
Abstract
Increased public awareness of the global significance of polar regions and opening of the Russian Arctic to foreign researchers have led to a pronounced intensification of benthic research in Arctic seas. The wealth of information gathered in these efforts has markedly enhanced our knowledge on the Arctic benthos. While some scientific concepts have been corroborated by the novel findings (e.g., low endemism and high faunistic affinity to northern Atlantic assemblages), other common notions need to be revised, particularly with regard to the often-cited differences between Arctic seas and the Southern Ocean. It has been demonstrated that benthos assemblages vary broadly in diversity between Arctic regions and that, hence, the idea of a consistently poor Arctic benthos—being in stark contrast to the rich Antarctic bottom fauna—is an undue overgeneralization. In terms of biogeographic diversity, both Arctic and Antarctic waters seem to be characterized by intermediate species richness. Levels of disturbance—a major ecological agent known to heavily affect benthic diversity and community structure—have been assumed to be relatively high in the Arctic but exceptionally low in the Southern Ocean. The discovery of the great role of iceberg scouring in Antarctic shelf ecosystems, which has largely been overlooked in the past, calls for a reconsideration of this notion. The novel data clearly demonstrate that there are marked differences in geographical and environmental setting, impact of fluvial run-off, pelagic production regime, strength of pelago–benthic coupling and, hence, food supply to the benthos among the various Arctic seas, impeding the large-scale generalization of local and regional findings. Field evidence points to the great significance of meso-scale features in hydrography and ice cover (marginal ice zones, polynyas, and gyres) as ‘hot spots’ of tight pelago–benthic coupling and, hence, high benthic biomass. In contrast, the importance of terrigenic organic matter discharged to the Arctic seas through fluvial run-off as an additional food source for the benthos is still under debate. Studies on the partitioning of energy flow through benthic communities strongly suggest that megafauna has to be adequately considered in overall benthic energy budgets and models of carbon cycling, particularly in Arctic shelf systems dominated by abundant echinoderm populations. Much progress has been made in the scientific exploration of the deep ice-covered Arctic Ocean. There is now evidence that it is one order of magnitude more productive than previously thought. Therefore, the significance of shelf–basin interactions, i.e., the importance of excess organic carbon exported from productive shelves to the deep ocean, is still debated and, hence, a major topic of on-going research. Another high-priority theme of current/future projects are the ecological consequences of the rapid warming in the Arctic. Higher water temperatures, increased fluvial run-off and reduced ice cover will give rise to severe ecosystem changes, propagating through all trophic levels. It is hypothesized that there would be a shift in the relative importance of marine biota in the overall carbon and energy flux, ultimately resulting in a switch from a ‘sea-ice algae–benthos’ to a ‘phytoplankton–zooplankton’ dominance.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Since the end of the 1980s, there has been a profound change in the significance that both the scientific community and the general public allocate to the polar regions. Now, it is generally accepted that both the Arctic and Antarctica are not only influenced by the same global processes that affect the entire Earth but they have also been recognized to heavily impact large-scale processes that in turn shape the global climate (Macdonald 1996; Clarke 2003). This is particularly true for the polar seas and their ice cover.
This paradigm shift and a remarkable intensification of scientific work in polar seas are interlinked by a positive feedback: the research results called for the change in the perception of Arctic and Antarctic areas, and this in turn stimulated further studies. Another important aspect contributing to the new bloom of polar marine research is the technical progress made in recent years. Powerful ice-breaking vessels being specifically devoted to research purposes, such as the German RV “Polarstern”, the British RV “James Clark Ross” and the USCGC “Healy”, became readily available to polar researchers and allowed for undertaking scientific expeditions that can go deeper into ice-covered regions than ever before. Last but not least, in case of the Arctic the scientific research was also significantly promoted by the revolution in the former Soviet Union starting in the late 1980s. This unexpected development, being intimately connected with the introduction of the novel policy of ‘glasnost’ (openness), was of truly global political significance. As a side effect, the Eurasian–Arctic waters off the vast northern Soviet territories, which account for nearly half of the Arctic seas but had effectively been barred to non-Soviet scientists during a decade-long period of isolationism, became open for international research (Piepenburg 1995).
Together, these developments sparked the launching of several multidisciplinary, often international research efforts since the late 1980s (Table 1), e.g., ‘The Northern North Atlantic: A changing environment’, a German collaborative research centre (SFB 313; Schäfer et al. 2001), the ‘Northeast Water Polynya Study’ (NEW; Hirche and Kattner 1994) and the ‘North Water Polynya Study’ (NOW; Deming et al. 2002), both performed within the framework of the ‘International Arctic Polynya Project’ (IAPP; Anonymous 1991), the ‘European Polarstern Study’ (Arctic EPOS; Rachor 1992), the Russian–German projects ‘System Laptev Sea’ (Thiede et al. 1999) and ‘Siberian River Run-Off’ (SIRRO; Stein et al. 2003), the Norwegian-led ‘Carbon flux and ecosystem feedback in the northern Barents Sea in an era of climate change’ project (CABANERA; Wassmann, personal communication), the ‘Western Arctic Shelf–Basin Interactions’ project (SBI; Grebmeier 2003), and the ‘Canadian Arctic Shelf Exchange Study’ (CASES; Fortier, personal communication). Within the frame of these multi-year projects, marine field studies have been carried out in various Eurasian–Arctic (Greenland, Barents, Kara, and Laptev Seas) and American–Arctic seas (Bering, Chukchi and Beaufort Seas, Baffin Bay). In the course of these investigations, a wealth of novel data has been collected, which allowed for extending our information base on Arctic benthos faunas considerably and, thus, for testing the general validity of common notions about the structure and functioning of high-latitude benthic systems.
A comprehensive review of the current knowledge on Arctic benthos in general would be clearly beyond the scope of this essay. Here, I will rather focus on the assemblages of ‘large’ benthic organisms (macro- and megabenthos), i.e., according to the well-established scientific usage (Gage and Tyler 1991), on those seafloor organisms that are large enough to be retained on sieves with a mesh size of 0.5 mm (macrobenthos, mostly infaunal) or to be visible in seabed images and/or to be caught by towed sampling gear (megabenthos, mostly epifaunal). Moreover, the extensive work recently performed in Arctic fjords, for example, at Spitsbergen (Wlodarska-Kowalczuk et al. 1998), especially in the Kongsfjord (Hop et al. 2002; Kendall et al. 2003; Wlodarska-Kowalczuk and Pearson 2004), is certainly not savored as it actually deserves to be, as I will largely concentrate on the findings reported from off-shore shelf and deep-sea environments. Finally, I will largely confine myself to deal with a few selected ecological aspects, for which recent research results have led to the necessity to revise certain well-beloved notions. In doing so, I will put emphasis on the comparisons of Arctic and Antarctic conditions, based on the rationale that the comparative analysis of the similarities and dissimilarities between the two polar systems is particularly suited to deduce general ecological implications. However, to set the framework for the further discussions, I will first give brief overviews on the most salient features of contemporary environmental conditions and geological origin of the Arctic seas, as well as on the state of knowledge on Arctic benthos at the beginning of the 1990s.
Geographical, environmental and historical setting
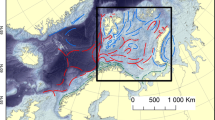
There is no precise geographical definition of the term ‘Arctic seas’, as one can find different southern demarcations in the literature. In the following, a widely accepted scheme proposed on the basis of comprehensive studies of the distribution patterns of benthic species is adopted (Zenkevitch 1963): besides the deep-sea basins in the Arctic Ocean proper as well in the Greenland Sea, the “Arctic seas” encompass the waters over the continental margin of Greenland, the Eurasian shelves of the Kara, Laptev and East Siberian Seas, the Amerasian Chukchi Sea, the American Arctic shelves of the Beaufort Sea and the Canadian Archipelago (including Baffin Bay), as well as the regions north of the Polar Front of the Barents and Bering Seas (Fig. 1).

Map of the Arctic Ocean and its marginal seas
In general, the contemporary ecological setting in all Arctic seas is primarily characterized by very low, but relatively constant water temperatures, permanent or long-lasting seasonal ice cover, as well as very pronounced seasonal fluctuations in insolation and, hence, primary production (Hempel 1985). However, there are also some ecologically important regional contrasts between the various marginal seas and the Arctic Ocean proper due to differences in geographical position, topography, climate, and hydrography (Curtis 1975; Grebmeier and Barry 1991).
On a geological time scale, the Arctic seas as extreme cold-water biotopes are relatively young, a fact that is particularly evident if compared to Antarctica (Dunbar 1977). For the Southern Ocean, the onset of a significant cooling can be traced back to the Eocene/Oligocene boundary about 40 million years ago (Clarke and Crame 1989). At latest with the opening of the Drake Passage between South America and the Antarctic Peninsula and the formation of the Antarctic Convergence in the early Miocene about 23 million years ago, an isolated cold circum-Antarctic Southern Ocean had evolved (Thomson et al. 1991). In contrast, the polar regions of the northern hemisphere continued to have temperate climate conditions during the entire Miocene, i.e., for further 19 million years. The Arctic Ocean was largely ice-free and had open connections to both the Atlantic and Pacific Ocean until the onset of a drastic fall of sea temperatures in the Pliocene about 4 million years ago. The decrease in temperature even intensified since the beginning of the Pleistocene 1.8 million years ago. This epoch was characterized by periodic alternations between ‘cold’ glacial and ‘warm’ inter-glacial climate conditions, which resulted in the evolution of the contemporary geographical setting: a permanently ice-covered central Arctic Ocean fringed by shelf seas with seasonally varying sea-ice cover (Bleil and Thiede 1990).
The Arctic shelves in particular are inhabited by ‘young’—in evolutionary terms—benthic assemblages. Because of the pronounced global sea-level variations between glacial and inter-glacial intervals of about 100 m, the shelf regions had repeatedly fallen dry and/or had, at least partly, been covered by huge glaciers during the Quaternary glacial intervals. Therefore, they had to be recolonized repeatedly by marine fauna during the inter-glacial transgressions following the recurrent Pleistocene glaciations, the last of which ended only about 13,000 years ago (Zenkevitch 1963). It should be noted, however, that vast portions of the markedly deeper and narrower shelves of Antarctica, which are usually thought to be characterized by much more stable conditions than their Arctic counterparts, were also scraped largely free of benthic biota by extending ice shelves during glacial intervals (Clarke and Crame 1989). For the Arctic, there is even evidence that substantial erosion by advancing large Eurasian ice sheets also affected greater water depths (Spielhagen 2001), e.g., at the Yermak Plateau off NW Spitsbergen at 550 m below sea level (mbsl) about 660,000 years ago (Myhre et al. 1995), in the St. Anna Trough in the Kara Sea at 630 mbsl during the late Weichselian ice age about 20,000 years ago (Polyak et al. 1997) or at the Lomonossov Ridge at 1,000 mbsl during the Saale glaciation 180,000–130,000 years ago (Jakobsson et al. 2001). There is no doubt that these events have very likely led to massive destruction of the marine benthic fauna in extensive areas of the Arctic, particularly on the shelves. To avoid total eradication, relic populations, which served as seeding sources for the subsequent re-colonization process, must have survived somehow, either in deeper waters or in protected areas where ice scouring was not so devastating. One can easily imagine that the alternating loss and subsequent re-colonization of vast shelf habitats had pronounced impacts on the diversity of benthic communities as well as on the adaptability of benthic organisms.
Besides this long-term climate changes, the Arctic seas have also experienced environmental variations on shorter time scales. For example, over the last century Arctic surface air temperatures rose significantly between 1925 and 1945, decreased between 1950 and 1970 and have risen again since 1980 (Lozán et al. 2001). In response to the recent trend in atmospheric warming, the spatial extent of the summertime Arctic pack ice has been reduced by about 30% over the last three decades (Cavalieri et al. 1997), and also the average thickness of the pack ice seems to have been shrunk by a corresponding proportion (Rothrock et al. 1999). It has been recognized that such climate variations follow a cyclic pattern that is not restricted to the Arctic (‘Arctic Oscillation’ AO) but affects the entire northern hemisphere (‘North Atlantic Oscillation’ NAO) (Dickson et al. 2000). However, models suggest that climate changes will be particularly pronounced in the Arctic and that this region may warm ~3–4°C or more than twice the global average under realistic greenhouse warming scenarios (IPCC 2001). Such changes will not only have immediate effects on Arctic mammals and sea-ice biota (ACIA 2004) but will most likely also have severe impacts on both pelagic productivity and organic matter export and will, hence, also affect benthic communities.
State of knowledge and research perspectives in the early 1990s
The fact that the scientific research in Arctic waters has been intensified considerably since the late 1980s does not at all mean that the benthos of the northern seas had been unknown to science before. As Dayton (1990) rightly pointed out, the history of the scientific exploration of Arctic regions is not as short and fragmentary as one might suppose because of their remoteness, inaccessibility and extreme climatic conditions. In the course of quite a number of field studies, most of which were conducted in the continental seas fringing the deep Arctic Ocean proper, a wealth of information on various aspects of the Arctic benthic fauna has been collected.
Surprisingly—and in contrast to a common notion—the vast Eurasian–Arctic waters off the Russian and Siberian coasts were at first better investigated than other Arctic regions. However, this fact remained largely unnoticed in the international scientific community because the research results were nearly always published in Russian publications that could neither be retrieved nor be understood by the vast majority of western scientists. Until the late 1980s, their knowledge of this enormous body of information relied almost exclusively on the English translation of Zenkevitch’s (1963) seminal textbook ‘Biology of the seas of the USSR’ that had actually already been published in Russian in 1955. In this comprehensive volume, the extensive research conducted by Russian scientists in the northern Soviet seas in the 1930s, 1940s and 1950s was synoptically presented, and it is still outstanding in its comprehensive description of the biogeography, composition, abundance and biomass of the Eurasian–Arctic benthos. The results of the more recent work of Russian researchers were made available to the western scientific community by the book edited by Herman (1989).
In the western Arctic, systematic benthological work started with an inventory of the benthic fauna off Point Barrow (Chukchi Sea) in the late 1940s and early 1950s (MacGinitie 1955). In the 1980s, the first multidisciplinary projects, including benthological studies, were launched in the Southeastern Bering Sea (‘Processes and resources of the Bering Sea’, PROBES; McRoy et al. 1986) and the northeastern Bering Sea and the Chukchi Sea (‘Inner Shelf Transfer and Recycling’, (ISHTAR; McRoy 1993). These projects led to some very important conclusions, such as the general importance of frontal zones for both pelagic and benthic systems, the significance of the pelago–benthic coupling and the existence of high-biomass assemblages in the northern Bering and southern Chukchi Seas.
From a western point of view, the information on Arctic benthos has repeatedly been summarized in several monographs and review articles, each with a different focus (e.g., Curtis 1975; George 1977; Knox and Lowry 1977; Hedgpeth 1977; Hempel 1985; Dayton 1990; Carey 1991; Grebmeier and Barry 1991). Summarizing the state of knowledge compiled until the early 1990s, as it has been presented in these publications, the following general statements about the Arctic benthos and the knowledge about it can be made:
-
Arctic benthic assemblages are rather poor in diversity and feature more cold-eurythermal boreal immigrants than truly Arctic species (endemics). This was considered as a salient characteristic of the Arctic seas in general—as well as a major difference to the Southern Ocean—and was attributed to the short geologic age of the Arctic; because of its rather unstable and oscillating environment, as well as its little developed biogeographical isolation, it has relatively intense faunal exchange with boreal regions.
