tardigrades, selected symposia and monographs U.Z.I., vol. 1. Mucchi, Modena, pp 93–107
Bertolani R (2001) Evolution of the reproductive mechanisms in tardigrades—a review. Zool Anz 240:247–252
Bertolani R, Rebecchi L (1999) Tardigrada. In: Knobil E, Neill JD (eds) Encyclopedia of reproduction, vol. 4. Academic, San Diego, pp 703–717
Bertolani R, Guidetti R, Rebecchi L (1993) Tardigradi dell’Appennino umbro-marchigiano. Biogeographia 17:223–245
Binda MG, Pilato G (1973) Le attuali conoscenze sulla fauna tardigradologica di alcune piccole isole circumsiciliane. Lav Soc Ital Biogeografia N S 3:1–7
Biserov VI (1988) Limnetic tardigrades of some areas in the USSR (In Russian). Zool Zh 67:1798–1811
Biserov VI (1991) An annotated list of Tardigrada from European Russia. Zool Jahrb Syst 118:193–216
Biserov VI (1996) Tardigrades of the Taimyr peninsula with descriptions of two new species. Zool J Linn Soc 116:215–237
Biserov VI (1999) A review of the Tardigrada from Novaya Zemlya, with descriptions of three new species, and an evaluation of the environment in this region. Zool Anz 238:169–182
Eibye-Jacobsen J (2001) A new method for making SEM preparations of the tardigrade buccopharyngeal apparatus. Zool Anz 240:309–321
Hansen JG, Katholm AK (2002) A study of the genus Amphibolus from Disko Island with special attention on the life cycle of Amphibolus nebulosus (Eutardigrada: Eohypsibiidae). In: Hansen JG (ed) Arctic biology field course quqertarsuaq, 2002. Zoological Museum, University of Copenhagen, H.C.Ø. TRYK, Copenhagen, pp 129–163
Heide-Jørgensen HS, Kristensen RM (1998) Puilassoq, Disko øens varmeste homoterme kilde. Arctic biology course, Disko Island, Greenland, pp 219–231
Heide-Jørgensen HS, Kristensen RM (1999) Puilassoq, the warmest homothermal spring of Disko Island. Ber Polarforsch 99:32–43
Hohberg K, Greven H (2005) Retention of embryonated eggs in parthenogenetic Macrobiotus richtersi J. Murray, 1911 (Eutardigrada). Zool Anz 243:211–213
de Jong R (1980) Some tools for evolutionary and phylogenetic studies. Z zool Syst Evol-forsch 18:1–23
Jørgensen A (2001) Graphical presentation of the African tardigrade fauna using GIS with the description of Isohypsibius malawiensis sp. n. (Eutardigrada: Hypsibiidae) from Lake Malawi. Zool Anz 240:441–449
Land J van der (1966) The tardigrada of the Scottish lake survey described as new species by James Murray. Proc R Soc Edinburgh, Section B 69:298–320
Marcus E (1929) Tardigrada. In: Bronns HG (ed) Klassen und Ordnungen des Tierreichs, vol. 5. Leipzig, pp 1–608
McInnes SJ (1994) Zoogeographic distribution of terrestrial/freshwater tardigrades from current literature. J Nat Hist 28:257–352
Morgan CI (1974) Studies on the biology of Tardigrades. PhD thesis, University of Wales, UK
Morgan CI (1980) A systematic survey of tardigrada from Iceland. Acta Nat Islandica 27:1–25
Murray J (1907a) Scottish tardigrada collected by the lake survey. Trans R Soc Edinburgh 45:641–668
Murray J (1907b) Arctic tardigrada collected by Wm. S. Bruce. Trans R Soc Edinburgh 45:669–681
Nederström P (1919) Die bis jetzt aus Finland bekannten Tardigraden. Acta Soc Fauna Flora Fennica 46:25
Pilato G (1979) Correlations between cryptobiosis and other biological characteristics in some soil animals. Boll Zool 46:319–332
Pilato G (1981) Analisi di nuovi caratteri nello studio degli Eutardigradi. Animalia 8:51–57
Pilato G (1987) Revision of the genus Diphascon Plate, 1889, with remarks on the subfamily Itaquasconinae (Eutardigrada, Hypsibiidae). In: Bertolani R (ed) Biology of tardigrades, Selected symposia and monographs, U.Z.I., vol. 1. Mucchi, Modena, pp 337–357
Pilato G, Binda MG (1987) Richtersia, nuovo genere di Macrobiotidae, e nuova definizione di Adorybiotus Maucci e Ramazzotti, 1981 (Eutardigrada). Animalia 14:147–152
Pilato G, Binda MG (1997) Acutuncus, a new genus of Hypsibiidae (Eutardigradi). Entomol Mitt Zool Mus Hamburg 12:159–162
Pilato G, Binda MG (2001) Biogeography and limno-terrestrial tardigrades: are they truly incompatible binomials? Zool Anz 240:511–516
Pugh PJA, McInnes JS (1998) The origin of Arctic terrestrial and freshwater tardigrades. Polar Biol 19:177–182
Rebecchi L, Guidi A, Bertolani R (2000) Maturative pattern of the ovotestis in two hermaphrodite species of eutardigrades. Invertebr Reprod Dev 37:25–34
Rudescu L (1964) Tardigrada. Acad Rep Pop Romîne 4:350–353
Stark S, Kristensen RM (1999) Tardigrades in the soil of Greenland. Ber Polarforsch 330:44-63
Thulin G (1928) Über die Phylogenie und das System der Tardigraden. Hereditas 11:207–266
Acknowledgement
The authors thank Clark Beasley for the English revision of the paper. Luisella Previdi for collecting the Nambino Lake sample, Valeria Lencioni and Bruno Maiolini for collecting Valle Pian Venezia samples, Birna V. Trygvadottir for sending us Faroe Island specimens and the Museum of Natural History of Verona for giving us the opportunity to examine Biserov’s collection. This work was supported by grants from the University of Catania (Fondo Ricerca d’Ateneo) and by the University of Modena and Reggio (Fondo Ricerca d’Ateneo).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Pilato, G., Guidetti, R., Rebecchi, L. et al. Geonemy, ecology, reproductive biology and morphology of the tardigrade Hypsibius zetlandicus (Eutardigrada: Hypsibiidae) with erection of Borealibius gen. n.. Polar Biol 29, 595–603 (2006). https://doi.org/10.1007/s00300-005-0094-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-005-0094-1