
Abstract
Key message
VvMATE1 and VvMATE2 encode putative PA transporters expressed during seed development in grapevine. The subcellular localization of these MATE proteins suggests different routes for the intracellular transport of PAs.
Abstract
Proanthocyanidins (PAs), also called condensed tannins, protect plants against herbivores and are important quality components of many fruits. PAs biosynthesis is part of the flavonoid pathway that also produces anthocyanins and flavonols. In grape fruits, PAs are present in seeds and skin tissues. PAs are synthesized in the cytoplasm and accumulated into the vacuole and apoplast; however, little is known about the mechanisms involved in the transport of these compounds to such cellular compartments. A gene encoding a Multidrug And Toxic compound Extrusion (MATE) family protein suggested to transport anthocyanins—named VvMATE1—was used to identify a second gene of the MATE family, VvMATE2. Analysis of their deduced amino acid sequences and the phylogenetic relationship with other MATE-like proteins indicated that VvMATE1 and VvMATE2 encode putative PA transporters. Subcellular localization assays in Arabidopsis protoplasts transformed with VvMATE–GFP fusion constructs along with organelle-specific markers revealed that VvMATE1 is localized in the tonoplast whereas VvMATE2 is localized in the Golgi complex. Major expression of both genes occurs during the early stages of seed development concomitant with the accumulation of PAs. Both genes are poorly expressed in the skin of berries while VvMATE2 is also expressed in leaves. The presence of putative cis-acting elements in the promoters of VvMATE1 and VvMATE2 may explain the differential transcriptional regulation of these genes in grapevine. Altogether, these results suggest that these MATE proteins could mediate the transport and accumulation of PAs in grapevine through different routes and cellular compartments.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Proanthocyanidins (PAs)—also known as condensed tannins—are abundant flavonoid compounds that play important roles in the interaction between plants and their environment. They can protect plants against diseases and herbivores, and are involved in physiological functions such as seed dormancy and viability (Dixon et al. 2005; Lepiniec et al. 2006). PAs are polyphenolic polymers consisting of flavan 3-ol subunits, (+)-catechin and/or (−)-epicatechin which are synthesized via the flavonoid pathway that also produces anthocyanins and flavonols (Boss et al. 1996; Xie et al. 2003). In grapevine (Vitis vinifera L.), tannins are of great importance to red wine quality and are associated to organoleptic properties such as astringency and bitterness (Peleg et al. 1999; Gawel et al. 2001). These compounds play important roles during storage by the interaction with other flavonoids involved in long-term color stability (Remy et al. 2000; Escribano-Bailon et al. 2001; Wang et al. 2003). In grape fruits, PAs are present in seed and skin tissues (Kennedy et al. 2001; Kennedy and Jones 2001) and in lesser extent in the pulp (Mané et al. 2007; Verries et al. 2008). These flavonoids are actively synthesized during flower and fruit development, mainly at the green stage of berry development. In fruit skin, PAs mainly accumulate before veraison (defined as the onset of ripening in grapevine) and when anthocyanins start to be synthesized (Kennedy et al. 2001; Downey et al. 2003; Verries et al. 2008). In contrast, PA accumulation in seeds appears to continue for 1–2 weeks after veraison, suggesting that PA accumulation in seeds is independent of that in the skin (Downey et al. 2003; Bogs et al. 2005).