-
Benthic standing stocks range broadly from very small under food-limited conditions (e.g., in the deep Arctic Ocean) to quite high in case of tight pelago–benthic coupling (e.g., in the seasonally ice-covered parts of the Bering, Chukchi and Barents Seas).
-
Physical and biological disturbance levels were hypothesized to be exceptionally high in Arctic waters and to have an important impact on the structure and functioning of benthic systems. This was assumed to be in stark contrast to Antarctica, which was thought to be characterized by markedly lower disturbance levels, resulting in the surprisingly high standing stock of the nearshore deep Antarctic shelf benthos (because stability, together with cold temperatures, enable benthic organisms to grow larger than in the Arctic).
-
Pelago–benthic coupling was regarded as a crucial process for the benthos in both polar regions, as it determines the food supply to the benthos from the overlying water column and, hence, directly influences benthic community abundance and biomass. Since the strength of the pelago–benthic coupling is, in turn, largely regulated by water-column factors, such as the hydrographic regime, ice coverage, primary production and pelagic food web structure, oceanographic processes have a great impact on polar benthic systems. In general, the relative strength of the pelago–benthic coupling was assumed to increase with latitude (irrespective of clear regional differences between the Arctic seas), thus leading to the ‘paradox’ of low water-column production and high benthic populations in many Arctic as well as Antarctic areas.
-
As Arctic shelf seas are characterized by high carbon inputs from terrestrial sources, benthic communities were hypothesized to make use of this additional food supply to a considerable degree. This was regarded as a further profound difference to the conditions in Antarctic waters where terrestrial inputs are virtually absent.
-
The information base, from which these conclusions were drawn, was unevenly distributed among the various Arctic Seas: while the Greenland, Barents, Bering and Chukchi Seas were comparatively well studied, the knowledge of less accessible regions, such as the central Arctic Ocean and the Eurasian–Arctic shelf seas, was very scarce.
-
While the knowledge on taxonomy and zoogeography was considered to be comparatively good, more work on quantitative characteristics (e.g., benthic secondary production rates and energy flow patterns), life-cycle adaptations (e.g., overwintering strategies) and the effects of pollution and anthropogenic perturbations was advised.
-
Therefore, it was commonly acknowledged that there was a need for a broad intensification of research on Arctic benthos in general and on the fauna in the regions, in particular, which were hardly explored so far. As major research themes, the significance of pelago–benthic coupling, the role of disturbance, the partitioning of carbon flow within the benthic communities (i.e., among microorganisms and the different fauna size classes), and the ecological consequences of global climate change (e.g., reduction of ice cover, increase of primary production, freshwater input and suspended load) for the benthos were proposed. However, this ecosystem-oriented work should be accompanied by further autoecological research on basic natural history and systematic studies.
-
To adequately address these ambitious issues, long-term multidisciplinary research programmes, possibly throughout the Arctic Basin and its surrounding seas, were strongly advocated. They should combine large-scale standardized surveys, including time-series studies, as well as the use of novel techniques in field work (e.g., direct observational work and in situ experiments, possibly throughout the year) and laboratory studies (e.g., physiological and behavioural investigation of living organisms maintained over lengthy periods of time).
-
Because the stark contrast between Arctic and Antarctic benthic systems is intertwined with obvious environmental similarities, comparative research projects in both polar regions—on subjects such as the interplay between nutrients, primary production and growth/reproduction rates, the significance of benthic–pelagic coupling on larval settlement and nutrient transfer, the patterns of succession and persistence stability, as well as ecological consequences of predator–prey relationships and perturbation caused by humans—were regarded to be particularly fertile.
Novel findings and their implications
History has shown that the call for integrated large-scale projects in the late 1980s has indeed been heeded; the impressive record of such research efforts has been indicated in the “Introduction”. Zoobenthic community structures, stock sizes and distribution patterns were investigated in almost all these projects. Overall, the Arctic shelf macro- and megafauna have received more attention than meiofaunal and microbial communities. Some studies aimed at a better understanding to which degree benthic communities of the Northern North Atlantic, the central Arctic Ocean and its adjacent Arctic shelf seas depend on the import of allochthonous carbon from the surface or from adjacent regions (see “Pelago–benthic coupling”). Other studies also attempted to analyze whether variations in benthic distribution patterns might be related to the environmental change (see “Consequences of climate change”). Regardless of their major research goals, all investigations provided a wealth of novel information which had—not surprisingly—different implications for the common notions about the Arctic benthos. Some of them were largely supported by the new data while others were not, especially those regarding the pronounced difference between Arctic and Antarctic systems. In general, the findings of comparative surveys suggest that Hedgpeth’s (1977) often-cited statement that “there is very little in common between the highly diversified Antarctic benthos fauna with its very dense epifaunal assemblages and the more monotonous predominantly infaunal assemblages of the Arctic basin” is a gross oversimplification of a rather complex and scale-dependent pattern of similarities and dissimilarities between the two polar biota.
Biogeography, species composition and endemism
The notion that most organisms inhabiting Arctic shelf and slope habitats are actually widespread boreal-Arctic species (Anisimova 1989; Smirnov 1994a) and that Arctic benthic communities thus bear quite a high faunistic similarity to North Atlantic ones (Zenkevitch 1963) has repeatedly been corroborated (Kröncke 1994, 1998; Clough et al. 1997b; Deubel 2000). Such recurrent findings have long been emphasized as strong indicators for the young age and the little developed isolation of the Arctic fauna (Zenkevitch 1963; Dunbar 1977; George 1977; Knox and Lowry 1977; Dunton 1992; Smirnov 1994a). This concept received much attention in the past because it contributes significantly to the astonishingly well-defined disparities between the two polar systems: in contrast to the conditions in the Arctic, most species and even higher taxa of the Antarctic benthos are endemic for the Southern Ocean, i.e., they are confined to the waters south of the Antarctic convergence (White 1984). The striking dissimilarity in the degree of endemism is commonly attributed to the great differences in geological age and biogeographic isolation (Hempel 1985).
Conclusion The findings gained during the past 15 years in both shelf and deep-sea regions did not provide any surprising evidence that would call for a shift of the paradigm that most species inhabiting the Arctic seas are wide-spread boreal-Arctic species whereas Antarctic species are more endemic.
Diversity, stability and disturbance
With regard to diversity, considered as a fundamental and very often studied benthic community attribute, matters are a bit more complicated. First, diversity itself is not a simple concept because it encompasses two independent aspects: (1) species richness and (2) species evenness (in the distribution of individuals among species), both of which can be measured—either separately or simultaneously—by a wide variety of different parameters (Magurran 1988). Second, it is very important to distinguish between ‘inventory diversity’, i.e., the diversity within an area of a given extent, and ‘differentiation diversity’, i.e., the species turnover between different areas (Whittaker 1972). Third, both these diversity modes can be measured at different spatial scales (Whittaker 1972). In case of inventory diversity, four different levels are commonly used (Gray 2001): ‘point’ (diversity related to a single sample), ‘alpha’ (total diversity of several samples within a habitat, i.e., at a local scale), ‘gamma’ (total diversity of a larger area, e.g., an island or a landscape, i.e., at a regional scale), and ‘epsilon’ (total diversity of a group of gamma-diversity areas, e.g., a biogeographical province). In case of differentiation diversity, for example, the term ‘beta’ refers to the rate of species turnover between habitats (local to regional scale). As these are relative levels without well-defined absolute boundaries, there is much confusion about the terminology in the scientific literature (Gray 2001). And fourth, regardless of which diversity aspect or mode is measured at whichever level with whichever parameter, the actual values gained are very much dependent of a host of methodological factors (e.g., sampling gear, mesh size, taxonomic expertise, etc.). Facing all these constraints, it is not surprising that any comparison of diversity between different study areas, especially if they are based on literature reviews of various surveys (which almost certainly differ in scales and methods), are very problematic (Hurlbert 1971), as the probability of severe methodological bias is quite high. Therefore, several authors have cautioned (Gutt 1991; Arntz et al. 1994; Clarke 1994; Arntz 1995) that the basic conclusions of such reviews might well be undue overgeneralizations.
A key question for polar benthic studies is whether, in terms of diversity, the Arctic is poor relative to lower latitudes and the Southern Ocean. With regard to large-scale (biogeographical) species richness, which is least susceptible to bias by systematic errors, the common notion was quite clear 15 years ago (Knox and Lowry 1977): Arctic seas harbor markedly less species than other regions of comparable size. However, recent studies have provided some contrasting evidence that advises caution when answering the question posed above. For some taxa, the hypothesis of a generally impoverished Arctic benthos has been corroborated. It is evident, for instance, that only few brittle star species, ranging from 15 (Smirnov 1994a) to 22 (Anisimova 1989), succeeded to permanently dwell in Arctic waters (Piepenburg 2000). These species numbers are very low, even if compared with other major taxa in the Arctic. Ophiuroids, although being the largest of the echinoderm classes with a total of about 2,000 extant species (Smith et al. 1995), do not rank in general among the very diverse taxa; and only about 1% of them occur in the Arctic. In other regions of comparable size, ophiuroid species numbers are distinctly higher, e.g., in the northern Pacific (58 species) (D’yakonov 1967), the abyssal northern Atlantic (about 120 species) (Paterson 1985) or the Southern Ocean (about 150 species) (Smirnov 1994b).
If the macrozoobenthos as a whole is considered, the picture is not that clear-cut. According to Sirenko and Piepenburg (1994) a total of more than 4,000 species have been reported from the entire Arctic, of which >1,000 were crustaceans, >400 molluscs, and >300 annelids. Seven years later, Sirenko (2001) listed a total of about known 4,800 macrobenthic species for the entire Arctic, a total of 2,895 known species for the Eurasian–Arctic shelf seas, and a total of about 400 known species for the deep Eurasian basins of the Arctic Ocean—a region that was almost unknown 15 years ago but has since been investigated in a number of studies (see below). With regard to the entire Arctic this is 20% more than in 1994. This increase can mainly be attributed to recent research efforts, such as the 10-year Russian–German Laptev Sea study (Kassens et al. 1999). It indicates that the species inventory is far from being complete for the Arctic shelves as well as, in particular, the even less accessible deep Arctic Ocean. For comparison, Arntz et al. (1997) and Clarke and Johnston (2003) estimated the number of all known macrozoobenthic species from Antarctica to range between 4,100 and 5,000. In conclusion, even if large-scale comparisons should always be interpreted with care because of the uneven distribution of sampling effort, the Arctic seems not to be as poor as previously thought. Gutt et al. (2004) concluded that the number of macrozoobenthic species for the Antarctic is only slightly higher than in the Arctic and that, at a global scale, both seem to have intermediate species richness—a notion that is clearly different from the old paradigm of a very poor Arctic benthos.
Does this conclusion also apply to other diversity aspects and other spatial scales? In a rigorous sense, valid diversity comparisons at smaller (regional and local) scales require case studies in comparable habitats using strictly standardized approaches to minimize methodological bias. In case of assessments along a wide latitudinal or even inter-polar gradient, this is quite difficult to achieve, and such investigations are thus rather rare. On the basis of strictly comparative benthic case studies, Kendall and Aschan (1993) and Kendall (1996) demonstrated that alpha diversities of soft-sediment shelf benthos assemblages around Svalbard (80°N) were not smaller than those of similar assemblages inhabiting similar substrata at similar depths in Java (7°S) and the North Sea (55°N). This finding suggests that there is no distinct latitudinal gradient in soft-bottom benthos diversity and supports the argument that high-latitude assemblages are not impoverished with regard to local diversity. Another comparative case study showed that various diversity parameters (species richness, Shannon diversity, evenness) of the ophiuroid fauna of the eastern Weddell Sea (Antarctica) were indeed significantly higher at both local (alpha) and regional (gamma) scales than off Northeast Greenland (Arctic) but that the values of the assemblages inhabiting the southern Weddell Sea shelf and shelf trenches were not (Piepenburg et al. 1997). Moreover, as many species are phylogenetically more closely related in the Weddell Sea than off Greenland, the Antarctic assemblages also did not differ significantly from those distinguished in the Greenland Sea in terms of ‘taxonomic diversity’ and ‘taxonomic distinctness’ (Warwick and Clarke 1995). These results suggest that the paradigm of a consistent Arctic–Antarctic diversity difference is a gross overgeneralization, at least with regard to certain taxa, such as brittle stars, and to regional and local scales (Piepenburg 2000).
Diversity—as well as other key community traits such as biomass and production (see below)—is closely related to ‘stability’ and ‘disturbance’. These terms are both not very clear ecological concepts, and one can find a multitude of definitions in the scientific literature. According to Pickett and White (1985), disturbances are “relatively discrete events in time, triggered by either endogenous or exogenous causes of either abiotic or biotic origin and differing in frequency and intensity, which disrupt ecosystem, community, or population structure and change resources, substrate availability, or the physical environment”. The great significance of disturbance as a key determinant of Arctic benthic shelf assemblages has already been emphasized by Dayton (1990) and Grebmeier and Barry (1991). They argued that benthic community patterns (diversity, biomass and productivity) vary not only as a result of food availability (see below) but also in response to disturbance levels. These levels are generally high in marginal Arctic seas—due to physical disturbances (ice gouging, freshwater and sediment input leading to variable salinities and high turbidity levels, variable ice cover), biological disturbances (feeding activities of invertebrate predators, such as neptunid gastropods, ophiuroids, asteroids and brachyuran crustaceans, as well as marine mammals, such as grey whales and walruses), and also increasing anthropogenic disturbances (Jewett et al. 1999)—and, hence, are assumed to limit overall benthic diversity and production. In contrast, Antarctic shelves were thought to be characterized by markedly low disturbance levels that enable benthic communities to reach high species diversities and benthic populations to grow to high levels of biomass in spite of rather low water-column production.
Recent research work has provided evidence that the notion of an ‘unstable and, hence, poor Arctic benthos’ versus a ‘stable and, hence, diverse Antarctic benthos’ is too simple. It has been demonstrated that grounding icebergs or deep pressure keels of drifting sea ice exert catastrophic disturbances on benthic habitats not only in the Arctic (Conlan et al. 1998), where they can—in addition to their abrasive primary effect—lead to local hypoxia at gouged seabed patches (Kvitek et al. 1998), but also in the Antarctic (Peck et al. 1999). There, they apparently happen much more often than as previously thought. Iceberg plough tracks were detected down to the depths of 450 m. At certain exposed locations, they cover 50% of the seabed, indicating that up to 7% of the total area of the Antarctic shelves (<350 m) might be affected by grounding impacts within the past 20 years (Gutt 2001). Like other discrete events, such as forest fires, tree falls or hurricanes, iceberg scouring is now recognized to be fundamental to the functioning of the entire Antarctic shelf ecosystem, as their disastrous initial impact is followed by positive effects on biodiversity (Gutt and Piepenburg 2003). Novel findings suggest that two processes, operating on different spatial and temporal scales, regulate the diversity of Antarctic shelf benthos. In accordance with the ‘stability-time hypothesis’ (Sanders 1968), the high local diversity of undisturbed Antarctic shelf benthos is primarily the result of the evolution of sponge-dominated assemblages over long and stable time spans. However, in accordance with the ‘intermediate disturbance hypothesis’ (Huston 1979), the diversity on regional scales—possibly even that on a larger scale encompassing the entire Antarctic shelf—is strongly influenced by catastrophic iceberg disturbances.
