
Abstract
Data on the timing of pupping by the three species of phocid that breed on the Antarctic pack-ice (crabeater, Ross and leopard seals) are limited. Better information would improve our understanding of these species' population and reproductive ecologies, and could facilitate planning and design of population surveys. Observations of the presence or absence of pups with adults during numerous voyages of the Australian National Antarctic Research Expeditions to East Antarctica during spring and early summer months are analysed and presented. The earliest sighting in any year of a crabeater pup accompanied by an adult was on 2 October and the latest sighting on 15 December. The ratio of crabeater pups to adults increased rapidly during the 10-day period 16–25 October, implying a pulse of births over this time. Ross seal pups with an accompanying adult were sighted between 24 October and 22 November, with a peak in the pup-adult ratio occurring in the period 6–15 November. Leopard seal pups were sighted between 8 November and 25 December, with the pup-adult ratio relatively constant during this period. The data provide circumstantial evidence that the maximum durations of lactation reported in the literature for the three species may be over-estimates. If lactation is shorter than reported, asynchrony in the timing of pupping, either among or within years, is implied.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Four species of pinniped (crabeater Lobodon carcinophagus, leopard Hydrurga leptonyx, Ross Ommatophoca rossii and Weddell Leptonychotes weddellii seals) breed entirely on the sea-ice surrounding Antarctica. The first three species (here, collectively called pack-ice species) breed on the floating pack-ice, the last on the fast-ice close to the Antarctic continent. The proximity of many Weddell seal populations to continental stations has facilitated extensive study of this species compared to the three pack-ice species (e.g. Plötz 1986; Testa 1987; Green et al. 1995). In contrast, the remoteness of the pack-ice species' habitat to research expeditions has meant that many aspects of their biology and ecology remain poorly known or unknown.
The three pack-ice species are known to breed in the spring to early-summer months (Laws 1984). However, more precise understanding of the timing of pupping is limited by the paucity of sightings at this time of the year when the extent of pack-ice is intermediate or close to maximal and access by research expeditions is very difficult. More precise quantitative information on the timing of pupping for these species could improve our understanding of their population and reproductive ecologies, and facilitate the planning and design of population surveys (the pupping season is the only time of the year when the entire female breeding population is potentially hauled out on the ice and "available" for surveying). We present here observations of pupping by pack-ice seals made on numerous voyages of the Australian National Antarctic Research Expeditions to East Antarctica during spring and early summer.
Materials and methods
Five research voyages in 1985, 1987 and 1997–1999 were undertaken specifically to carry out ship and aerial surveys, or study the biology and ecology, of crabeater seal populations during the breeding season (e.g. Shaughnessy and Kerry 1989). The ship's track on these voyages was determined by the pack-ice seal research and survey requirements. Shipboard observations were made continuously from the bridges of the M.V. Nella Dan (1985, 1987) and R.S.V. Aurora Australis (1997–1999) during daylight hours while moving through the ice. During research voyages, aerial observations were made from Hughes 500 (1985, 1987) and Sikorsky S76 (1997–1999) helicopters flying at constant altitude and speed (225 and 120 m altitude, and 90 and 80 knots speed, respectively). Ten early-season (September/December) resupply voyages in 1984–1987 (Nella Dan) and 1994–1997 (Aurora Australis) also provided a platform for shipboard observations of pack-ice seals by dedicated observers using the same survey techniques applied on research voyages. In addition, further observations on pack-ice seals were made opportunistically by observers whose primary task was to collect data on seabirds (Woehler 1997) during numerous other voyages between 1980 and 2000. Unlike research voyages, the ship's track on resupply and other voyages was determined by priorities unrelated to pack-ice seal research.
Results
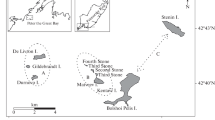
Observations were recorded on research and resupply voyages along a total of 24,900 km of ship track and 19,500 km of aerial track. Ship and aerial tracks on these voyages extended between 60 and 120°E, and focussed on the Prydz Bay region, in September, October and November, but were relatively evenly distributed between longitudes 75 and 150°E in December (Fig. 1). Search effort was biased toward the outer pack-ice in September and October but sampled the north-south extent of ice more thoroughly in November and December. Only sightings of seal groups for which species identity was recorded as certain (87% of all sighted groups) were considered in our analyses. We defined a group according to spatial (individuals <10 m apart) and behavioural criteria. Solitary seals were considered to be a group of size one. These sightings totalled 6,562 groups of crabeater seals, comprising 8,418 adults and 519 pups, 191 groups of Ross seals, comprising 197 adults and 30 pups, and 88 groups of leopard seals, comprising 89 adults and 16 pups.