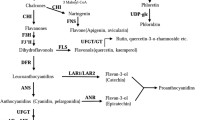
At the cellular level, synthesis of PA precursors and other flavonoids is believed to take place at the cytoplasmic side of the endoplasmic reticulum (ER) by an enzymatic complex and then transported for storage (Winkel 2004). Several studies have shown that PAs are stored in the vacuoles of seed coat endothelial cells during Arabidopsis early seed development (Abrahams et al. 2003; Kitamura et al. 2004) and it is likely (but not definitively proven) that PA oligomerization/polymerization occurs in the vacuole (Pourcel et al. 2005). However, the major fractions of PAs in grapes are accumulated in the apoplast in both seeds and skin, and tannins from cell walls have a higher degree of polymerization than those from the inner part of the cell (Gény et al. 2003). Because flavonoids are normally accumulated in cellular compartments other than those where they are synthesized, their transport to the final destination is a critical process in PA biosynthesis and assembly. Even in Arabidopsis, transport of flavonoids from the ER to the vacuole is not completely understood. Currently, two proposed models for this process include vesicle trafficking and membrane transporters (Grotewold and Davies 2008; Braidot et al. 2008; Zhao and Dixon 2009; Zhao et al. 2010). In Arabidopsis seedlings, Poustka et al. (2007) observed that anthocyanins accumulate in vesicle-like structures probably derived from the ER. These structures have also been observed in grapevine hairy roots and in cell suspension cultures (Cutanda-Pérez et al. 2009; Conn et al. 2010; Gomez et al. 2011). Additionally, new evidence suggests that tannin synthesis could also take place in chloroplasts, their polymerization inside tannosomes, chloroplast-derived organelles, and their final transport to the vacuole in structures named tannosome shuttle (Wang et al. 2010; Brillouet et al. 2013). Regarding the membrane transport mechanism, two families of transporters have been shown to participate in anthocyanin and PA transport: the ATP-binding cassette (ABC) transporter family and Multidrug And Toxic Compound Extrusion (MATE) transporters. A mutation in the maize ABC-C transporter ZmMRP3 caused a reduction in anthocyanin production and pigment mislocalization (Goodman et al. 2004). More recently, the grapevine ABCC1 protein, a tonoplast-localized ABC transporter, was shown to be involved in the transport of anthocyanidin 3-O-glucosides into the vacuole (Francisco et al. 2013). On the other hand, plant MATE proteins constitute a large family that has been involved in the transport of different organic molecules such as salicylic acid (Nawrath et al. 2002), alkaloid vacuolar transport (Shoji et al. 2009) and citrate efflux (Durrett et al. 2007; Liu et al. 2009); (Magalhaes et al. 2007); (Furukawa et al. 2007); (Yokosho et al. 2009); (Maron et al. 2010). In addition, MATE transporters have been implicated in the transport of flavonoids (Mathews et al. 2003); (Zhao and Dixon 2009; Zhao et al. 2011); (Gomez et al., 2009); (Frank et al. 2011). In Arabidopsis, three genes have been reported to participate in the vacuolar transport of flavan-3-ol (epicatechin). Mutations in TT12 or TT19 affect PA accumulation producing pale seed coats with a transparent testa phenotype due to a reduction in oxidized PAs (Debeaujon et al. 2003; Kitamura et al. 2004). TT12 exhibits a strong homology to MATE transporters (Debeaujon et al. 2001; Marinova et al. 2007) whereas TT19 to members of the multigenic glutathione S-transferase (GST) family, essential for anthocyanin and PA accumulation in Arabidopsis, maize and petunia (Kitamura et al. 2004, 2010). The third gene, AHA10, encodes a H+-ATPase (H+ pump) that is indirectly involved in PA metabolism by producing a H+ gradient necessary to support the MATE’s H+/flavonoid antiporter function (Baxter et al. 2005; Zhao et al. 2010). In grapevine, VvGST4 is strongly associated to anthocyanin accumulation and is able to complement the maize mutant bz2, defective in anthocyanin transport (Conn et al. 2008). Down-regulation of VvGST4 expression in grapevine hairy roots showed that this protein is necessary for anthocyanin transport from the ER to the tonoplast independently of flavonoid-filled vesicle trafficking (Gomez et al. 2011).
Two grapevine MATE transporters, named anthoMATEs (AMs), were shown to be involved in the transport of acylated anthocyanins in vitro. AM1 and AM3 are proteins mainly targeted to the tonoplast and membrane structures attached to the nucleus (Gomez et al. 2009). However, in tissues that accumulate anthocyanins, these proteins are closely associated to small anthocyanin-filled vesicles derived from the ER (rather than the tonoplast). AMs are also necessary for the formation of the ER-derived vesicles (Gomez et al. 2011). Grapevine genes homologous to AHA10 have yet to be reported. Despite all these reports, little is known about the PA transport process in grapevine. Most proteins involved in flavonoid transport have been characterized only for anthocyanin transport. Ectopic expression of VvMYBPA1 or VvMYBPA2 in grapevine hairy roots promotes PA synthesis and induction of a set of PA biosynthetic genes. Interestingly, one up-regulated gene encodes a putative MATE transporter (Terrier et al. 2009). Because this gene is highly homologous to TT12, functional characterization of homologous genes in the grape genome could provide better understanding on the mechanisms of PA transport and accumulation in this species. In line with this notion, we isolated and characterized two genes, VvMATE1 and VvMATE2. Sequence analysis, subcellular localization and expression pattern during berry development of these grapevine MATE members suggest that both genes encode putative PA transporters with probable involvement in different transport pathways and accumulation of PAs in Vitis vinifera L.