Grounding icebergs—as well as grounding pressure ice keels of drifting pack ice in the Arctic—devastate large seabed patches and virtually eradicate the benthos at these places. However, after being released, they leave behind free seafloor space that can be recolonized. Over a longer time span and at a regional scale, iceberg groundings positively affect the spatial and temporal habitat diversity and prevent the competitive displacement of species, which is characteristic for undisturbed systems near the ecological equilibrium and results in a reduction of diversity. They create a mosaic of habitats in different succession stages and thus enhance beta (between-habitat) and, hence, gamma (regional) diversity. Moreover, indirect disturbance effects of iceberg or sea-ice scouring, such as changes of small-scale bottom-water current regime or modification of small-scale seabed topography, may have further ecological consequences, which may be even more far-reaching and, hence, possibly even more significant than the initial mechanical habitat destruction through the abrasion of large seabed patches.
Conclusions As it has been demonstrated that seabed assemblages vary broadly in faunistic diversity among Arctic regions, the idea that the Arctic benthos is generally very poor—and much less diverse than the rich Antarctic bottom fauna—is an undue oversimplification. There is some evidence that, in terms of large-scale diversity, both Arctic and Antarctic waters apparently do not differ much and seem to be characterized by intermediate species richness. Recent results, especially with regard to the ecological role of ice gouging, suggest that the notion of ‘high disturbance levels in the Arctic versus low disturbance levels in the Antarctic,’ which has been a major argument for explaining the striking differences between the two polar systems, should be modified. However, to draw a definite conclusion there is still a great need for quantitative studies, preferably combining field work, laboratory analyses and numerical modelling, to determine the responses of benthic organisms and/or communities to natural and anthropogenic disturbances.
Communities, standing stocks and distribution patterns
A fundamental conclusion drawn from the findings of all benthic surveys conducted in the recent past is that there is not just one typical Arctic benthos but a wide variety of communities found in distinct depth zones (shelf, slope, and basin) and regions, which differ profoundly in almost all aspects of benthic ecology. This variation is caused by differences in, e.g., water depth, geographical setting, biogeographical history, water current and advection regime, river runoff, ice cover, seafloor composition, and food availability. The ecological effects of these factors are often interrelated (Piepenburg et al. 2001). River runoff, for instance, strongly affects sea-ice dynamics and oceanic circulation pattern, hence pelagic and sympagic productivity regime, and therefore ultimately also benthic food supply. In addition to this important indirect effect, fluvial discharge, as well as coastal erosion, can result into a significant import of suspended inorganic and organic matter, especially in the Siberian marginal seas. The latter is subject to various geochemical and biological transformations, and might, for instance, serve as an allochthonous energy source within the marine food webs. Clearly, the knowledge about the quantity, spatio-temporal distribution, and fate of the terrigenic organic matter is essential for a profound understanding of its relevance for the pelagic and benthic systems in the Arctic seas (Fahl et al. 2001).
The macrofaunal assemblages in different regions and at different depths are characterized by typical indicator taxa. Shelf regions with fine sand and mud sediments, for instance, are often dominated by bivalves and polychaetes (Feder et al. 1994a; Grebmeier and Cooper 1995; Deubel et al. 2003), while gammaridean amphipods are the most prominent faunal elements in coarse-grained sediments (Grebmeier et al. 1995). A general circumpolar pattern in the composition of epibenthic communities is the pronounced numerical importance of brittle stars in a great number of Arctic shelf and slope habitats (Starmans et al. 1999; Piepenburg 2000; Sejr et al. 2000; Ambrose et al. 2001). Other conspicuous epifaunal megabenthic elements, which may even exceed the ophiuroid stocks in terms of biomass, are sea urchins in the Barents Sea (Bluhm et al. 1998), sea cucumbers and bivalves in the Laptev Sea (Piepenburg and Schmid 1997), as well as sea stars and crustaceans in the Bering and Chukchi Seas (Jewett and Feder 1981; Feder et al. 2005).
A recurrent finding of several recent benthic surveys was that many Arctic shelf assemblages were strongly dominated by the brittle star Ophiocten sericeum. Photographic censuses showed that on the Greenland, Barents and Laptev Sea shelves, this species reaches several hundreds ind.m−2 in abundance (Piepenburg and Schmid 1996a, b, 1997), forming very dense brittle star beds as they have also been reported from several locations in different climatic zones (Aronson 1989). In these beds, O. sericeum account for up to >99% of the ophiuroid fauna and for up to 98% of total epibenthic abundance. At some sites, however, other species, e.g., Ophiura robusta off Northeast Greenland (Piepenburg and Schmid 1996b) or Ophiura sarsi in the Chukchi Sea (Ambrose et al. 2001), can attain similarly high densities (>100 ind.m−2).
The finding of very high O. sericeum abundances was surprising for the Laptev Sea. The benthos of this high-Arctic shelf region had been reported to be distinctly poorer, in terms of diversity but also in abundance and biomass, than in other Eurasian–Arctic seas (Zenkevitch 1963, Golikov 1990). The scarcity was attributed to the most severe climate in the Laptev Sea, as well as to very low salinities due to a pronounced fluvial dilution during summer. Accordingly, the shallow shoals of the Laptev Sea (<30 m) have been reported to be poor in terms of brittle star abundance and biomass (Piepenburg and Schmid 1997). However, on the flanks of the shelf valleys (>30 m) that are not subject to reduced and fluctuating salinities and possible iceberg gouging impacts, very high densities of O. sericeum have been recorded. These figures are similar to that of the maximum values found for O. sericeum on shelf banks in the Barents Sea (Piepenburg and Schmid 1996a) or at the Belgica Bank off Northeast Greenland (Piepenburg and Schmid 1996b). Maximum ophiuroid biomass values were not lower in the Laptev Sea either: nearly 2 g C m−2 in the Laptev Sea, which were primarily made up by Ophiura sarsi (Piepenburg and Schmid 1997), versus about 2.5 g C m−2 in the Barents Sea (Piepenburg and Schmid 1996a) and about 1 g C m−2 off Northeast Greenland (Piepenburg and Schmid 1996b), which were mainly made up by O. sericeum. Except for the very shallow Laptev Sea shoals (<23 m), there was no significant difference between the Greenland, Barents and Laptev Sea shelves in terms of either ophiuroid abundance or biomass.
With regard to brittle stars, the standing stock figures reported from high-Arctic offshore shelf sites are among the highest hitherto observed in northern seas or sub-Arctic regions (Piepenburg 2000). They reached the same order of magnitude as the abundance and biomass of dense brittle star populations reported from bathyal non-polar regions of the Atlantic and Pacific. In comparison with values reported from the Southern Ocean (Brey and Clarke 1993; Dahm 1996), abundances of Arctic brittle stars appear to be somewhat higher, whereas there was no pronounced difference in terms of biomass. In conclusion, the ophiuroid standing stocks on Arctic shelves are surprisingly high. On average, they match those observed for highly abundant endobenthic Amphiura and/or epibenthic Ophiura species on boreal shelves (Muus 1981; Salzwedel et al. 1985; Sköld et al. 1994).
A pronounced depth zonation in the composition and distribution of benthic assemblages, accompanied by an exponential decline of benthic standing stock along a shelf-slope-basin gradient, is a common phenomenon in Arctic seas (Curtis 1975). In this regard the Arctic does not differ from other regions, as similar bathymetric patterns have recurrently been detected in numerous studies for a wide range of latitudes, water depths and benthic taxa (Rowe et al. 1974; Haedrich et al. 1980; Stewart 1983; Lampitt et al. 1986; Brey and Clarke 1993; Dahm 1996). The actual cause of this ubiquitous pattern is difficult to assess. Most scientists agree in that any zonation observed must very likely to be viewed as the result of not just one single factor but of several direct and indirect processes operating on various spatial and temporal scales (Carney et al. 1983). It is commonly acknowledged that hydrostatic pressure, the factor directly related to water depth, does indeed cause specific physiological adaptations of the organisms (Somero et al. 1983) but is of only marginal significance for explaining the large-scale (100–1,000 km) depth zonation in composition and standing stock of benthos assemblages (Somero 1990). Results from many field studies rather suggested that this is primarily caused by gradients in food availability and sea-bed properties. Both determinants are known to be strongly related to water depth, hydrodynamics and various processes of particle transport, such as turbidity plumes, Taylor columns and internal waves. There is a general inverse relationship between sedimentation rates and water depths (Suess 1980; Martin et al. 1987), which explains the fundamental influence of water depth on the quality and quantity of organic carbon reaching the sea floor and, hence, food supply for the benthos (Graf 1992). In energy-limited systems, such as the deep sea or polar seas, food supply has repeatedly been proposed as the prime agent controlling meio-, macro- and megabenthic biomass, being more important than physiological adaptations, biological interactions or competition for space (Hessler and Jumars 1974; Rowe et al. 1974; Aldred et al. 1979; Lampitt et al. 1986; Grebmeier and Barry 1991). The novel results on Arctic benthos were consistent with this chain of arguments, which points to the special significance of the pelago–benthic coupling for the benthos in Arctic waters.
Conclusions Recent field data have clearly demonstrated that the various Arctic seas differ markedly in the composition, stock size and distribution of benthic assemblages, which largely mirrors the differences in geographical and environmental setting (particularly in water depth), hence in the impact of fluvial run-off, pelagic production regime and strength of pelago–benthic coupling, and hence in the food supply to the benthos. This pronounced heterogeneity severely impedes large-scale generalizations of local and regional findings and calls for a pan-Arctic perspective in the quest to advance the fundamental understanding of key features of polar marine ecology.
Pelago–benthic coupling and the significance of food supply
Besides the large-scale depth zonation, the most prominent feature in the distribution of Arctic benthos assemblages is the spatial concordance with meso-scale (10–100 km) patterns in current regime and sea-ice cover, such as marginal ice zones, polynyas, and gyres (Piepenburg 2000). This finding further emphasizes the importance of pelago–benthic coupling for the benthos, as these patterns are known to be related to variations in the primary production regime, the sedimentation of organic matter out of the water column, and, hence, the food supply to the seabed.
It is well known that benthic distribution and community features, such as composition, diversity, and standing stock, are influenced by a complex of abiotic and biotic factors (Dayton 1984). In the scientific literature, a wide variety of different, and partly interacting parameters, have been discussed in this context, e.g., water depth, habitat heterogeneity, sea floor properties, bottom-water hydrography and current regime, food availability, as well as inter- and intraspecific competition and disturbance caused by predation or burrowing activities (Gray 1981). These factors are not only connected within a complex network of interrelationships (Dayton 1984), which is often difficult to unravel, but their relative importance also strongly depend on the spatial scale considered (Dayton and Tegner 1984; Gage and Tyler 1991). In the recent past, a growing body of evidence has been compiled that seabed attributes, which have long been suggested to be of foremost importance (Snelgrove and Butman 1994), are usually most significant at small (i.e., local) scales, while the quantity, quality, and temporal pattern of food supply seem to primarily regulate the benthic distribution and stock size at larger (i.e., regional) scales (Dayton and Oliver 1977; Graf 1992).
It is an almost trivial statement that the food supply of the vast majority of benthic faunas depends entirely on the import of organic matter, ultimately originating from the autotrophic production in the upper euphotic layer of the water column (Tyler 1995). Of course, there are prominent exceptions from this rule, e.g., littoral habitats at comparatively small depths, to which sufficient sunlight can penetrate to sustain benthic primary producers, as well as the chemosynthetic communities at hot vents (Grassle 1986; Van Dover 2000) and cold seeps (Sibuet and Olu 1998). At a global scale, however, these biota are of only minor significance in spatial extent and energy budget. The downward flux of matter and energy from the water column to the seabed is the most prominent aspect of a fundamental relationship between the pelagic realm and the benthos, for which the term ‘pelago–benthic coupling’ has been coined (Hargrave 1973). It has generally been acknowledged since some time that the food supply to the benthos, hence the import of allochthonous organic matter, strongly affects a wide range of benthic patterns and processes, including biogeography, diversity, population densities, biomass, and the activity of the seafloor communities. Consequently, the notion that food availability is a principal benthic community determinant implies that the benthos is also strongly affected by abiotic and biotic water-column processes controlling the pelagic production as well as the sedimentation of organic matter to the seabed (Grebmeier and Barry 1991; Graf 1992). It should be noted, however, that on the other hand several studies have demonstrated that benthic organisms strongly influence the particle flux in the near-bottom water layer (Thomsen et al. 1995) and the deposition of particles in the sediment (Graf et al. 1995).
In general, primary production varies considerably among the various Arctic regions as a consequence of the effects of different promoting/impeding processes (Sakshaug 2003): upwelling of very nutrient-rich water along the shelf break (Bering Sea: >230 g C m−2 year−1), nutrient transport mediated by turbulence caused by Ekman pumping, tides, banks and islands, and shear forces (Nordic Seas, Atlantic Barents Sea: about 100 g C m−2 year−1), mixing of nutrient-poor shelf water (Siberian Shelf Seas: about 35 g C m−2 year−1), light limitation by multi-year ice (Central Arctic Ocean: 15–30 g C m−2 year−1). The proportion of new production ranges from about 2% of the total annual production in the central Arctic Ocean to 5–30% in the high-productive seas (Sakshaug 2003).
In polar seas, pelagic production and sedimentation of organic matter are strongly affected by sea ice (Honjo 1990; Smith and Sakshaug 1990). Furthermore, sea-ice algae can contribute considerably to the total productivity of polar seas (Legendre et al. 1992). The spatio-temporal pattern of sea-ice cover thus influences, via the pelago–benthic coupling, the trophic basis of seabed communities and has to be regarded as another important control agent of benthic systems, particularly for those on the shelves of Arctic and Antarctic seas (Grebmeier and Barry 1991; Arntz et al. 1994).
There is evidence that in high latitudes the strength of pelago–benthic coupling and, hence, food supply is of particular importance for the benthos. Clarke (1983) claimed that the amount of sedimenting food particles rather than the low temperatures per se control the metabolism, growth and survival of benthic organisms. Petersen and Curtis (1980) stated that, for similar depths and substratum types, benthic biomass seems to be greater in the Arctic than in boreal or tropical areas. On the basis of this observation, Petersen (1984) suggested a general increase in the efficiency of energy transfer between water column and seabed—and, thus, significance of pelago–benthic coupling—with increasing latitude. This pattern was assumed to be primarily caused by generally reduced zooplankton grazing and, hence, enhanced sedimentation rates of ungrazed organic matter to the seabed, resulting from the greater time lag in the response of zooplankton populations to the high seasonal oscillations in phytoplankton production at higher latitudes (Petersen and Curtis 1980). This contention has apparently been corroborated by the findings of a number of pelagic sedimentation studies, indicating that in higher latitudes a progressively larger proportion of the organic carbon fixed in the euphotic zone falls to the sea floor (Wassmann et al. 1991). Therefore, the benthos was postulated to generally have a greater role in the marine carbon production and turnover regime than at lower latitudes (Petersen and Curtis 1980) and, as a consequence, substantial benthic biomass in some areas support major feeding grounds of resident and migrating sea birds (Gould et al. 1982) and mammals (Highsmith and Coyle 1990, 1992). However, Grebmeier and Barry (1991) stated that the significance of the pelago–benthic coupling varies broadly among Arctic seas, reaching from high in biomass-rich areas to rather low in poor food-limited regions, and advised against making too broad generalizations. According to Grebmeier and Barry (1991), in most Arctic and Antarctic regions a large amount of the organic matter produced in the upper water column or the sea ice is consumed by zooplankton or recycled via the microbial loop before it reaches the seabed, resulting in food-limited regimes for the underlying benthos. However, some continental shelves, such as those in the Arctic Bering, Chukchi, and Barents Seas, where a tight coupling between pelagic/sympagic primary production and benthic secondary production causes high benthic standing stocks, are exceptions from this general pattern.