Distribution of shipboard and aerial search effort by month in East Antarctica during five research and ten resupply voyages when dedicated seal observers were operating. The ice extent for the middle of each month, averaged across years when voyages were conducted (1984–1987 and 1994–1999), is shown as the dashed line
The earliest sighting within any year of a crabeater pup accompanied by an adult was on 2 October (in 1994) and the latest sighting on 15 December (in 1999). An additional 20 pups were sighted unaccompanied by an adult; the first of these was sighted on 25 October (1996) and the last on 13 December (1985). Taken across all years, the ratio of pups to adults increased rapidly during the 10-day period 16–25 October, implying a pulse of births over this time, and the ratio peaked at around 0.24 in the period 21–31 October (Fig. 2). At this time, pairs of adults were more frequent than lone adults (59% and 37% of groups, respectively).

Ratio of pups to adults for crabeater, Ross and leopard seals within 5-day bins from early September to early January in East Antarctica across all years when data were collected. The ratios are calculated for pups accompanied by adults and hence do not include lone pups. Sample size, as number of adults sighted, is shown along the top of each plot
Ross seal pups with an accompanying adult were sighted between 24 October (1987) and 22 November (1985). No pups were sighted alone. Adult Ross seals were almost always solitary (97% of groups). Taken across all years, a peak in the pup-adult ratio of around 0.64 occurred during the 10-day period 6–15 November.
Leopard seal pups were sighted between 8 November (1987) and 25 December (1999), and the pup-adult ratio was relatively constant at around 0.3 between the date of first and last pup sightings. The only leopard seal pup unaccompanied by an adult was sighted on 25 December (1999). Like Ross seals, adult leopard seals were almost always solitary (98% of groups).
The higher peak in pup-adult ratios for Ross and leopard than crabeater seals (Fig. 2) are likely to reflect inter-specific differences in haul-out and grouping behaviours by adults rather than differing reproductive rates.
Discussion
The aspects of reproduction that would contribute to the observed temporal patterns in pup-adult ratios, namely mean birth date, mean lactation period, and the variability about these means, are poorly known in pack-ice seals, as opportunities for directly observing birth and weaning of individual animals are extremely limited. Current knowledge of these parameters is largely based on indirect estimation or speculation.
To our knowledge, there are no published observations of crabeater seal births in the wild. Øritsland (1970) observed newly born pups on 2 and 29 October (1964), and found no pregnancies in shot samples of females after 11 October (1964) near the South Orkney Islands. Laws (1984) reported a mean birth date of 19 October but did not comment on variability. Siniff et al. (1979) reported sighting pups as early as 7 October, with a peak from 23 to 26 October (1975–1977) in the Bransfield and Gerlache Straits region. Shaughnessy and Kerry (1989) observed pups on sea-ice off Enderby Land in East Antarctica on 29 October and 3 November (1985) with remains of umbilical cords that they considered were <11 days old. Our data suggest that variation in the birth date in East Antarctica extends from early October until at least late October when the pulse in the pup-adult ratio occurred, which is largely in keeping with the above reports.
Laws (1958) estimated the duration of lactation in crabeater seals to be up to 6 weeks, from the pattern of growth layers in teeth. Siniff et al. (1979) speculated on a shorter period of 4 weeks, but acknowledged that final determination required studies of marked animals. Shaughnessy and Kerry's (1989) estimated lactation period of 14–21 days (mean 17 days) is based on consideration of the birth mass, measured weaning mass, and measured daily mass gain. Assuming that pups accompanied by an adult are still suckling and that lone pups have weaned, the difference in time between the first sighting of a pup with an adult and the first sighting of a lone pup should approximate the duration of the lactation period. From our data, this difference in the time of sightings (2 and 25 October: 23 days) is near the upper end of the range estimated by Shaughnessy and Kerry (1989), supporting their contention of a shorter lactation period than previously suggested by Laws (1958) and Siniff et al. (1979). A shorter lactation period of ≤ 3 weeks is also more consistent with that known for northern-hemisphere seals breeding in a similar habitat (King 1983; Reidman 1990). Given that crabeater pups were sighted over a period of >2 months, accepting a lactation period of ≤ 3 weeks implies variation of up to 5 weeks in the timing of the onset of pupping.