Results
VvMATE1 and VvMATE2 are members of the MATE family of transporters
To identify genes encoding putative MATE proteins in the genome of Vitis vinifera, a search was conducted in the Genoscope database (http://www.genoscope.cns.fr) for those presenting the highest sequence similarity to GSVIVT18839001 (named VvMATE1), which was described as highly homologous to TT12 from Arabidopsis thaliana (Terrier et al. 2009). Only the sequence GSVIVT00018841001 (named VvMATE2) had an identity greater than 95 %. The full-length coding sequences of VvMATE1 and VvMATE2 were amplified and cloned from cDNA of Carménère berries in the veraison stage. Their sizes were 1,521 and 1,515 bp and were predicted to encode a protein of 506 and 504 amino acid residues, respectively.
To establish the phylogenetic relationship between VvMATE1 and VvMATE2 with plant MATE-like proteins, a phylogenetic tree was created using the deduced full-length amino acid sequences of VvMATE1 and VvMATE2. The analysis was performed with proteins that were described to be involved in PA transport such as Arabidopsis TT12 (Debeaujon et al. 2001), Malus x domestica MATE1 and MATE2 (Frank et al. 2011) and Medicago truncatula MATE1 (Zhao and Dixon 2009); and in anthocyanin transport, AM1 and AM3 from grapevine (Gomez et al. 2009) and MTP77 from tomato (Mathews et al. 2003). Proteins encoded by these grapevine genes specifically clustered with MATE proteins involved in PA transport like those of M. domestica, M. truncatula, Arabidopsis and Brassica rapa (Fig. 1) and were clearly separated from other MATE transporters participating in anthocyanin, nicotine or citrate compartmentation. Others putative transporters from Vitis vinifera grouped into a different clade showing high homology with NtMATE1 and NtMATE2 have been described as nicotine transport into the vacuoles of tobacco roots (Shoji et al. 2009) (Fig. 1).

Phylogenetic relationship of VvMATE1 and VvMATE2 with other plant MATE transporters. The tree was obtained from the multiple alignments of the deduced VvMATE and other MATE proteins following the neighbor-joining method in the MEGA4 package with 1,000 replicates for bootstrap values
VvMATE1 and VvMATE2 share 84 % amino acid sequence identity with each other and present significant similarities with other MATE proteins as determined by a BLASTP search. The most similar sequences to both grapevine proteins were Malus x domestica MATE1 (78 and 82 % identity, respectively) and MATE2 proteins (76 and 82 % identity, respectively) and with Populus tricocarpa XP-002307572 with 77 and 81 % identity, respectively. VvMATE1 and VvMATE2 are the closest sequences to the Arabidopsis TT12 of all MATE-like proteins found in grapevine (with a 72 and 74 % identity, respectively). Multiple alignments of VvMATE1 and VvMATE2 with MATE-like proteins revealed a strong sequence homology among them (Fig. 2). Pfam database predicted two MatE domains (PF01554) in both grapevine MATE proteins, that are similar to the MatE domain present in TT12 and other PA-transporting MATE proteins. In agreement with the structure of other MATE family members involved in PAs transport, analysis of VvMATE1 and VvMATE2 with the HMMTOP program predicted 12 transmembrane helices consistent with those observed in other MATE transporters (Debeaujon et al. 2001).

Alignment of the deduced amino acid sequences of VvMATE and VvMATE2 and PA-transporting MATE transporters by ClustalW. Grapevine VvMATE1 (CAO69962) and VvMATE2 (CAO69963), Arabidopsis TT12 (NP_191462), M. truncatula MtMATE1 (FJ858726) and apple MdMATE1 (GU064953). The predicted MATE domains (PF01554) using the Pfam database are shown in dotted line. Identical amino acid residues among predicted protein sequences are shaded in black. Black bars indicate the 12 transmembrane domains (TM) predicted by the TMHMM program (http://www.cbs.dtu.dk/services/TMHMM/)
At the genomic level, both genes are located in the same scaffold in chromosome 12 (Fig. 3). Their structure reveals high conservation in the size and number of introns/exons (both containing eight exons interrupted by seven introns), similar to the structure observed in other genes that encode PA-transporting MATE proteins (Debeaujon et al. 2001; Frank et al. 2011).