There is no doubt, however, that both polar regions feature prominent ‘hot spots’ of tight pelago–benthic coupling, i.e., certain meso-scale patterns in hydrography and sea-ice cover, which regionally enhance pelagic and/or sympagic primary production and the food supply to the benthos and, hence, have obviously a great impact on benthic spatial distribution patterns (see above). For instance, marginal ice zones (MIZ) are such well-known sites of locally and temporally increased production in Arctic and Antarctic ice-covered seas (Slagstad 1985; Sakshaug and Skjodal 1989; Niebauer 1991; Wassmann et al. 1991; Savidge et al. 1996). In particular ‘seasonally receding ice edges’, as they are found in the northern Barents Sea (Loeng 1989) and in the Laptev Sea (Timokhov 1994), apparently induce prolonged diatom blooms by favourable modifications of the local hydrography (Rey and Loeng 1985; Smith and Nelson 1985). Moreover, several sediment trap studies in both Arctic and Antarctic waters have demonstrated that a large amount of the organic matter produced in the MIZ tends to sink out of the euphotic layer in strongly pulsed sedimentation events (Honjo 1990; Hebbeln and Wefer 1991; Bauerfeind et al. 1994; Andreassen et al. 1996) and is thus exported as potential food for the benthos (Schewe and Soltwedel 2003). Not surprisingly, MIZs are known as regions of high benthic standing stocks (Grebmeier and Barry 1991; Piepenburg 2000).
Polynyas are permanent or recurrent ice-free areas in polar pack-ice zones, which have been identified to be of special importance for both physical and ecological processes (Stirling 1980; Smith et al. 1990; Gradinger 1995; Grebmeier and Cooper 1995). Pronounced meso-scale gradients characterize their hydrographic regimes (Schneider and Budéus 1994). Compared with surrounding ice-covered areas, pelagic production is often relatively high (Gradinger and Baumann 1991). Field studies in the Northeast Water (NEW) polynya off Northeast Greenland have demonstrated a tight pelago–benthic coupling. Results of isotope studies suggested that fresh ungrazed organic carbon reaches the sea bed below the polynya (Hobson et al. 1995), presumably due to low levels of zooplankton grazing (Hirche et al. 1994; Ashjian et al. 1995, 1997; Hirche and Kwasniewski 1997), and benthic abundance and biomass were reported to be considerably higher than in adjacent ice-covered regions (Ambrose and Renaud 1995; Brandt 1995; Piepenburg and Schmid 1996b). Similar conditions have been described for an area in the Bering Sea influenced by the St Lawrence Island polynya in winter/spring (Grebmeier 1993; Cooper et al. 2002). For the Laptev Sea, there is evidence that endobenthic biomass (Gukov 1995) as well as brittle star stocks (Piepenburg and Schmid 1997) are positively influenced by the spring flaw lead off the coastal fast ice belt. In general, high Chl a concentrations in the sediments indicated a tight coupling between sympagic and pelagic primary production and food supply to the benthos throughout the entire Laptev Sea (Schmid et al. 2005).
Another meso-scale oceanographic feature enhancing the strength of pelago–benthic coupling are anti-cyclonic gyres superimposed on major currents due to hydrodynamic effects induced by morphological shelf features like shallow banks or small islands (Bourke et al. 1987; Loeng 1989). These convergent eddies act as retention mechanisms that favour the food supply to the benthos by enhancing vertical particle flux and accumulating organic matter produced in adjacent productive areas (Feder et al. 2005). Moreover, they probably also have a positive effect on the survival and spatfall of meroplanktonic larvae of benthic species, such as the brittle star Ophiocten sericeum (Clough et al. 1997a). Therefore, such gyres are often found over shelf banks featuring exceptionally high benthic standing stocks (Piepenburg and Schmid 1996a, b).
Pelago–benthic coupling is certainly a key process of marine ecosystems and has, therefore, been an overarching theme of many research efforts in the past 15 years. In Arctic waters, the pathways and processes of the coupling between sea-ice, water-column and seabed biota have been studied quite intensively. The mechanisms and effects involved in the coupling have been investigated on different scales of time and space. For instance, the metabolic response of micro- and meiobenthic organisms to seasonal food pulses in terms of their activity and biomass has been shown to be quite rapid, i.e., within days (Graf 1989). In contrast, community patterns are pronouncedly more inert in their reaction to environmental forcing, especially those of macro- and megabenthic animals that grow larger and live longer than the micro- and meioorganisms (Gage and Tyler 1991). Hence, the distribution and structure of assemblages integrate the impact of control factors over longer periods of time. They reflect relatively enduring and/or predictable recurrent environmental states and can provide valuable clues to the long-term effects (i.e., at a time scale of months to years) of potential community determinants (Piepenburg et al. 2001).
All this convincing evidence of the great significance of pelago–benthic coupling and food supply for the benthos does, of course, not mean that the organic matter sustaining the bottom fauna originates exclusively or even primarily from the primary production in the overlying waters. It has been emphasized that, at a scale of an individual benthic organism, food supply by lateral advection is actually more important than the direct (inherently vertical) sedimentation (Graf 1992), as the velocities of even the slowest horizontal bottom currents are at least one order of magnitude higher than the sinking speeds of the fastest sedimenting particles. At larger spatial scales, allochthonous organic matter advected from adjacent more productive areas has repeatedly been shown to be an additional major food source of benthic communities in comparatively unproductive high-Arctic seas (Grebmeier 1993; Feder et al. 1994a, b, 2005). In the Barents Sea, for instance, the southward inflow of Arctic surface water is thought to be counterbalanced by a northward transport of warmer but more saline Arctic–Atlantic bottom water formed mainly at the Polar Front (Loeng 1989). These water masses might carry organic matter produced in the more productive southern Barents Sea to the north (Piepenburg and Schmid 1996a). In the northeastern Chukchi Sea, the advection of allochthonous POC helps to sustain a biomass-rich population of benthic ampeliscid amphipods that serves as the main food resource of migrating bottom-feeding grey whales in each summer (Feder et al. 2005). In general, the great significance of lateral advection for the dispersal of organic carbon in the benthos is indicated by the fact that the benthic carbon demand is only rarely matched by the carbon supply via sedimentation out of the overlying water column estimated from sediment-trap data (Christensen 2000).
Allochthonous food for the benthos can also originate from terrestrial production that primarily reaches the sea through fluvial transports. This source of organic carbon might be particularly important in the Arctic, as about 10% of the global river run-off enters the Arctic seas and leads to a strong coupling between terrestrial and marine ecosystems (Klages et al. 2003). Especially the Laptev Sea is heavily affected by fluvial inflow (Timokhov 1994), with the Lena river being estimated to discharge about 5.3×106 tons of organic carbon each year, most of it during the flood period in June/July (Cauwet and Sidorov 1996). Schmid et al. (2005) provide evidence that autochthonous primary production is apparently not sufficient to fuel both pelagic and benthic secondary production in the Laptev, implying that an input of allochthonous organic carbon is required to balance the overall carbon budget. However, most of the imported fluvial organic matter is fairly degraded (Stein 1996; Fahl et al. 2001) and, consequently, of probably rather poor nutritional quality. Its actual significance as food source for the bottom fauna is, hence, difficult to be estimated but is probably rather small (Klages et al. 2003).
Conclusions Pelago–benthic coupling has repeatedly been demonstrated as a key process of marine ecosystems, as it ultimately determines the level of food supply to the benthos. In polar waters, sea-ice cover strongly influences the strength of pelago–benthic coupling. Field evidence clearly indicates the great significance of meso-scale hydrography and ice cover (marginal ice zones, polynyas, and gyres) in forming ‘hot spots’ of tight pelago–benthic coupling and, hence, high benthic biomass.
Benthic carbon flow and its partitioning
Besides the assessment of the strength of pelago–benthic coupling, the quantification of benthic carbon fluxes and the description of its partitioning among different benthic community fractions have been major objectives of a number of recent research efforts. In most cases, the carbon flux through benthic communities has been estimated by assessing the sediment oxygen uptake, i.e., by incubating sediment cores and following the decrease of dissolved oxygen in the ambient overlying water with time (Smith and Hinga 1983). These measurements—regardless of whether they are performed in situ with benthic landers (Smith 1974) or using shipboard techniques (Hulth et al. 1994)—provide a bulk parameter, the ‘Sediment Community Oxygen Consumption’ (SCOC) or ‘Sediment Oxygen Demand’ (SOD), which integrates chemical oxygen uptake plus total aerobic respiration of all benthic organisms contained in the core. These respiration values, sometimes also called ‘benthic community metabolism’, are often converted to carbon remineralization rates using conversion factors based on Redfield ratios.
Recently, the current knowledge on the role of the Arctic benthos for the carbon cycle at the seafloor has been reviewed (Klages et al. 2003). Major conclusions from this review are that (1) the benthic carbon remineralization rates vary broadly depending on season, year, depth and sedimentary site characteristics and (2) there is a broad overlap between the rates determined at Arctic shelf sites (10–70 g C m−2 year−1) and those reported from coastal sediments in temperate and tropical regions (35–130 g C m−2 year−1), although pelagic productivities are up to an order of magnitude lower (40–100 g C m−2 year−1 vs. 200–750 g C m−2 year−1). The latter result is a surprising finding, as it suggests that (a) the low ambient temperatures in the Arctic do not limit benthic shelf community metabolism, and (b) shelf SCOC may not always reflect overlying productivity because the supply of oxygen from the bottom-near water to the sediment may limit this rate (Christensen 2000).
However, recent findings strongly suggest that SCOC rates can considerably underestimate the total benthic carbon demand. The cores used for sediment-water incubations, which are commonly used for the assessment of benthic carbon utilization, are usually rather small and cover only modest sample areas (<500 cm2). Therefore, they contain organisms ranging in body size from micro-, meio- to small macrobenthos but no megafauna. The population or assemblage respiration and carbon mineralization—as well as carbon demand (computed from mineralization values by applying suitable conversion factors to account for growth and assimilation efficiencies)—of these animals cannot be measured directly with an integrating method but has to be approximated by other approaches, such as combining abundance or biomass figures with individual respiration rates (Piepenburg 2000). Total benthic carbon remineralization can then be estimated by adding SCOC and megafaunal respiration values.
It has repeatedly been shown that megafaunal organisms, in particular echinoderms, dominate Arctic shelf benthos and can reach very high abundances (see above). For highly abundant brittle star populations (Ophiocten sericeum) in the Greenland, Barents and Laptev Seas, daily carbon mineralization rates and carbon demand reached maximum values of 5–11 mg C m−2 day−1 and 10 to >20 mg C m−2 day−1, respectively (Piepenburg 2000). A case study on the partitioning of carbon flow between different benthic community fractions in the Barents Sea showed that dense O. sericeum beds actually dominated the benthic carbon flux and mineralized an amount of carbon that was equivalent to the highest total sediment community carbon uptake recorded in the study area (Piepenburg et al. 1995). A comparison of ophiuroid carbon demand and summer (June–September) estimates of possible food supply (pelagic productivity and sedimentation of particulate organic carbon, POC) suggested that dense brittle star populations in the Barents Sea can consume up to 9% of the average net daily primary production and about 30% of the daily vertical POC flux to the seabed (Piepenburg 2000). In the Laptev Sea, these values were 7% and 20%, respectively (Piepenburg 2000). Ambrose et al. (2001) reported mineralization rates of up to 14 mg C m−2 day−1 for dense ophiuroid assemblages (Ophiura sarsi, O. maculata, Stegophiura nodosa, Ophiopholis aculeata) in the northern Chukchi Sea. This rate was about a third of the infaunal respiration that was concomitantly measured using the SCOC approach, implying that the echinoderm respiration accounted for about 25% of the total benthic respiration (Ambrose et al. 2001).
These findings from both the Eurasian and American Arctic have provided convincing evidence that epifaunal brittle star mineralization rates at high-Arctic shelves can be in the same order of magnitude as bulk infaunal mineralization rates measured in several Arctic sediments or assumed to be typical for upper slope habitats in general (Grebmeier and McRoy 1989). Furthermore, there is good reason to assume that other abundant epibenthic megafauna, e.g., sea stars in the Chukchi Sea (Feder et al. 2005), may also have considerable—but to date unknown—importance in the overall benthic mineralization. There is thus sufficient evidence to contend that respiration budgets, which are based exclusively on SCOC measurements, may severely underestimate the total benthic oxygen flux and carbon demand (Piepenburg 2000).
Conclusions Studies on the partitioning of energy flow through the benthic communities suggest that abundant epibenthic megafauna represent an important pathway of the benthic carbon and energy flow, the significance of which cannot be discounted in overall benthic energy budgets. Hence, it has to be adequately considered in the on-going attempts to understand how carbon is processed on Arctic shelves, which are commonly dominated by abundant echinoderm populations, and to build accurate models of carbon cycling in the Arctic.
The deep Arctic Ocean and the role of shelf–basin interactions
Our knowledge on Arctic benthos before the 1990s varied considerably among the various marginal shelf seas (and it still does), but it was particularly scarce for the deep-sea central Arctic Ocean, largely relying on occasional samples taken from Arctic ice drift stations (Paul and Menzies 1974). However, the past 15 years have witnessed particular progress in the exploration of this remote, almost inaccessible region, and a number of quantitative studies have provided novel information on the benthos of the Amerasian and Eurasian basins (Kröncke 1994, 1998; Clough et al. 1997b; Soltwedel and Schewe 1998; Schewe and Soltwedel 1999; Deubel 2000; Kröncke et al. 2000; Vanreusel et al. 2000; Bluhm et al. 2005).
The Arctic Ocean proper is unique in its physical and biological properties. It is characterized by the most extreme limitations in solar radiation and nutrient availability, permanent ice cover and temperatures always close to the freezing point—a combination of factors that intuition tells us to have a very negative effect on overall biological productivity. Therefore, the Arctic Ocean has long been considered to be one of the least favorable habitats for life on Earth and, hence, the poorest—in terms of both diversity and productivity—part of the world’s ocean. This notion is still not totally false but novel findings have called for a rectification of the estimates of annual (particulate) primary production by one order of magnitude (from 2 g C m−2 year−1 to 15–30 g C m−2 year−1; MacDonald and Carmack 1991; Macdonald et al. 1993; Wheeler et al. 1996; Gosselin et al. 1997; Sakshaug 2003).
The benthic inventories of the past 15 years corroborated previous results that deposit feeders dominate the benthic communities. Polychaetes, crustaceans, and bivalves were the most important taxa in the deep-sea samples, followed by sponges, cnidarians, tunicates, as well as echinoderms (Kröncke 1994, 1998; Clough et al. 1997b; Deubel 2000; Bluhm et al. 2005). The latter are thus important elements of the macrobenthic deep-sea fauna as well as in the marginal Arctic shelf seas. However, while ophiuroids often dominate echinoderm shelf communities (Piepenburg 2000; Ambrose et al. 2001), sea urchins and sea cucumbers usually do so in the deep sea, e.g., the irregular echinoid Pourtalesia jeffreysii and the holothurian Elpidia glacialis, both of which are, however, not confined to the Arctic but also occur at abyssal depths at lower latitudes (Heding 1942; Vinogradova 1979).
Furthermore, the recent investigations largely confirmed the expectation that infaunal species richness, as well as abundance and biomass, decrease along a shelf–basin gradient with water depth and latitude (Kröncke 1994, 1998; Clough et al. 1997b; Deubel 2000; Bluhm et al. 2005). They are generally at rather low levels (5–500 species m−2, 5 to 6,625 ind.m−2, and 5–130 mg C m−2), lying at the lower margin of values reported from the deep basins of the North Atlantic (Levin and Gooday 2003). These findings corroborated the notion that the major factor affecting the Arctic deep-sea benthos is the energy limitation caused by very limited organic matter supply to the abyssal seafloor.