Similarly, we are unaware of any published observations of Ross seal births, and few sightings of pups, in the wild. Solyanik (1964) reported a female and pup on 6 December (1950) at the South Sandwich Islands, but no assessment was provided of its age. Thomas et al. (1980) found a newborn pup on 14 November (1978) in the Antarctic Peninsula region. Tikhomirov (1975) reported eight pups that were estimated to have been born between 3 and 18 November (1968) near the Balleny Islands. Øritsland (1970) found a high pregnancy rate (88%) in eight females collected in September/October (1964) near the South Orkney Islands. King (1983) noted, from examination of reproductive tracts, that mating possibly occurred at the end of December, and by extrapolation that pups could be born in mid-November. With the exception of Solyanik (1964), all these reports are consistent with our observations.
Lactation in Ross seals has been variously reported to last 4 weeks (Reidman 1990; Bonner 1994) and 4–6 weeks (Skinner and Klages 1994), although the basis for these estimates is unclear. Our sighting of adults with pups over the 4-week period from 24 October to 22 November (in all years), assuming this represents the full period over which pups are present with adults, is inconsistent with the upper limit for the lactation period of 6 weeks, and if the lower limit of 4 weeks were accepted, very tight synchrony in births of just a few days would be implied. In the absence of any further data, we question whether the lactation period for Ross seals may, in reality, be shorter than the very speculative estimates currently in the literature.
The only reported births of a leopard seal in the wild are at Heard Island on 14–15 November (1951) (Brown 1957) and at Bird Island in September (1987) (Walker et al. 1998), both well north of the pack ice where most breeding occurs. Hamilton (1939) noted sightings of new-born pups by other investigators in September (no location or year given) and November (South Orkney Islands, no year given). Siniff et al. (1978) reported sighting two females with newly born pups on 7 and 13 November (1977) in the Antarctic Peninsula region. In a sample of nine females collected between 6 and 22 November (1979) in the same region, Siniff et al. (1980) found one to be pregnant and another to have given birth. Siniff and Stone (1985) inferred birth of pups from October to mid-November from observation of near-term foetuses and nursing pups. These reports, apart from Hamilton's (1939) sighting of pups in September, are consistent with our observations for East Antarctica.
The understanding of the duration of lactation in the leopard seal is also very poor. Based on the advanced development of teeth in a new-born pup on Heard Island, Brown (1957) proposed that it may be as short as 10–14 days, which is consistent with the lactation periods of northern-hemisphere seals breeding in similar habitats (King 1983; Reidman 1990). Maxwell (1967), in contrast, proposed a period of about 8 weeks, but the basis of this estimate is unclear. More recent opinion, though still lacking clear evidence and qualified as speculative by its proponents, is a shorter period of 4 weeks (King 1983; Laws 1984; Reidman 1990). Our observation of adults with pups over a 7-week period from 8 November to 25 December, assuming this represents the full period over which pups are present with adults, is inconsistent with the upper limit for the lactation period of 8 weeks. If the alternative estimates of 2 or 4 weeks are accepted, asynchrony in births of 3–5 weeks is implied. This is consistent with the observation by King (1983) of a large variation in the length of foetuses sampled at one time.
Our data provide circumstantial evidence that the maximum durations of lactation reported in the literature for the 3 species may be over-estimates. Shorter lactation periods would seem advantageous, given the unstable nature of the breeding platform, and would be consistent with better-studied species occupying similar habitats in the northern hemisphere (e.g. the harp seal, Pagophilus groenlandicus; Stewart and Lavigne 1980). If lactation periods are indeed relatively short, asynchrony in the time of pupping, either among years or within years, is implied. Within-year asynchrony may occur along some environmental gradient, as demonstrated for harbour seal (Phoca vitulina) by Temte et al. (1991), or be constant across the entire region. A detailed analysis of synchrony among years or along environmental gradients is beyond the scope of this paper, but further such analysis, at least for crabeater seals for which sample sizes are large, could provide insights into the little-known breeding strategies of pack-ice seals.