Genomic structures of VvMATE1 and VvMATE2. Both genes were present at the scaffold_8 at the chromosome 12 on the genoscope database (http://www.genoscope.cns.fr)
VvMATE1 is targeted to the tonoplast while VvMATE2 is localized to the Golgi complex
Due to the strong evidence that MATE proteins involved in PA transport are confined in the tonoplast, the subcellular localization of the grapevine proteins was investigated. First, a subcellular localization prediction was performed by in silico analysis of their deduced amino acid sequence. Wolf PSORT-II software (Horton et al. 2007) predicts a vacuolar location for VvMATE1, coincident with other PA transporters (Debeaujon et al. 2001; Zhao and Dixon 2009) whereas in VvMATE2 it showed a possible destination to the plasma membrane or the Golgi complex. To establish this, each cDNA was fused to GFP under the control of the CaMV 35S promoter. In addition, the vacuolar marker V-rk fused to the mCherry reporter (Nelson et al. 2007) with a red signal was used in the co-transformation of Arabidopsis protoplasts. Fluorescence from GFP and mCherry was observed after 24 h of incubation post transfection (Fig. 4). In the case of VvMATE1, the GFP signal overlapped with the mCherry signal from the vacuolar marker, observed in the merged images (Fig. 4c, d), strongly suggesting that this protein is localized to the tonoplast. On the other hand, co-localization assays of VvMATE2 with V-rk showed that VvMATE2–GFP did not localize to vacuoles (Supplementary Fig. 1). On the contrary, VvMATE2 was visualized as small dotty structures (with a granular pattern) similar to Golgi structures. To confirm this, other organelle-specific markers were used. When an ER marker fused to mCherry and MATE2-GFP were co-expressed in protoplasts (Fig. 4i, j), fluorescence from GFP and mCherry did not completely overlap, but apparently VvMATE2 embedded the ER (Fig. 4k, l). However, after transforming the protoplasts with a Golgi marker fused to mCherry, the GFP signal was found to overlap with the mCherry signal (Fig. 4g, h), indicating that the VvMATE2–GFP fusion protein is targeted to the Golgi complex.

Subcellular localization of VvMATE1 and VvMATE2 proteins. Fluorescent microscopy images presenting subcellular localization of VvMATE1 and VvMATE2 fusion proteins and organelle markers in transiently transformed Arabidopsis protoplasts. (a–d) Co-expression of MATE1-GFP with a vacuole marker (mCherry). (e, f) Co-expression of MATE2-GFP with the Golgi marker (mCherry). (i–l) Co-expression of MATE2-GFP with the ER marker (mCherry). (D, H and L) Merged fluorescence of GFP, mCherry plus chlorophyll (light blue) autofluorescence. Yellow denotes co-localization. Bars in all pictures are 15 μm
VvMATE1 and VvMATE2 are mainly expressed in seed berries
The transcriptional profiles of these putative transporter genes were determined in skin and seeds at different berry developmental stages in Carménère and associated with flavonoid synthesis. Early stages of fruit development and vegetative tissues such as leaves and roots were also analyzed. Spatial and temporal variations in expression were observed for each gene. The transcript levels of VvMATE1 were higher in seeds than in the rest of the tissues analyzed. This gene showed to be highly expressed from the time of the earliest stages of seed development; however, the highest level of transcript was observed at veraison and then decreased during ripening. Significant expression was also detected during flower development and fruit setting. In contrast, transcript levels of VvMATE1 were less abundant in grape skin, similar to those found in vegetative organs (Fig. 5a). On the other hand, VvMATE2 showed a different expression pattern than VvMATE1. This gene was predominantly expressed in grape seeds and leaves. The expression in seeds presented a peak at fruit setting stage following a decline at pre-veraison and then a second peak at veraison to finally decrease during seed maturity. In leaves, VvMATE2 was found to be transcribed at high levels, being comparable with those found in seeds. Similar to VvMATE1, abundant transcript levels were also found in reproductive tissue and small berries but not in skin and roots (Fig. 5b).