The benthic carbon mineralization rates, estimated from SCOC measurements, range from 1 g C m−2 year−1 to 10 g C m−2 year−1, i.e., they are one order of magnitude lower than Arctic shelf values (see above) but by all means comparable to other oligotrophic oceanic regions (Klages et al. 2003). Recent foraminiferal investigations have revealed that benthic communities in the deep basins of the Arctic Ocean are driven by the sedimentation of fresh organic material (Kröncke et al. 2000). These findings imply that the autochthonous production in the Arctic Ocean, albeit being rather low, may be sufficient to nourish the underlying benthic deep-sea communities (Klages et al. 2003). This conclusion is contradictory to the notion that the Arctic deep-sea benthos relies trophically on the import of organic carbon from productive Arctic shelf regions (Grebmeier 2003). The finding that bacterial biomass and activity does not decrease along a bathymetrical and latitudinal gradient to the North Pole rather suggests that it is largely decoupled from the production on the adjacent marginal seas (Klages et al. 2003). Although there is also some evidence of lateral transport from terrestrial, coastal, and shelf sources to the central Arctic Ocean, the supply of utilizable carbon, feeding the abyssal benthic communities, apparently depends very much on the vertical flux of organic matter to the seabed, either from overlying pelagic/sympagic production or in the form of large food falls (Klages et al. 2001; Soltwedel et al. 2003). Additional food imported from the shelves does not seem to be necessary to meet the benthic carbon demand (Klages et al. 2003).
In general, continental shelves are regions that are characterized by strong lateral exchanges of heat and matter between both each other and adjacent deep-sea regions. These transport processes include the export of organic carbon, as the shelves are usually much more productive than oceanic areas and, hence, excess production is available. The significance of organic matter transport from the shelves across ocean margins to the deep sea has been the topic of several oceanographic programmes in the past 20 years in temperate seas (Walsh et al. 1988; Buscail et al. 1990; Biscaye et al. 1994; Blake et al. 1994). A general outcome of these studies was that the lateral input of biogenic detritus at the slope is usually so high that it causes an enhanced activity and abundance of benthic organisms at depth (Anderson et al. 1994; Kemp 1994; Rowe et al. 1994). Therefore, a major objective of several investigations of the relationship between benthic biomass, activity and carbon turnover in Arctic sediments was to evaluate the impact of shelf zones on food availability at Arctic margins and the deep sea (Vanaverbeke et al. 1997; Boetius and Damm 1998; Grebmeier 2003).
It has been hypothesized that such shelf–basin interactions might be particularly intense in the Arctic, as the deep central basins are nearly landlocked and surrounded by large marginal shelves, some of which are quite productive (Grebmeier 2003). In addition, some receive significant imports of freshwater and matter (a total of about 10% of the global river run-off), including terrigenic and fluvial carbon, from several major rivers, leading to a strong coupling between the terrestrial and marine environment, especially in the estuaries and shallow shelves but also in the entire Arctic in general (Macdonald 2000). The total freshwater discharge is large enough to generate the overall estuarine circulation of the Arctic Ocean and, hence, riverine and terrigenic carbon can be expected to contribute significantly to the possible export of organic material from the shelves to the central Arctic (Grebmeier 2003).
Evidence has been gathered that Arctic marginal shelves indeed export both dissolved and particulate organic carbon to the central basins (Stein and Macdonald 2003). However, it is still not clear of what nutritional quality and how significant the quantities of particulate organic carbon (POC) exports are. As the terrigenic and fluvial POC has already undergone significant biological degradation during its passage through the rivers and the coastal seas, their suitability as food is presumably rather low (Stein 1996; Fahl et al. 2001). Therefore, it has been hypothesized that these materials may be used to track processes involved in long-term burial in the sediments rather than be involved in the supply of usable organic matter to benthic communities (Klages et al. 2003). Christensen (1989) argued that the export of shelf POC depends very much on their width: narrow shelves tend to export a large proportion of their production while at wide ones most carbon is retained in the shelf system and only a rather small part is exported. The major Arctic shelves, such as the Barents Sea, the Siberian marginal seas, and the Chukchi Sea, are among the widest shelf regions in the world, exceeding up to 800 km from the coast to the shelf break. This fact suggests that most organic carbon may be trapped on the shelves and that the central Arctic Ocean may receive only a small magnitude of shelf or terrestrial carbon. Lisitsin (1995) provided evidence for this hypothesis, as he has shown that 90–95% of the suspended matter is deposited on the shelf of the Kara Sea south of 74°N. The Beaufort Sea shelf, however, is rather narrow and, hence, may well be a source of sediments, nutrients, organic matter and organisms exported to the Arctic Ocean (Grebmeier 2003).
Conclusions The most obvious progress in the past 15 years has been made in the scientific exploration of the deep ice-covered Arctic Ocean. There is now evidence that it is one order of magnitude more productive than previously thought. However, the role of shelf–basin interactions for the food supply of Arctic abyssal benthos and in the overall Arctic carbon cycle remains an open issue of current and future research.
Consequences of climate change
The effects of disturbance were among the research foci of a number of recent studies (see “Diversity, stability and disturbance”). A major disturbance agent, operating on a time scale of 10–1,000 years, is climate change, affecting life at virtually all ecologically relevant levels ranging from biochemical processes to ecosystem dynamics. Therefore, its consequences have been the overarching theme of most research programmes conducted in the past 15 years.
It is now commonly accepted that the Arctic Ocean is the region where changes in climate, hydrography and ecology, related to global warming, are expected to be strongly expressed and, hence, the Arctic can serve as a harbinger of global change (IPCC 2001; ACIA 2004). However, while there is no doubt that the Arctic climate is changing quite dramatically, the relative importance of natural variability (Macdonald et al. 1999; Vanegas and Mysak 2000) versus global warming due to anthropogenic greenhouse gas emissions is still debated (Johannessen et al. 1995; Shindell et al. 1999). The vast Arctic shelves are likely to be particularly sensitive because of their shallowness, their seasonally varying ice cover, and their dependence upon inflowing waters from the oceans and continents to the south. On the other hand, as just these Arctic systems are thought to be characterized by a variety of natural disturbances and have experienced pervasive environmental shifts in the past, they might be particularly well adapted to extreme climatic variations in terms of resilience (Dayton 1990).
Global change will not only lead to temperature increases but will also affect freshwater runoff of Arctic rivers (Peterson et al. 2002) and, hence, the proportions of freshwater in sea ice. In general, the Arctic Ocean is vertically stratified, largely due to sea-ice cover, with cold, less saline water on its surface and warmer, more saline water at depth (Aagaard et al. 1985). Recent studies have shown that deep Arctic Ocean temperatures have increased over the past 30 years (Dickson 1999). Warming will remove ice, whose annual cycle of melting and freezing structures the varying pan-Arctic rates of shelf exchange with the basin. Reduced sea-ice coverage will lead to lower cloud cover (due to less cold condensation), a lower albedo (which has in turn a positive feedback on sea-ice coverage), and possibly a higher primary production, resulting in a higher removal of atmospheric CO2 and increased sequestration of carbon on the Arctic shelves. In addition, a possible decrease in stratification would also bring warmer water to the surface. All these effects will surely give rise to large ecosystem changes.
Even given the proposed resilience and adaptability of Arctic systems in general (Dayton 1990), a climate change being so drastic that it results in a shift from a ‘cold/abundant ice’ to a ‘warm/limited ice’ mode will probably have profound ecological consequences propagating through all trophic levels, as sea-ice dynamics is the prime physical factor driving marine Arctic biology from cellular physiology and biochemistry to food web and habitat structure. For the eastern Arctic, it is hypothesized that upon warming the relative importance of sea-ice biota, pelagic communities and benthic assemblages will shift from a ‘benthos-dominated’ to a ‘zooplankton-dominated’ mode, which will fundamentally change the general pattern of kryo-pelago–benthic fluxes of matter and energy in Arctic seas. Accordingly, such a persistent change in the pelagic regime of primary and secondary production would most likely lead to profound changes in species composition, productivity and standing stocks of Arctic benthos and marine mammals. In general, there would be a shift in the relative importance of sea-ice, pelagic and benthic biota in the overall carbon flux from a ‘sea-ice algae–benthos’ to a ‘phytoplankton–zooplankton’ dominance (Carroll and Carroll 2003). Increased zooplankton grazing would result in a reduced flux of POC to the seabed and, consequently, decreased benthic biomass. The negative impact would be particularly intense for epifaunal megabenthic predators, such as gastropods, crabs, benthic and nektobenthic shrimps, sea stars, and probably brittle stars. Moreover, there is evidence that such a fundamental alteration will have a negative impact on large marine carnivores (seabirds and sea mammals) and will favour smaller carnivores (fish), because the average body size of prey species will decrease significantly (Karnovsky et al. 2003).
A similar shift is envisaged for the western Arctic, where unutilized shelf nutrients and dissolved organic matter (DOM) are currently exported to the deep Arctic Ocean (Walsh et al. 1997). In case of reduced exports and increased light availability on the shelves, primary production will probably increase, because of the enhanced nutrient uptake and recycling of the DOM, and this may lead to larger fish yields, with diversion of food away from the benthos and the higher trophic levels, as well as greater sequestration of atmospheric carbon dioxide in the shelf–basin sediments. Both effects will greatly impact Arctic human populations (because of the negative effects on ice-dependent seals and, consequently, on the future of seal-hunting indigenous people); the latter will even have global consequences (ACIA 2004).
Although studies on the ecological consequences of global change are generally regarded to be of high importance, their actual number is currently rather scarce. Because of the time scales involved, the response and adaptability of macrobenthic organisms to changing environmental conditions can hardly be examined directly and is, hence, usually deduced from time-series data of community parameters such as diversity, distribution patterns and composition of benthic assemblages. An example of this kind of studies is a comparison of epifaunal standing stocks in the southeastern Chukchi Sea (Feder et al. 2005). The results suggested that overall community structure did not change from 1976 to 1999, but some dominant taxa were clearly more abundant and had higher biomass in the recent surveys. However, findings like this should be interpreted with due caution, as they cannot unambiguously be related to regime shifts caused by the general warming of the Arctic (Feder et al. 2003). In contrast, there is good evidence from comparative studies that the diversity of Arctic fjord communities will decline in response to the climate-induced retreat of glaciers and the resultant increase in mineral sedimentation from melt waters (Wlodarska-Kowalczuk and Weslawski 2001).
Conclusions The study of the ecological effects of the pronounced climate change in the Arctic has been—and will remain—a major theme of recent, current and future research projects. The rapid warming will result in higher water temperatures, increased fluvial run-off and reduced ice cover—factors which will certainly give rise to severe ecosystem changes propagating through all trophic levels. According to a (yet) hypothetical scenario, the relative importance of sea-ice, pelagic and benthic biota in the overall carbon and energy flux will shift from a ‘sea-ice algae–benthos’ to a ‘phytoplankton–zooplankton’ dominance.
Possible directions of further research
As recommended in the reviews published between 1975 and 1991 (see “State of knowledge and research perspectives in the early 1990s”), major research themes in the past 15 years were the significance of pelago–benthic coupling, the quantification of benthic carbon fluxes and the modelling of their partitioning among different community fractions, the role of disturbances, and the ecological consequences of global change. Much has been achieved in the various research efforts, and a number of common notions on the Arctic benthos need to be modified due to the novel findings. However, not each scientific issue could be addressed with the same intensity and, as usual, each new result, each settled issue also produced new questions. For example, the wealth of data gained on benthic standing stocks and activities did indeed greatly extend our knowledge base but at the same time it revealed unequivocally a very uneven distribution of the values in space and time. This patchiness, which is pronounced on a variety of scales, strongly advises to take due caution in the up-scaling of point-measurement results. It clearly hampers the sound generalization of the results and has to be adequately taken into account in the budgeting and modelling of carbon fluxes.
With regard to future research issues, some of the recommendations of the former reviews are still effective. Winter data, for instance, are still rare, although more information about this part of the year is crucial to understand the adaptation of the benthic organisms to the extreme Arctic environment. The issue of the role of disturbances in the functioning of Arctic benthic ecosystems is also not fully settled. There is still a need to (1) quantify the various disturbance effects, (2) perform in situ experiments to monitor the response of benthic systems to natural or artificial disturbances (e.g., by following the succession of recolonization of disturbed areas; Jewett et al. 1999), and (3) eventually generate predictive models of disturbance effects and responses.
Closely intertwined with the disturbance topic are studies on diversity. It is now commonly accepted that a basic requirement for addressing the process-oriented issue of what key mechanisms regulate diversity is the sound determination of diversity itself, i.e., a thorough census of the species pool in a given area. Apart from this motivation, I expect that there will be more emphasis on taxonomic inventories in the future because of the increased awareness for the great significance and the threat to diversity in general. This idea has ignited several large-scale research initiatives, such as the ‘Census of Marine Life’ (O’Dor 2004) and the ‘Encyclopedia of Life’ (Wilson 2003), aiming at a complete inventory of the biodiversity of the world’s oceans and of the entire biosphere, respectively. In the Arctic, diversity-related work will include new surveys in remote and difficult-to-access areas and the increased use of modern molecular techniques for studies on taxonomy and population genetics.
Furthermore, I agree with Dayton’s (1990) concluding remark that sound autecological investigations (on reproduction, life cycles, growth, population dynamics, production, metabolic performance, and feeding) should not be neglected for the benefit of more work on processes and rates (as it has often been the case in the recent past). This should be so not only for its own sake but also because experience has taught us that bulk processes and rates can only be well understood (and modelled) if there is a sound knowledge on the basic natural history of the ecological key species involved.
Pelago–benthic coupling and carbon flow patterns have been and still are major issues of past and on-going research programmes. This will not change in the near future. A growing body of evidence has now been compiled that Arctic seas differ considerably in their environmental setting, hence in the strength of pelago–benthic coupling, and hence in benthic productivity and carbon flow. Consequently, it is generally recognized that a pan-Arctic approach is necessary to better comprehend the mechanisms responsible for the regional differences and to create an ample understanding of the benthic carbon utilization and cycling. Such a pan-Arctic effort should comprise a network of international co-ordinated collaborative studies in different seas around the Arctic, applying highly standardized methodological approaches to ensure highest possible data comparability, and an appropriate organizational framework to foster subsequent data integration and modelling work.
The response of Arctic benthos to global-change induced environmental regime shifts is another topic that will remain a research theme of great importance (ACIA 2004). To address this issue, more long-term observations are necessary to gain time-series data on environmental and faunal variations, if possible at a decadal time scale, which is most significant to mankind. Another approach to investigate possible ecological effects of global change are strictly comparative case studies in Arctic regions that are representative of different climate modes (‘cold/abundant ice’ vs. ‘warm/limited ice’). Process-oriented work on the issue of how and to what extent environmental changes affect benthic organisms and communities will focus on small-scale but multi-year investigations at selected sites in various habitats (coast, shelf, slope, and deep-sea). An example for such an approach is the “AWI Hausgarten", a deep-sea long-term station in the eastern Fram Strait at 2,600 m depth that was erected in 1999 (Soltwedel 1999). Latest progress in under-water and telecommunication technology might render possible that already in the near future the increasing use of autonomous and/or remotely operated research platforms, either mobile and stationary, will allow for the observation and measurement of global-change effects in Arctic seafloor habitats on a more long-term and cost-effective basis than nowadays conceivable.