References
Bonner N (1994) Seals and sea lions of the World. Blandford, London
Brown KG (1957) The leopard seal at Heard Island, 1951–54. ANARE Int Rep 16
Green K, Burton HR, Watts DJ (1995) Studies of the Weddell seal in the Vestfold Hills, East Antarctica. ANARE Res Note 93
Hamilton JE (1939) The leopard seal Hydrurga leptonyx (de Blainville). Discovery Rep 18:239–264
King JE (1983) Seals of the world. University of Queensland Press, St Lucia
Laws RM (1958) Growth rates and ages of crabeater seals, Lobodon carcinophagus Jacquinot and Pucheran. Proc Zool Soc Lond 130:275–288
Laws RM (1984) Seals. In: Laws RM (ed) Antarctic ecology. Academic, London, pp 621–715
Maxwell G (1967) Seals of the world. Constable, London
Øritsland T (1970) Sealing and seal research in the south west Atlantic pack ice, Sept-Oct 1964. In: Holdgate MW (ed) Antarctic ecology. Academic, London, pp 367–376
Plötz J (1986) Summer diet of Weddell seals (Leptonychotes weddelli) in the eastern and southern Weddell Sea, Antarctica. Polar Biol 6:97–102
Reidman M (1990) Seals, sea lions and walruses. University of California Press, Los Angeles
Shaughnessy PD, Kerry KR (1989) Crabeater seals Lobodon carcinophagus during the breeding season: observations on five groups near Enderby Land, Antarctica. Mar Mammal Sci 5:68–77
Siniff DB, Stone S (1985) The role of the leopard seal in the tropho-dynamics of the Antarctic marine ecosystem. In: Siegfried WR, Condy PR, Laws RM (eds) Antarctic nutrient cycles and food webs. Springer, Berlin Heidelberg New York, pp 555–559
Siniff DB, Laws R, Øritsland T, Stirling I, Bengtson J, DeMaster D, Reichle R (1978) Biota of the antarctic pack ice: R/V Hero cruise 77-5. Antarct J US 13:161–162
Siniff DB, Stirling I, Bengtson JL, Reichle RA (1979) Social and reproductive behavior of crabeater seals (Lobodon carcinophagus) during the austral spring. Can J Zool 57:2243–2255
Siniff DB, Stone S, Reichle D, Smith T (1980) Aspects of leopard seals (Hydrurga leptonyx) in the Antarctic pack ice. Antarct J US 15:160
Skinner JD, Klages NTW (1994) On some aspects of the biology of the Ross seal Ommatophoca rossii from King Haakon VII Sea, Antarctica. Polar Biol 14:467–472
Solyanik GA (1964) Experiment in marking seals from small ships. Sov Antarct Exped Bull 5:212
Stewart REA, Lavigne DM (1980) Neonatal growth in Northwest Atlantic harp seals, Pagophilus groenlandicus. J Mammal 61:670–680
Temte JL, Bigg MA, Wiig O (1991) Clines revisited: the timing of pupping in the harbour seal (Phoca vitulina). J Zool Lond 224:617–632
Testa JW (1987) Long-term reproductive patterns and sighting bias in Weddell seals (Leptonychotes weddelli). Can J Zool 65:1091–1099
Thomas J, DeMaster D, Stone S, Andriashek D (1980) Observations of a newborn Ross seal pup (Ommatophoca rossii) near the Antarctic peninsula. Can J Zool 58:2156–2158
Tikhomirov EA (1975) Biology of the ice forms of seals in the Pacific sector of the Antarctic. Rapp PV Reun Cons Int Explor Mer 169:409–412
Walker TR, Boyd IL, McCafferty DJ, Huin N, Taylor RI, Reid K (1998) Seasonal occurrence and diet of leopard seals (Hydrurga leptonyx) at Bird Island, South Georgia. Antarct Sci 10:75–81
Woehler EJ (1997) Seabird abundance, biomass and prey consumption within Prydz Bay, Antarctica, 1980/81–1992/93. Polar Biol 17:371–383
Acknowledgements
We thank the captains and crews of the M.V. Nella Dan and R.S.V. Aurora Australis on the numerous voyages when this work was undertaken, and the many expeditioners who assisted in observations. Melissa Sharpe extracted data on seal sightings obtained by seabird observers.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Southwell, C., Kerry, K., Ensor, P. et al. The timing of pupping by pack-ice seals in East Antarctica. Polar Biol 26, 648–652 (2003). https://doi.org/10.1007/s00300-003-0534-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-003-0534-8