Expression analysis of VvMATE1 (a) and VvMATE2 (b) in vegetative tissues and reproductive tissues during berry development. Seeds (black bars) and skins (gray bars) from pre-veraison were analyzed separately and only whole berries (white bar) and seeds at the fruit set stage were analyzed. Gene expression of VvMATE1 and VvMATE2 is relative to VvGAPDH and data are means of three biological replicates with error bars indicating SD
To understand the molecular basis that may explain the differential expression of both genes, an in silico analysis of a 1,500-bp upstream regulatory region of VvMATE1, VvMATE2 and AtTT12 was performed. Five groups of common target motifs for plant MYB transcription factors, including MYB1AT (WAACCA), MYBCORE (CNGTTR), MYBPLANT (CCWACC), MYBPZM and MYBST1 (GGATA), were identified in the promoters of the three MATE genes. Additionally, the MYB4 binding motif (AACTACC), which is a target of the transcriptional repressor AtMYB4, was also identified. These elements have been related to dehydration response and to regulation of phenylpropanoid biosynthetic genes. The analysis also revealed differences in the type, number and position of the MYB target elements among the promoters. VvMATE1 only presents the MYB1AT motif at positions −603, −1,086 and −1,317 from the translation start codon. Conversely, VvMATE2 contains 1–3 copies of all five above-mentioned MYB target sites, whereas TT12 presents four of them (Fig. 6). The MYB4 binding site was more abundant in the promoter of VvMATE2 (four copies) than in TT12 (3 copies) and VvMATE1 (two copies).

In silico analysis of the promoter region of VvMATE1, VvMATE2 and AtTT12. MYB- and MYC-binding cis-motifs were detected in the sense coding strand of a 1,500-bp upstream region from the start codon using the PLACE and PLANTCARE databases
Furthermore, the analysis led to the identification of organ/tissue specific elements. For example, various cis-elements found in promoters of embryo- and seed-specific genes were present in the TT12, VvMATE1 and VvMATE2 promoters such as the GCN4 core, ACGT, and AACA, which are conserved in seed storage protein genes, mainly in monocot species (Müller and Knudsen 1993; Wu et al. 2000). However, the DPBF motif (ACACNNG) which is mainly embryo specific (Kim et al. 1997) was only present in the promoters of VvMATE1 (−18, −432, −534) and TT12 (−379) (Fig. 6). Specific elements involved in hormonal responsiveness were also detected. The Arabidopsis TT12 promoter presents both ABRE and CGTCA elements, involved in ABA and MeJA responsiveness, respectively. In contrast, the promoter of VvMATE1 contains only an ABRE element (at the position −872), whereas VvMATE2 only has the cis-regulatory element involved in the MeJA responsiveness at positions −28 and −463 from the ATG codon (Fig. 6).
Discussion
Two models have been proposed for the synthesis, transport, polymerization and accumulation of PAs in plants. In the first, the classic model: synthesis of PAs precursors is performed on the cytoplasmic face of rough endoplasmic reticulum by a multienzymatic complex (Winkel 2004), and then transported to different cellular compartments such as the vacuole, the nucleus or the apoplast, either by ER-derived vesicles or by specific membrane transporters (Debeaujon et al. 2003; Kitamura et al. 2004; Saslowsky et al. 2005; Zhao and Dixon 2009; Zhao et al. 2010). A second model, recently described, proposes that in chlorophyllous organs of Tracheophyta, including Vitis vinifera, the synthesis of PAs precursors occurs in chloroplasts, and their polymerization takes place inside chloroplast-derived organelles (tannosomes). Several tannosomes are transported towards the vacuole by structures called tannosome shuttle and then incorporated by invagination of the tonoplast (Wang et al. 2010; Brillouet et al. 2013). To gain insight on the PAs transport and accumulation in grapevine fruits, the characterization of two putative PAs transporter encoding genes identified in the grapevine genome was performed.
VvMATE1 and VvMATE2 encode putative PA-transporting MATE proteins
In the Vitis vinifera genome, 65 virtual genes that encode MATE-like proteins have been reported, however, only few of them have been functionally characterized. The anthoMATE proteins AM1 and AM3 have been associated to acylated anthocyanin transport into the vacuole (Gomez et al. 2009, 2011. As far as we know, no MATE proteins have been characterized as PA or flavonoid transporters in grapevine. Interestingly, Terrier et al. (2009) reported the induction of GSVIVT00018839001 (named VvMATE1 in our study) after the ectopic expression of the transcription factor VvMYBPA1 in grapevine hairy roots. In this work, we described features that point to establish the identity of VvMATE1 and a related protein, VvMATE2, as PA-transporting proteins in Vitis vinifera. First, VvMATE1 and VvMATE2 are the most closely related grapevine proteins to Arabidopsis TT12 and other known PA transporters (Fig. 2; Debeaujon et al. 2001; Zhao and Dixon 2009; Gomez et al. 2009; Frank et al. 2011). Second, the phylogenetic analysis distinguished these VvMATEs from the clade containing anthocyanin transporters (Fig. 1). Third, VvMATE1 and VvMATE2 present the two characteristic MatE domains and share similar membrane topology with PA-transporting MATE proteins (Debeaujon et al. 2001; Frank et al. 2011).