Conclusions
The past two decades have witnessed a growing public awareness of the global climate significance of polar regions, as well as the opening of the Russian Arctic to foreign researchers. This, together with the considerable progress in ice-breaking technology, led to a pronounced intensification of multidisciplinary scientific research in Arctic seas, including numerous benthic field studies. The wealth of original data and information gathered in these efforts has markedly enhanced our knowledge on the Arctic benthos. While some scientific concepts have largely been corroborated by the novel findings (e.g., low endemism and high faunistic affinity to northern Atlantic assemblages), other common notions need to be—at least partly—revised, particularly with regard to the often-cited differences between Arctic seas and the Southern Ocean:
-
There is a growing body of evidence that benthos assemblages vary broadly in faunistic diversity between Arctic regions and that, hence, the idea of a consistently very poor Arctic benthos—being in stark contrast to the rich Antarctic bottom fauna—is an undue overgeneralization. At a global scale, in terms of biogeographical diversity, both Arctic and Antarctic waters seem to be characterized by intermediate species richness.
-
Levels of disturbance—a major ecological agent known to heavily affect benthic diversity and community structure—have been assumed to be relatively high in the Arctic but exceptionally low in the Southern Ocean. The discovery of the great role of iceberg scouring in Antarctic shelf ecosystems, which has largely been overlooked in the past, calls for a reconsideration of this notion.
-
The novel data clearly demonstrate that there are marked differences in geographical and environmental setting, impact of fluvial run-off, pelagic production regime, strength of pelago–benthic coupling and, hence, food supply to the benthos among the various Arctic seas, impeding the large-scale generalization of local and regional findings.
-
Field evidence strongly indicates the great significance of meso-scale hydrography and ice cover (marginal ice zones, polynyas, and gyres) in forming ‘hot spots’ of tight pelago–benthic coupling and, hence, high benthic biomass.
-
In contrast, the importance of the huge amounts of terrigenic organic matter discharged to the Arctic seas through fluvial run-off as an additional food source for the benthos is still under debate.
-
Studies on the partitioning of energy flow through the benthic communities strongly suggested that the often-overlooked megafauna has to be adequately considered in overall benthic energy budgets and models of carbon cycling, particularly in Arctic shelf systems that are commonly dominated by abundant echinoderm populations.
-
The most obvious progress has been made in the scientific exploration of the deep ice-covered Arctic Ocean. There is now evidence that it is one order of magnitude more productive than previously thought. Therefore, the significance of shelf–basin interactions, i.e., the importance of excess organic carbon exported from productive shelf areas to the deep sea of the interior ocean, is still debated and, hence, a major topic of on-going research projects.
-
Another high-priority theme of current—and future—research projects are the ecological consequences of the rapid warming in the Arctic. Higher water temperatures, increased fluvial run-off and reduced ice cover will give rise to severe ecosystem changes propagating through all trophic levels. It is hypothesized that there would be a shift in the relative importance of sea-ice, pelagic and benthic biota in the overall carbon and energy flux, ultimately resulting in a switch from a ‘sea-ice algae–benthos’ to a ‘phytoplankton–zooplankton’ dominance.
-
Besides the process-oriented studies on shelf–basin interactions and climate-change effects, possible routes of further research should encompass:
-
a continuation of the work on the role of disturbance in the functioning of Arctic–and Antarctic–ecosystems,
-
a large-scale census of the Arctic species pool,
-
autoecological studies of key species, and
-
the acquisition of long-term time-series information, including winter data.
-
The latter research work will be strongly facilitated by the upcoming availability of autonomous or remotely operated in situ research platforms.
References
Aagaard K, Swift JH, Carmack EC (1985) Thermohaline circulation in the Arctic Mediterranean seas. J Geophys Res 90:4833–4846
Ambrose Jr WG, Clough LM, Tilney PR, Beer L (2001) Role of echinoderms in benthic remineralization in the Chukchi Sea. Mar Biol 139:937–949
Ambrose Jr WG, Renaud PE (1995) Benthic response to water column productivity patterns: evidence for benthic-pelagic coupling in the Northeast Water Polynya. J Geophys Res 100:4411–4421
Anderson LG, Björk G, Holby O, Kattner G, Koltermann PK, Jones EP, Liljeblad B, Lindegren R, Rudels B, Swift JH (1994) Water masses and circulation in the Eurasian Basin: results from the Oden 91 North Pole expedition. J Geophys Res 99:3273–3283
Andreassen I, Nöthig E-M, Wassmann P (1996) Vertical particle flux on the shelf off northern Spitsbergen, Norway. Mar Ecol Prog Ser 137:215–228
Anisimova NA (1989) Distributional patterns of echinoderms in the Eurasian sector of the Arctic Ocean. In: Herman Y (ed) The Arctic seas. Van Nostrand, New York, pp 281–301
Anonymous (1991) Mission Statement and Core Programme for the International Arctic Polynya Programme. Joint Oceanographic Institutions, Washington, D.C., 10 pp
Arctic Climate Impact Assessment (ACIA) (2004) Impacts of a warming Arctic. Cambridge University Press, Cambridge, 139 pp
Arntz WE (1995) Antarktisches Benthos: Mythen und Daten. In: Hempel I, Hempel G (eds) Biologie der Polarmeere. Gustav Fischer, Jena, pp 230–239
Arntz WE, Brey T, Gallardo VA (1994) Antarctic zoobenthos. Oceanogr Mar Biol Annu Rev 32:241–304
Arntz WE, Gutt J, Klages M (1997) Antarctic marine biodiversity: an overview. In: Battaglia B, Valencia J, Walton DWH (eds) Antarctic communities: species, structure and survival. Cambridge University Press, Cambridge, pp 3–14
Aronson RB (1989) Brittlestar beds: low-predation anachronisms in the British Isles. Ecology 70:856–865
Ashjian CJ, Smith SL, Bignami F, Hopkins T, Lane PVZ (1997) Distribution of zooplankton in the Northeast Water Polynya during summer 1993. J Mar Syst 10:279–298
Ashjian CJ, Smith SL, Lane PVZ (1995) The Northeast Water Polynya during summer 1992: distribution and aspects of secondary production of copepods. J Geophys Res 100:4371–4388
Bauerfeind E, Bodungen Bv, Arndt K, Koeve W (1994) Particle flux and composition of sedimenting matter in the Greenland Sea. J Mar Syst 5:411–423
Biscaye PE, Flagg CN, Falkowski PG (1994) The shelf edge exchange processes experiment, SEEP-II: an introduction to hypotheses, results and conclusions. Deep-Sea Res II 41:231–252
Blake JA, Diaz RJ (1994) Input, accumulation and cycling of materials on the continental slope off Cape Hatteras: an introduction. Deep-Sea Res II 41:707–710
Bleil U, Thiede J (1990) The geological history of Cenozoic polar oceans: Arctic versus Antarctic—an introduction. In: Bleil U, Thiede J (eds) Geological history of Cenozoic polar oceans: Arctic versus Antarctic. Kluwer, Dordrecht, pp 1–8
Bluhm BA, MacDonald IR, Debenham C, Iken K (2005) Macro- and megabenthic communities in the high Arctic Canada Basin: initial findings. Polar Biol 28:218–231
Bluhm BA, Piepenburg D, Juterzenka Kv (1998) Distribution, standing stock, growth, mortality and production of Strongylocentrotus pallidus (Echinodermata: Echinoidea) in the northern Barents Sea. Polar Biol 20:325–334
Boetius A, Damm E (1998) Benthic oxygen uptake, hydrolytical potentials and microbial biomass at the Arctic continental slope. Deep-Sea Res I 45:239–275
Bourke RH, Newton JL, Paquette RG, Tunnicliffe MD (1987) Circulation and water masses of the East Greenland Shelf. J Geophys Res 92:6729–6740
Brandt A (1995) Peracarid fauna (Crustacea, Malacostraca) of the Northeast Water Polynya off Greenland: documenting close benthic-pelagic coupling in the Westwind Trough. Mar Ecol Prog Ser 121:39–51
Brey T, Clarke A (1993) Population dynamics of marine benthic invertebrates in Antarctic and subantarctic environments: are there unique adaptations? Antarct Sci 5:253–266
Buscail R, Pocklington R, Daumas R, Guidi L (1990) Fluxes and budget of organic matter in the benthic boundary layer over the northwestern Mediterranean margin. Cont Shelf Res 10:1089–1122
Carey Jr AG (1991) Ecology of North American Arctic continental shelf benthos: a review. Cont Shelf Res 11:865–883
Carney RS, Haedrich RL, Rowe GT (1983) Zonation of fauna in the deep sea. In: Rowe GT (ed) The sea, vol. 8: Deep-sea biology. Wiley, New York, pp 371–398
Carroll MK, Carroll J (2003) The Arctic seas. In: Black KD, Shimmield GB (eds) Biogeochemistry of Marine Systems. Blackwell, Sheffield, pp 127–156
Cauwet G, Sidorov I (1996) The biogeochemistry of Lena River: organic carbon and nutrients distribution. Mar Chem 53:211–227
Cavalieri DJ, Gloersen P, Parkinson CL, Comiso JC, Zwally HJ (1997) Observed hemispheric asymmetry in global sea ice changes. Science 278:1104–1106
Christensen JP (1989) Sulfate reduction and carbon oxidation rates in continental shelf sediments, an examination of off-shelf carbon transport. Cont Shelf Res 9:223–246
Christensen JP (2000) A relationship between deep-sea benthic oxygen demand and oceanic primary productivity. Oceanologica Acta 23:65–82
Clarke A (1983) Life in cold water: the physiological ecology of polar marine ectotherms. Oceanogr Mar Biol Annu Rev 21:341–453
Clarke A (1994) Temperature and evolution: southern Ocean cooling and the Antarctic marine fauna. In: Kerry KR, Hempel G (eds) Antarctic ecosystems—ecological change and conservation. Springer, Berlin Heidelberg New York, pp 9–22
Clarke A (2003) Evolution, adaptation and diversity: global ecology in an Antarctic context. in: Huikes AHL, Gieskes WWC, Rozema J, Schorno RML, van der Vries SM, Wolff WJ (eds) Antarctic biology in a global context. Backhuys Publishers, Leiden, pp 3–17
Clarke A, Crame JA (1989) The origin of the Southern Ocean marine fauna. In: Crame JA (ed) Geological Society Special Publication, Vol. 47: Origins and evolution of the Antarctic biota. Geological Society, London, pp 253–268
Clarke A, Johnston NM (2003) Antarctic marine benthic diversity. Oceanogr Mar Biol Annu Rev 41:47–114
Clough LM, Ambrose Jr WG, Ashjian CJ, Piepenburg, D, Renaud PE, Smith SL (1997a) Meroplankton abundance in the Northeast Water Polynya: insights from oceanographic parameters and benthic abundance patterns. J Mar Syst 10:343–357
Clough LM, Ambrose Jr WG, Cochran JK, Barnes C, Renaud PE, Aller RC (1997b) Infaunal density, biomass and bioturbation in the sediments of the Arctic Ocean. Deep-Sea Res 44:1683–1704
Conlan KE, Lenihan HS, Kvitek RG, Oliver JS (1998) Ice scour disturbance to benthic communities in the Canadian High Arctic. Mar Ecol Prog Ser 166:1–16
Cooper LW, Grebmeier JM, Larsen IL, Egorov VG, Theodorakis C, Kelly HP, Lovvorn JR (2002) Seasonal variation in sedimentation of organic materials in the St. Lawrence Island polynya region, Bering Sea. Mar Ecol Prog Ser 226:13–26
Curtis MA (1975) The marine benthos of Arctic and sub-Arctic continental shelves. A review of regional studies and their general results. Polar Rec 17:595–626
Dahm C (1996) Ökologie und Populationsdynamik antarktischer Ophiuroiden (Echinodermata). Ber Polarforsch 194:1–289
Dayton PK (1984) Processes structuring some marine communities: are they general? In: Strong DR, Simberloff D, Lawrence GA, Thistle AB (eds) Ecological communities: conceptual issues and the evidence. Princeton University Press, Princeton, pp 181–197
Dayton PK (1990) Polar benthos. In: Smith Jr WO (ed) Polar oceanography, Part B: Chemistry, biology, and geology. Academic, San Diego, pp 631–685
Dayton PK, Oliver JS (1977) Antarctic soft-bottom benthos in oligotrophic and eutrophic environments. Science 197:55–58
Dayton PK, Tegner MJ (1984) The importance of scale in community ecology: a kelp forest example with terrestrial analogs. In: Price PW, Slobodchikoff CN, Gaud WS (eds) A new ecology: novel approaches to interactive systems. Wiley, New York, pp 457–481
Deming J, Fortier L, Fukuchi M (2002) The International North Water Polynya Study (NOW): a brief overview. Deep-Sea Res II 49:4887–4892
Deubel H (2000) Struktureigenschaften und Nahrungsbedarf der Zoobenthosgemeinschaften im Bereich des Lomonssowrückens im Arktischen Ozean. Ber Polarforsch 370:1–147
Deubel H, Engel M, Fetzer I, Gagaev S, Hirche HJ, Klages M, Larionov V, Lubin P, Lubina O, Nöthig EM, Okolodkov Y, Rachor E (2003) The Kara Sea ecosystem: phytoplankton, zooplankton and benthos communities influenced by river run-off. In: Stein R, Fahl K, Fütterer DK, Galimov EM, Stepanets OV (eds) Siberian river run-off in the Kara Sea: characterisation, quantification, variability, and environmental significance. Elsevier, Amsterdam, pp 237–266
Dickson RR (1999) All change in the Arctic. Nature 397:389–391
Dickson RR, Osborn TJ, Hurrell JW, Meincke J, Blindheim J, Adlandsvik B, et al (2000) The Arctic Ocean response to the North Atlantic Oscillation. J Clim 13:2671–2696
Dunbar MJ (1977) The evolution of polar ecosystems. In: Llano GA (ed) Adaptations within the Antarctic ecosystems. Smithsonian Institution, Washington, pp 1063–1076
Dunton K (1992) Arctic biogeography: the paradox of the marine benthic fauna and flora. Trends Ecol Evol 7:183–189
D’yakonov AM (1967) Ophiuroids of the USSR seas. Israel Program for Scientific Translations, Jerusalem, 123 pp
Fahl K, Cremer H, Erlenkeuser H, Hanssen H, Hölemann J, Kassens H, Knickmeier K, Kosobokova K, Kunz-Pirrung M, Lindemann F, Markhaseva E, Lischka S, Petryashov V, Piepenburg D, Schmid MK, Spindler M, Stein R, Tuschling K (2001) Sources and pathways of organic carbon in the modern Laptev Sea (Arctic Ocean): implications from biological, geochemical and geological data. Polarforschung 69:193–205