Distinct subcellular localization of VvMATE1 and VvMATE2 may imply different roles in PA accumulation
Following flavonoid synthesis in the cytoplasm, the products are stored mainly in the vacuole and in the cell wall (Kitamura 2006). Most of plant MATE proteins characterized so far appear to be targeted to the vacuolar or plasma membranes (Uhde-Stone et al. 2005; Marinova et al. 2007; Gomez et al. 2009; Zhao and Dixon 2009). The in silico prediction of the tonoplast localization of VvMATE1 was confirmed when the VvMATE1–GFP fusion protein co-localized with a tonoplast marker, strongly suggesting a role as a vacuolar transporter (Fig. 4). This result is highly consistent with the vacuole being an organelle that plays an important role in flavonoid sequestration and storage (Debeaujon et al. 2001). It was interesting to observe that the VvMATE1–GFP signal was only found in the vacuolar membrane and not in ER-derived vesicles as suggested for MATE1 of M. trunculata in tobacco leaf epidermal cells (Zhao and Dixon 2009). Flavonoids/PAs contained in cytoplasmatic prevacuole-like vesicles can end in the vacuole after vesicular fusion (Abrahams et al. 2002). This type of ER-derived anthocyanin-filled vesicles has also been found in grapevine hairy roots cells (Gomez et al. 2011). Subcellular localization experiments showed that grapevine AM1 and AM3 are closely associated to ER-derived vesicle structures (Gomez et al. 2011). Therefore, studying the transport activity, transport mechanism and substrate specificity of VvMATE1 and also determining its precise localization in cells that accumulate PAs in grapevine may contribute to elucidate this controversy. In contrast, localization for VvMATE2 was predicted for the plasma membrane although the in vivo assay shows a Golgi-associated VvMATE2–GFP protein fusion (Fig. 4). While this organelle localization differs from that previously observed for all other MATE proteins transporting flavonoids, is coincident with a Golgi-localized MATE transporter that has been described in Arabidopsis (BCD1) and is involved in organ initiation and iron homeostasis under osmotic stress (Burko et al. 2011; Seo et al. 2012).
In grape fruit, unlike anthocyanins, PAs are accumulated not only in vacuoles but also in the cell wall where they are linked to polysaccharides in both seeds and skin (Gény et al. 2003). The mechanisms involved in PA transport from vacuole and/or cytoplasm to the apoplastic space are still unknown. In this regard, the localization of VvMATE2 in the Golgi complex provides a possible exocytic route for transporting PAs to the cell wall. The Golgi complex can be an intermediary organelle for the final plasma membrane destination of the transporter. Taking these observations into account, we hypothesize that VvMATE2 could be involved in the transport of PAs/flavonoids to the apoplast. The absence of plasma membrane-associated VvMATE2 in our approach could be explained by the use of protoplasts derived from Arabidopsis leaves, since cells from this tissue do not accumulate PAs in their cell wall. Therefore, its localization in cells that accumulate flavonoids in the apoplast deserves further investigation.