Feder HM, Foster NR, Jewett SC, Weingartner TJ, Baxter R (1994a) Mollusks in the Northeastern Chukchi Sea. Arctic 47: 145–163
Feder HM, Jewett CJ, Blanchard A (2005) Southeastern Chukchi Sea (Alaska) epibenthos. Polar Biol 28:402–421
Feder HM, Naidu AS, Jewett SC, Hameedi JM, Johnson WR, Whitledge TE (1994b) The north-eastern Chukchi Sea: benthos-environmental interactions. Mar Ecol Prog Ser 111:171–190
Feder HM, Norton DW, Geller JB (2003) A review of apparent 20th century changes in the presence of mussels (Mytilus trossulus) and macroalgae in Arctic Alaska, and of historical and paleontological evidence used to relate mollusc distributions to climate change. Arctic 56:391–407
Gage JD, Tyler PA (1991) Deep-sea biology: a natural history of organisms at the deep-sea floor. Cambridge University Press, Cambridge
George RY (1977) Dissimilar and similar trends in antarctic and arctic marine benthos. In: Dunbar MJ (ed) Polar oceans. Arctic Institute of North America, Calgary, pp 391–408
Golikov AN (1990) Explorations of the fauna of the seas 37(45): ecosystems of the New Siberian shoals and the fauna of the Laptev Sea and adjacent waters of the Arctic Ocean. Nauka, Moscow
Gosselin M, Levasseur M, Wheeler PA, Horner RA, Booth BC (1997) New measurements of phytoplankton and ice algal production in the Arctic Ocean. Deep-Sea Res II 44:1623–1644
Gould PJ, Forsell DJ, Lensink CJ (1982) Pelagic distribution and abundance of seabirds in the Gulf of Alaska and eastern Bering Sea. US Fish Wildlife Ser FWS/OBS-82/48, 294 pp
Gradinger R (1995) Climate change and biological oceanography of the Arctic Ocean. Phil Trans R Soc Lond A 352:277–286
Gradinger R, Baumann MEM (1991) Distribution of phytoplankton communities in relation to the large-scale hydrographical regime in the Fram Strait. Mar Biol 111:311–321
Graf G (1989) Benthic-pelagic coupling in a deep-sea benthic community. Nature 341:437–439
Graf G (1992) Benthic-pelagic coupling: a benthic view. Oceanogr Mar Biol Annu Rev 30:149–190
Graf G, Gerlach SA, Linke P, Queisser W, Ritzrau W, Scheltz A, Thomsen L, Witte U (1995) Benthic-pelagic coupling in the Greenland-Norwegian Sea and its effect on the geological record. Geol Rundschau 84:49–58
Grassle JF (1986) The ecology of deep-sea hydrothermal vent communities. Adv Mar Biol. 23:301–362
Gray JS (1981) The ecology of marine sediments. Cambridge University Press, New York, 185 pp
Gray JS (2001) Antarctic marine benthos biodiversity in a world-wide latitudinal context. Polar Biol 24:633–641
Grebmeier JM (1993) Studies of pelagic-benthic coupling extended onto the Soviet continental shelf in the northern Bering and Chukchi seas. Cont Shelf Res 13:653–668
Grebmeier JM (2003) The western Arctic shelf-basin interactions project. Arctic Research of the United States 17:24–32
Grebmeier JM, Barry JP (1991) The influence of oceanographic processes on pelagic-benthic coupling in polar regions: a benthic perspective. J Mar Syst 2: 495–518
Grebmeier JM, Cooper JP (1995) Influence of the St. Lawrence Island Polynya upon the Bering Sea benthos. J Geophys Res 100:3349–4460
Grebmeier JM, McRoy CP (1989) Pelagic-benthic coupling on the shelf of the northern Bering and Chukchi Seas. III. Benthic food supply and carbon cycling. Mar Ecol Prog Ser 53:79–91
Grebmeier JM, Smith WO, Conover RB (1995) Biological processes on Arctic continental shelves: ice-ocean-biotic interactions. In: Smith WO, Grebmeier JM (eds) Arctic oceanography: marginal zones and continental shelves. American Geophysical Union, Washington, D.C., pp 231–261
Gukov AY (1995) Hydrobiological research in the Lena polynya. Ber Polarforsch 176:228–229
Gutt J (1991) On the distribution and ecology of holothurians in the Weddell Sea (Antarctica). Polar Biol 11:145–155
Gutt J (2001) On the direct impact of ice on marine benthic communities, a review. Polar Biol 24:553–564
Gutt J, Piepenburg D (2003) Scale-dependent impact on diversity of Antarctic benthos caused by grounding of icebergs. Mar Ecol Prog Ser 253:77–83
Gutt J, Sirenko BI, Smirnov IS, Arntz WE (2004) How many macrozoobenthic species might inhabit the Antarctic shelf? Antarct Sci 16:11–16
Haedrich RL, Rowe GT, Polloni PT (1980) The megabenthic fauna in the deep sea south of New England, USA. Mar Biol 57:165–179
Hargrave BT (1973) Coupling carbon flow through some pelagic and benthic communities. J Fish Res Board Can 30:1317–1326
Hebbeln D, Wefer G (1991) Effects of ice coverage and ice-rafted material on sedimentation in the Fram Strait. Nature 350:409–411
Hedgpeth JW (1977) The Antarctic marine ecosystem. In: Llano GA (ed) Adaptations within Antarctic ecosystems. Smithsonian Institution, Washington DC, pp 3–10
Heding SG (1942) Holothurioidea. Part II. Aspidochirota – Elasipoda – Dendrochirota. Danish Ingolf-Expedition 4:43–78
Hempel G (1985) On the biology of polar seas, particularly the Southern Ocean. In: Gray JS, Christiansen ME (eds) Marine biology of polar regions and the effect of stress on marine organisms. Wiley, Chichester, pp 3–34
Herman Y (1989) The Arctic seas. Van Nostrand Reinhold, New York, 888 pp
Hessler RR, Jumars PA (1974) Abyssal community analysis from replicate box cores in the central North Pacific. Deep-Sea Res 21:185–209
Highsmith RC, Coyle KO (1990) High productivity of northern Bering Sea benthic amphipods. Nature 344:862–864
Highsmith RC, Coyle KO (1992) Productivity of Arctic amphipods relative to gray whale energy requirements. Mar Ecol Prog Ser 83:141–150
Hirche H-J, Hagen W, Mumm N, Richter C (1994) The Northeast Water Polynya, Greenland Sea. III. Meso- and macrozooplankton distribution and production of dominant herbivorous copepods during spring. Polar Biol 14:491–503
Hirche H-J, Kattner G (1994) The 1993 Northeast water expedition. Scientific cruise report of RV “Polarstern" Arctic cruises ARK-IX/2 and 3, USCG “Polar Sea" cruise NEWP and the NEWLand expedition. Ber Polarforsch 142:1–190
Hirche H-J, Kwasniewski S (1997) Distribution, reproduction and development of Calanus species in the Northeast Water in relation to environmental conditions. J Mar Syst 10:299–317
Hobson KA, Ambrose Jr WG, Renaud PE (1995) Sources of primary production, benthic-pelagic coupling, and trophic relationships within the Northeast Water Polynya: insights from δ13C and δ15N analysis. Mar Ecol Prog Ser 128:1–10
Honjo S (1990) Particle fluxes and modern sedimentation in the polar oceans. In: Smith Jr WO (ed) Polar oceanography, Part B: Chemistry, biology, and geology. Academic, San Diego, pp 687–734
Hop H, Pearson T, Hegseth EN, Kovacs KM, Wiencke C, Kwasniewski S, Eiane K, Mehlum F, Gulliksen B, Wlodarska-Kowalczuk M, Lydersen C, Weslawski JM, Cochrane S, Gabrielsen GW, Leakey RJG, Lønne OJ, Zajaczkowski M, Falk-Petersen S, Kendall M, Wängberg S-Å, Bischof K, Voronkov AY, Kovaltchouk NA, Wiktor J, Poltermann M, di Prisco G, Papucci C, Gerland S (2002) The marine ecosystem of Kongsfjorden, Svalbard. Polar Res 21:167–208
Hulth S, Blackburn TH, Hall POJ (1994) Arctic sediments (Svalbard)—consumption and microdistribution of oxygen. Mar Chem 46:293–316
Hurlbert SH (1971) The non-concept of species diversity: a critique and alternative parameters. Ecology 52:577–586
Huston M (1979) A general hypothesis of species diversity. Am Nat 113:81–101
Intergovernmental Panel on Climate Change (IPCC) (2001) Climate Change 2000. Third assessment report, Cambridge University Press
Jakobsson M, Løvlie R, Arnold EM, Backman J, Polyak L, Knutsen JO, and Musatov E (2001) Pleistocene stratigraphy and paleoenvironmental variation from Lomonossov Ridge sediments, central Arctic Ocean. Glob Planet Change 31:1–21
Jewett SC, Feder HM (1981) Epifaunal invertebrates of the continental shelf of the eastern Bering and Chukchi Seas. In: Hood DW, Calder JA (eds) The eastern Bering Sea shelf, Vol. II: Oceanography and resources. Univ. Washington Press, Seattle, pp 1131–1153
Jewett SC, Feder HM, Blanchard A (1999) Assessment of the benthic environment following offshore placer gold mining in the northeastern Bering Sea. Mar Env Res 48:91–122
Johannessen OM, Miles M, Bjorgo E (1995) The Arctic’s shrinking sea ice. Nature 376:126–127
Karnovsky N, Kwasniewski S, Weslawski JM, Walkusz W, Beszczynska-Moller A (2003) Foraging behaviour of little auks in a heterogeneous environment. Mar Ecol Prog Ser 253:289–303
Kassens H, Bauch HA, Dmitrenko IA, Eicken H, Hubberten H-W, Melles M, Thiede J, Timokhov LA (eds) (1999) Land-ocean systems in the Siberian Arctic: Dynamics and history. Springer, Berlin Heidelberg New York, 711 pp
Kemp PF (1994) Microbial carbon utilization on the continental shelf and slope during the SEEP-II experiment. Deep-Sea Res II 41:563–581
Kendall MA (1996) Are Arctic soft-sediment macrobenthic communities impoverished? Polar Biol 16:393–399
Kendall MA, Aschan M (1993) Latitudinal gradients in the structure of macrobenthic communities: a comparison of Arctic, temperate and tropical sites. J Exp Mar Biol Ecol 172:157–169
Kendall MA, Widdicombe S, Weslawski JM (2003) A multi-scale study of the biodiversity of the benthic infauna of the high-latitude Kongsfjord, Svalbard. Polar Biol 26:383–388
Klages M, Boetius A, Christensen JP, Deubel H, Piepenburg D, Schewe I, Soltwedel T (2003) The benthos of Arctic Seas and its role for the carbon cycle at the seafloor. in: Stein R, Macdonald RW (eds) The organic carbon cycle in the Arctic Ocean: present and past. Springer, Berlin Heidelberg New York, pp 139–167
Klages M, Vopel K, Bluhm H, Brey T, Soltwedel T, Arntz WE (2001) Deep-sea food falls: first observation of a natural event in the Arctic Ocean. Polar Biol 24:292–295
Knox GA, Lowry JK (1977) A comparison between the benthos of the Southern Ocean and the North Polar Ocean, with special reference to the Amphipoda and the Polychaeta. In: Dunbar MJ (ed) Polar oceans. Arctic Institute of North America, Calgary, pp 423–462
Kröncke I (1994) Macrobenthos composition, abundance and biomass in the Arctic Ocean along a transect between Svalbard and the Makarov Basin. Polar Biol 14:519–529
Kröncke I (1998) Macrofauna communities in the Amundsen Basin, at the Morris Jesup Rise and at the Yermak Plateau (Eurasian Arctic Ocean). Polar Biol 19:383–392
Kröncke I, Vanreusel A, Vincx M, Wollenburg J, Mackensen A, Liebezeit G, Behrends B (2000) Different benthic size-compartments and their relationship to sediment chemistry in the deep Eurasian Arctic Ocean. Mar Ecol Prog Ser 199:31–41
Kvitek RG, Conlan KE, Iampietro PJ (1998) Black pools of death: Hypoxic, brine-filled ice gouge depressions become lethal traps for benthic organisms in a shallow Arctic embayment. Mar Ecol Prog Ser 162:1–10
Lampitt RS, Billett DSM, Rice AL (1986) Biomass of the invertebrate megabenthos from 500 to 4100 m in the northeast Atlantic Ocean. Mar Biol 93:69–81
Legendre L, Ackley SF, Dieckmann GS, Gulliksen B, Horner R, Hoshiai T, Melnikov IA, Reeburgh WS, Spindler M, Sullivan CW (1992) Ecology of sea ice biota. 2. Global significance. Polar Biol 12:429–444
Levin LA, Gooday AJ (2003) The deep Atlantic Ocean. In: Tyler PA (ed) Ecosystems of the world, vol. 28: Ecosystems of the deep sea. Elsevier, Amsterdam, pp 111–178
Lisitsin AP (1995) The marginal filter of the ocean. Oceanology 34:583–590
Loeng H (1989) Ecological features of the Barents Sea. In: Rey L, Alexander V (eds) Proceedings of the 6th conference of the Comité Arctique International. Brill, Leiden, pp 327–365
Lozán JL, Graßl H, Hupfer P (2001) Climate of the 21st century: changes and risks. Wissenschaftliche Auswertungen, Hamburg, 449 pp
Macdonald RW (1996) Awakenings in the Arctic. Nature 380:286–287
Macdonald RW (2000) Arctic estuaries and ice: a positive-negative estuarine couple. In: Lewis EL (ed) The freshwater budget of the Arctic Ocean. NATO ASI Series, pp 383–407
Macdonald RW, Carmack EC (1991) Age of Canada Basin deep waters: a way to estimate primary production for the Arctic Ocean. Science 254:1348–1350
Macdonald RW, Carmack EC, McLaughlin FA, Falkner KK, Swift JH (1999) Connections among ice, runoff and atmospheric forcing in the Beaufort Gyre. Geophys Res Lett 26:2223–2226
Macdonald RW, Carmack EC, Wallace DWR (1993) Tritium and radiocarbon dating of Canada Basin deep waters. Science 259:103–104
MacGinitie GE (1955) Distribution and ecology of the marine invertebrates of Point Barrow, Alaska. Smithsonian Miscellaneous Collections 128:1–201
Magurran AE (1988) Ecological diversity and its measurement. Chapman and Hall, New York, 179 pp
Martin JH, Knauer GA, Karl DM (1987) Vertex: carbon cycling in the northeast Pacific. Deep-Sea Res 34:267–285
McRoy CP (1993) ISHTAR, the project: an overview of Inner Shelf Transfer and Recycling in the Bering and Chukchi Seas. Cont Shelf Res 13:473–479
McRoy CP, Hood DW, Coachman LK, Walsh JJ, Goering J (1986) Processes and resources of the Bering Sea (PROBES): the development and accomplishments of the project. Cont Shelf Res 5:5–21
Muus K (1981) Density and growth of juvenile Amphiura filiformis (Ophiuroidea) in the Øresund. Ophelia 20:153–168
Myhre AM, Thiede J, Firth JV et al. (1995) Proceedings Of the Ocean Drilling Program (Initial reports) 151. College Station, Texas, pp 1–926
Niebauer HJ (1991) Bio-physical oceanographic interactions at the edge of the Arctic pack ice. J Mar Syst 2:209–232
O’Dor R (2004) A census of marine life. BioScience 54:92–93
Paterson GLJ (1985) The deep-sea Ophiuroidea of the North Atlantic Ocean. Bull Brit Mus Nat Hist (Zool) 49:1–162
Paul AZ, Menzies RJ (1974) Benthic ecology of the high Arctic deep sea. Mar Biol 27:251–262
Peck LS, Brockington S, Vanhove S, Beghyn M (1999) Community recovery following catastrophic iceberg impacts in a soft-sediment shallow-water site at Signy Island, Antarctica. Mar Ecol Prog Ser 186:1–8