VvMATE1 and VvMATE2 have different expression patterns
In grapevine, PAs are produced and accumulate in leaves and in seeds and skin of the berries (Souquet et al. 1996). PA accumulation is initiated before flowering but is most active at flowering and at fruit setting (Downey et al. 2003; Bogs et al. 2005). In contrast, the synthesis and accumulation of PAs in leaves occur throughout leaf expansion (Bogs et al. 2005). Nonetheless, PA synthesis and accumulation can be increased when the plant is under abiotic or biotic stress (Peters and Constabel 2002; Dixon et al. 2005). In this context, the expression of both VvMATE genes is consistent with that expected for PA transporters. VvMATE1 transcripts were detected at high levels in reproductive tissues, specifically in seeds during early berry development. The expression profile of VvMATE1 is in agreement with that shown for TT12 in Arabidopsis, which is expressed specifically in ovules and developing seeds (Fig. 5a; Debeaujon et al. 2001). The transcriptional profile of VvMATE1 shows a good correlation with the expression of the transcription factors VvMYBPA1 and VvMYBPA2, which have been shown to induce VvMATE1 expression (Bogs et al. 2007; Terrier et al. 2009). In spite of that, analysis of its promoter did not show the presence of a MYBCORE cis-element which can be recognized by PA MYB transcription factors (MYBPAs) (Akagi et al. 2009). One possibility is that VvMATE1 might be induced by a transcription factor target of MYBPAs. On the other hand, the higher expression of VvMATE1 in seeds, similar to TT12, might be supported by the presence of the DPBF element (E-box) that has been shown to be mainly embryo specific (Kim et al. 1997). In contrast, VvMATE2 mRNA accumulates primarily in seeds and leaves (Fig. 5b) and the MYBCORE elements in its promoter can account for its expression in seeds. Similarly, its transcription profile in leaves resembles that of VvMYBPA2 in the same tissue (Terrier et al. 2009; Fig. 6). The over-representation of MYB-binding elements in the promoter of VvMATE2 compared to that of VvMATE1 might be an important determinant of the differential transcriptional regulation among MATE genes.
Comparative analysis of the promoters of VvMATE1, VvMATE2 and TT12 suggests a possible differential hormonal regulation mediated by ABA and MeJA. The involvement of ABA in the biosynthesis and accumulation of PAs has been described in grapevine and other crops. A seasonal ABA signal and a bZIP transcription factor, DkbZIP5, which act as an ABRE-binding protein, are responsible for controlling PA levels in Diospyros kaki (Akagi et al. 2012). Also, exogenous ABA treatment seems to affect PA biosynthesis in the earlier developmental stages of grape fruit skin, with expression changes in at least two Myb transcription factors, VvMYB5 and VvMYBPA1 (Lacampagne et al. 2010). On the other hand, the MeJA signal has been associated with defense responses to pathogen and wounding, where PAs play an important role, especially in the defense against herbivores (Harborne and Grayer 1993; Peters and Constabel 2002). Therefore, a MeJA-responsive motif in the promoter of VvMATE2, which is also expressed in leaves, was not unexpected. However, additional examination is needed to determine the functionality of these cis-elements in the VvMATE genes.
Regarding the two models proposed to PAs transporter and accumulation in plants, results presented in this article suggest that both models coexist in grapevine. The two putative PAs transporters, VvMATE1 and VvMATE2, appear to be involved in the transport and distribution of PAs in the different compartments of the seed cells in grape fruits. In this context, our results are in agreement rather with the classic model than with the recently proposed model which does not consider specific membrane transporters for accumulation of PAs in the vacuoles or for PAs distribution to other cellular compartments as nucleus or apoplast.
Materials and methods
Plant material
Grapevine (Vitis vinifera L. var. Carménère) plants grown in field conditions in a commercial vineyard in the Maule Valley (Central Chile) during the 2009–2010 growing season were used in this study. Random sampling of different organs was performed including roots and leaves and starting at early flowering until mature fruit stage (from October to April). Stages to be sampled were defined according to the modified Eichhorn-Lorenz System (Coombe 1995): flowers (EL 23), setting (EL 29–31), pre-veraison (EL 33), veraison (EL 35), post-veraison (EL 36) and mature (EL 38). The skin samples were only readily separated from the rest of the berry starting at the pre-veraison stage. Leaf samples were collected from fully expanded mature leaves. All collected samples were frozen in liquid nitrogen and stored at −80 °C until their use for RNA isolation. Sampling was carried out consistently at approximately 10:00 a.m. to discard possible oscillations in transcript levels.
RNA isolation and cDNA synthesis
RNA isolation was performed using 1–2 g of frozen tissue using the perchlorate method (5 M sodium perchlorate; 300 mM Tris–HCl pH 8; 1 % v/v SDS; 2 % v/v PEG 20,000; 8.5 p/v PVPP; 3 % v/v 2-mercaptoethanol) modified according to Davies and Robinson (1996). Three independent RNA extractions from each tissue were performed. RNA integrity was corroborated by formaldehyde agarose gel electrophoresis and their purity by OD260/280 ratio >1.95. Following DNase (DNAse I, Ambion) treatment of total RNA, first-strand cDNA synthesis was carried out from 2 μg of total RNA for each sample using oligo (dT) according to the manufacturer’s instructions (AffinityScript QPCR cDNA Synthesis Kit, Stratagene, La Jolla, CA).