Petersen GH (1984) Energy flow in comparable aquatic ecosystems from different climatic zones. Rapp P-v Réun Cons int Explor Mer 183:119–125
Petersen GH, Curtis MA (1980) Differences in energy flow through major components of subarctic, temperate and tropical marine shelf ecosystems. Dana 1:53–64
Peterson BJ, Holmes RM, McClelland JW, Vorosmarty CJ, Lammers RB, Shiklomanov AI, Shiklomanov IA, Rahmstorf S (2002) Increasing river discharge to the Arctic Ocean. Science 298:2171–2173
Pickett STA, White PS (eds) (1985) The ecology of natural disturbance and patch dynamics. Academic Press, Orlando, 472 pp
Piepenburg D (1995) “Glasnost'' in der arktischen Benthosforschung: Politisches Tauwetter im sibirischen Nordpolarmeer. In: Hempel I, Hempel G (eds) Biologie der Polarmeere. Gustav Fischer, Jena, pp 240–247
Piepenburg D (2000) Arctic brittle stars (Echinodermata: Ophiuroidea). Oceanogr Mar Biol Annu Rev 38:189–256
Piepenburg D, Blackburn TH, Dorrien CF, Gutt J, Hall POJ, Hulth S, Kendall MA, Opalinski KW, Rachor E, Schmid MK (1995) Partitioning of benthic community respiration in the Arctic (northwestern Barents Sea). Mar Ecol Prog Ser 118:199–213
Piepenburg D, Brandt A, Juterzenka K v, Mayer M, Schnack K, Seiler D, Witte U, Spindler M (2001) Patterns and determinants of the distribution and structure of benthic faunal assemblages in the northern North Atlantic. In: Schäfer P, Ritzrau W, Schlüter M, Thiede J (eds) The northern North Atlantic: a changing environment. Springer, Berlin Heidelberg New York, pp 179–19
Piepenburg D, Schmid MK (1996a) Brittle star fauna (Echinodermata: Ophiuroidea) of the Arctic northwestern Barents Sea: composition, abundance, biomass and spatial distribution. Polar Biol 16:383–392
Piepenburg D, Schmid MK (1996b) Distribution, abundance, biomass, and mineralization potential of the epibenthic megafauna of the Northeast Greenland shelf. Mar Biol 125:321–332
Piepenburg D, Schmid MK (1997) A photographic survey of the epibenthic megafauna of the Arctic Laptev Sea shelf: distribution, abundance, and estimates of biomass and organic carbon demand. Mar Ecol Prog Ser 147:63–75
Piepenburg D, Voß J, Gutt J (1997) Assemblages of sea stars (Echinodermata: Asteroidea) and brittle stars (Echinodermata: Ophiuroidea) in the Weddell Sea (Antarctica) and off Northeast Greenland (Arctic): A comparison of diversity and abundance. Polar Biol 17:305–322
Polyak L, Forman SL, Herlihy FA, Ivanov G, Krinitsky P (1997) Late Weichselian deglacial history of the Svyataya (Saint) Anna Trough, northern Kara Sea, Arctic Russia. Mar Geol 143:169–188
Rachor E (1992) Scientific cruise report of the 1991 Arctic Expedition ARK-VIII/2 of RV “POLARSTERN" (EPOS II: Study of the European Arctic Shelf, “SEAS", of the European Science Foundation. Ber Polarforsch 115:1–150
Rey F, Loeng H (1985) The influence of ice and hydrographic conditions on the development of phytoplankton in the Barents Sea. In: Gray JS, Christiansen ME (eds) Marine biology of polar regions and effects of stress on marine organisms. Wiley, Chichester, pp 49–63
Rothrock DA, Yu Y, Maykut GA (1999) Thinning of the Arctic sea-ice cover. Geophys Res Lett 26:3469–3472
Rowe GT, Boland GS, Phoel WC, Anderson RF, Biscaye PE (1994) Deep-sea respiration as an indication of lateral input of biogenic detritus from continental margins. Deep-Sea Res II 41:657–668
Rowe GT, Polloni PT, Horner SG (1974) Benthic biomass estimates from the northwestern Atlantic Ocean and the northern Gulf of Mexico. Deep-Sea Res I 21:641–650
Sakshaug E (2003) Primary and secondary production in the Arctic seas. In: Stein R, Macdonald RW (eds) The organic carbon cycle in the Arctic Ocean: present and past. Springer, Berlin Heidelberg New York, pp 57–82
Sakshaug E, Skjodal HR (1989) Life at the ice edge. Ambio 18:60–67
Salzwedel H, Rachor E, Gerdes D (1985) Benthic macrofauna communities in the German Bight. Veröff Inst Meeresforsch Bremerh 20:199–267
Sanders HL (1968) Marine benthic diversity: a comparative study. Am Nat 102:243–282
Savidge G, Priddle J, Gilpin LC, Bathmann U, Murphy EJ, Owens NJP, Pollard RT, Turner DR, Veth C, Boyd P (1996) An assessment of the role of the marginal ice zone in the carbon cycle of the Southern Ocean. Antarct Sci 8:349–358
Schäfer P, Ritzrau W, Schlüter M, Thiede J (2001) The northern North Atlantic: a changing environment. Springer, Berlin Heidelberg New York, 500 pp
Schewe I, Soltwedel T (1999) Deep-sea meiobenthos of the central Arctic Ocean: distribution patterns and size-structure under extreme oligotrophic conditions. Vie et Milieu 49:79–92
Schewe I, Soltwedel T (2003) Benthic responses to ice-edge-induced particle flux in the Arctic Ocean. Polar Biol 26:610–620
Schmid MK, Piepenburg D, Kassens H, Abramova EN, Gukov AY, Petryashov VV (2005) Trophic dynamics and carbon flux in the Laptev Sea. Prog Oceanogr (in prep.)
Schneider W, Budéus G (1994) The North East Water Polynya (Greenland Sea). I. A physical concept of its generation. Polar Biol 14:1–9
Sejr MK, Jensen KT, Rysgaard S (2000) Macrozoobenthic community structure in a high-arctic East Greenland fjord. Polar Biol 23:792–801
Sibuet M, Olu K (1998) Biogeography, biodiversity and fluid dependence of deep-sea cold-seep communities at active and passive margins. Deep-Sea Res II 45:517–567
Sirenko BI (2001) List of species of free-living invertebrates of Eurasian Arctic seas and adjacent deep waters. Explor Fauna Seas 51(59):1–76
Sirenko BI, Piepenburg D (1994) Current knowledge on biodiversity and benthic zonation patterns of Eurasian Arctic shelf seas, with special reference to the Laptev Sea. Ber Polarforsch 144:69–77
Shindell DT, Miller RL, Schmidt GA, Pandolfo L (1999) Simulation of recent northern winter climate trends by greenhouse-gas forcing. Nature 399:452–455
Sköld M, Loo L-O, Rosenberg R (1994) Production, dynamics and demography of an Amphiura filiformis population. Mar Ecol Prog Ser 103:81–90
Smirnov AV (1994a) Arctic echinoderms: composition, distribution and history of the fauna. In: David B, Guille A, Féral JP, Roux M (eds) Echinoderms through time (Echinoderms Dijon). Balkema, Rotterdam, pp 135–143
Smirnov IS (1994b) Biogeography and area types of the Southern Ocean ophiuroids (Echinodermata, Ophiuroidea). In: David B, Guille A, Féral JP, Roux M (eds) Echinoderms through time (Echinoderms Dijon). Balkema, Rotterdam, pp 477–488
Smith AB, Paterson GLJ, Lafay B (1995) Ophiuroid phylogeny and higher taxonomy: morphological, molecular and paleontological perspectives. Zool J Linn Soc 114:213–243
Smith Jr KL (1974) Oxygen demands in San Diego Trough sediments: an in-situ study. Limnol Oceanogr 19:939–944
Smith Jr KL, Hinga KR (1983) Sediment community respiration in the deep sea. In: Rowe GT (ed) The sea, vol. 8: Deep-sea biology. Wiley, New York, pp 331–337
Smith Jr WO, Nelson DM (1985) Phytoplankton bloom produced by a receding ice edge in the Ross Sea: spatial coherence with the density field. Science 227:163–167
Smith Jr WO, Sakshaug E (1990) Polar phytoplankton. In: Smith Jr WO (ed) Polar oceanography, Part B: Chemistry, biology, and geology. Academic, San Diego, pp 477–519
Smith SD, Muench RD, Pease CH (1990) Polynyas and leads: an overview of physical processes and environment. J Geophys Res 95:9461–9479
Snelgrove PVR, Butman CN (1994) Animal-sediment relationships revisited: cause versus effect. Oceanogr Mar Biol Annu Rev 32:111–177
Slagstad D (1985) A model of phytoplankton in the marginal ice zone of the Barents Sea. In: Gray JS, Christiansen ME (eds) Marine biology of polar regions and effects of stress on marine organisms. Wiley, Chichester, pp 35–48
Soltwedel T (1999) The expedition ARKTIS XV/1 of RV “Polarstern" in 1999: scientific dives with the ROV system. Ber Polarforsch 339:7–9
Soltwedel T, Juterzenka Kv, Premke K, Klages M (2003) What a lucky shot! Photographic evidence for a medium-sized natural food-fall at the deep seafloor. Oceanol Acta 26:623–628
Soltwedel T, Schewe I (1998) Activity and biomass of the small benthic biota in the Central Arctic Ocean. Polar Biol 19:52–62
Somero GN (1990) Life at low volume change: hydrostatic pressure as selective factor in the aquatic environment. Am Zool 30:123–135
Somero GN, Siebenaller JF, Hochachka PW (1983) Biochemical and physiological adaptations of deep-sea animals. In: Rowe GT (ed) The sea, Vol. 8: Deep-sea biology. Wiley and Sons, New York, pp 261–330
Spielhagen R (2001) Enigmatic Arctic ice sheets. Nature 410: 427–428
Starmans A, Gutt J, Arntz WE (1999) Mega-epibenthic communities in Arctic and Antarctic shelf areas. Mar Biol 135:269–280
Stein R (1996) Organic-carbon and carbonate distribution in Eurasian continental margin and Arctic Ocean deep-sea surface sediments: sources and pathways. Ber Polarforsch 212:243–267
Stein R, Fahl K, Fütterer DK, Galimov EM, Stepanets OV (eds) (2003) Siberian river run-off in the Kara Sea: characterisation, quantification, variability, and environmental significance. Elsevier, Amsterdam, 484 pp
Stein R, Macdonald RW (eds) (2003) The organic carbon cycle in the Arctic Ocean: present and past. Springer, Berlin Heidelberg New York, 363pp
Stewart PL (1983) Measurements of benthic macroinvertebrate standing crop of the Canadian continental shelf and slope of southern Davis Strait and Ungava Bay. Can J Fish Aquat Sci 40:652–658
Stirling I (1980) The biological importance of polynyas in the Canadian Arctic. Arctic 33:303–315
Suess E (1980) Particulate organic carbon flux in the oceans—surface productivity and oxygen utilization. Nature 288:260–265
Thiede J, Timokhov L, Bauch HA, Bolshiyanov DY, Dmitrenko I, Eicken H, Fahl K, Gukov A, Hölemann J, Hubberten H-W, von Juterzenka K, Kassens H, Melles M, Petryashov V, Pivovarov S, Priamikov S, Rachold V, Schmid MK, Siegert C, Spindler M, Stein R, Scientific Party (1999) Dynamics and history of the Laptev Sea and its continental hinterland: a summary. In: Kassens H, Bauch HA, Dmitrenko I, Eicken H, Hubberten H-W, Melles M, Thiede J, Timokhov L (eds) Land-ocean systems in the Siberian Arctic: dynamics and history. Springer, Berlin Heidelberg New York, pp 695–711
Thomsen L, Graf G, Juterzenka Kv, Witte U (1995) An in situ experiment to investigate the depletion of seston above an interface feeder field on the continental slope of the western Barents Sea. Mar Ecol Prog Ser 123:295–300
Thomson MRA, Crame JA, Thomson JW (1991) Geological evolution of Antarctica. Cambridge University Press, Cambridge, 722 pp
Timokhov LA (1994) Regional characteristics of the Laptev and the East Siberian Seas: climate, topography, ice phases, thermohaline regime, circulation. Ber Polarforsch 144:15–31
Tyler PA (1995) Conditions for the existence of life at the deep-sea floor: an update. Oceanogr Mar Biol Annu Rev 33:221–244
Van Dover CL (2000) The ecology of deep-sea hydrothermal vents. Princeton University Press, Princeton, 424 pp
Vanaverbeke J, Arbizu PM, Dahms HU, Schminke HK (1997) The metazoan meiobenthos along a depth gradient in the Arctic Laptev Sea with special attention to nematode communities. Polar Biol 18:391–401
Vanegas SA, Mysak LA (2000) Is there a dominant timescale of natural climate variability in the Arctic? J Climate 13:3412–3434
Vanreusel A, Clough L, Jacobsen K, Ambrose W, Jivaluk J, Ryheul V, Herman R, Vincx M (2000) Meiobenthos of the central Arctic Ocean with special emphasis on the nematode community structure. Deep-Sea Res I 47:1855–1879
Vinogradova NG (1979) The geographical distribution of the abyssal and hadal (ultra-abyssal) fauna in relation to the vertical zonation of the ocean. Sarsia 64:41–50
Walsh JJ, Biscaye PE, Csanady GT (1988) The 1983–1984 shelf edge exchange processes (SEEP)-I experiment: hypothesis and highlights. Cont Shelf Res 8:435–456
Walsh JJ, Dieterle DA, Muller-Karger FE, Aagaard K, Roach AT, Whitledge TE, Stockwell D (1997) CO2 cycling in the coastal ocean. II. Seasonal organic loading to the Canadian Basin from source waters south of Bering Strait. Cont Shelf Res 17:1–36
Wassmann P, Peinert R, Smetacek V (1991) Patterns of production and sedimentation in the boreal and polar northeast Atlantic. Polar Res 10:209–228
Warwick RM, Clarke KR (1995) New ’biodiversity’ measures reveal a decrease in taxonomic distinctness with increasing stress. Mar Ecol Prog Ser 129:301–305
Wheeler PA, Gosselin M, Sherr E, Thibault D, Kirchman DL, Benner R, Whitledge TE (1996) Active cycling of organic carbon in the central Arctic Ocean. Nature 380:697–699
White MG (1984) Marine benthos. In: Laws RM (ed) Antarctic ecology, vol. 2. Academic, London, pp 421–461
Whittaker RH (1972) Evolution and measurement of species diversity. Taxon 21:213–251
Wilson EO (2003) The encyclopedia of life. Trends Ecol Evol 18:77–80
Wlodarska-Kowalczuk M, Pearson TH (2004) Soft-bottom macrobenthic faunal associations and factors affecting species distributions in an Arctic glacial fjord (Kongsfjord, Spitsbergen). Polar Biol 27:155–167
Wlodarska-Kowalczuk M, Weslawski JM (2001) Impact of climate warming on Arctic benthic biodiversity: a case study of two Arctic glacial bays. Clim Res 18:127–132
Wlodarska-Kowalczuk M, Weslawski JM, Kotwicki L (1998) Spitsbergen glacial bays macrobenthos—a comparative study. Polar Biol 20:66–73
Zenkevitch LA (1963) Biology of the seas of the USSR. George Allen and Unwin, London, 955 pp
Acknowledgments
I would like to thank Gotthilf Hempel (Kiel), Karen von Juterzenka (Kiel), Howard Feder (Fairbanks) and two anonymous reviewers for their constructive suggestions on the manuscript. Moreover, I gratefully acknowledge the input of all my colleagues who have helped with the development of the ideas presented in this essay.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Piepenburg, D. Recent research on Arctic benthos: common notions need to be revised. Polar Biol 28, 733–755 (2005). https://doi.org/10.1007/s00300-005-0013-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-005-0013-5