Phylogenetic analysis
The full-length amino acid sequences of VvMATE1, VvMATE2 and other plant MATE were used as templates to perform a multi-alignment using the BioEdit Sequence Alignment Editor v7.0 software (Hall 1999). The potential transmembrane domains were predicted by the TMHMM program (Krogh et al. 2001). The phylogenetic tree was built from the resulting aligned sequences using the MEGA software (version 4; http://www.megasoftware.net) (Tamura et al. 2007) by the neighbor-joining method with bootstrap analysis of 1,000 replicates. The accession numbers were Vitis vinifera MATE1 (XP_002282907), MATE2 (XP_002282932), AM1 (ACN91542), AM3 (ACN88706); Arabidopsis thaliana AtTT12 (NP_191462), AtFRD3 (NP_187461); Brassica rapa BrTT12 (ACJ36213), Malus domestica MATE1 (ADO22709) and MATE2 (ADO22711); Medicago truncatula MATE1 (ACX37118) and MATE2 (HM856605); Oryza sativa TT12 (ABA99853); Nicotiana tabacum MATE1 (BAF47751), MATE2 (BAF47752) and JAT1 (CAQ51477); Populus tricoparca XP_002307572; Hordeum vulgare HvAACT1(BAF75822); Sorghum bicolor SbMATE (ABS89149).
Genetic constructions and transient expression assay in Arabidopsis protoplasts
The coding sequence of the target genes was amplified from cDNA of berries (cv. Carmenere) using Platinum® Taq DNA Polymerase High Fidelity (Invitrogen) and the following pair of primers: M1GFPfw: 5′-ACTCGAGATGGCCTCCGCAGCAGAG-3′ and M1GFPrv: 5′-ATCCATGGCTATCTTGTCTAGTACTAAATCCAAC-3′ for VvMATE1, and M2GFPfw: 5′-ACTCGAGATGGGTTCAGAAGAGTATCA-3′ and M2GFPrv: 5′-ATCCATGGCTACATCTTCCAATAAGTTTAAATTCTC-3′ for VvMATE2. Forward and reverse primers included XhoI and NcoI restriction sites, respectively, and the reverse primers had the stop codon deleted. The resulting DNA fragments were cloned into pGEM-T vector (Promega, Madison, WI, USA) and then sequenced. The pAM1 vector used for transient transformation was derived from pGreen0029 and pA7-GFP (Katrin Czempinski, Potsdam University, Germany) vectors, carrying an enhanced version of CaMV 35S promoter. The target genes were inserted between the XhoI and NcoI sites of the pAM1 vector upstream of the GFP gene, which creating VvMATE:GFP fusions. The final constructions were also confirmed by sequencing. The following organelle markers for vacuole, ER and Golgi were used respectively: V-rk, ER-rk and G-rk (Nelson et al. 2007). All organelle markers contained the mCherry reporter gene. The original constructs were reduced in size by cloning them into the pAM1 vector (Ryngajllo et al. 2011) using the following restriction sites: HindIII and XmaI for the vacuole and ER marker constructs, and HindIII and XbaI for the Golgi marker construct. These constructs were used in co-transformation experiments along with VvMATE1 and VvMATE2. Protoplasts obtained from 5-week-old Arabidopsis thaliana Col-0 plants by the tape-Arabidopsis sandwich method (Wu et al. 2009) were transformed with the different plasmids. The transformation was done as described in Wu et al. 2009 with prolonged incubation time of 15 min in PEG solution. The transformed Arabidopsis protoplasts were visualized 24 h after transformation using a confocal laser scanning microscope (TCS SP2/UV, Leica, Germany). The instrument was equipped with Argon and He/Ne lasers, beam splitter 488/543/633 and the following objectives: PL APO 20×, 63× W. Two different filter settings were used: (1) for GFP fluorescence, excitation with 488 nm, beam splitter DD 488/568 (double dichroic, reflects at 488 and 568 nm) and barrier filter BP 530 (band pass, 515–545 nm); (2) for mCherry, excitation at 568 nm, beam splitter DD 488/568 and barrier filter BP 590 (long pass > 590 nm). Chlorophyll autofluorescence was detected at 683–747 nm. During image acquisition, each line was scanned twice and averaged. Image analyses were performed with the Leica Confocal Software of TCS SP2 (version 2.61. build 1,537).
Gene expression analyses
Gene transcript levels were measured by quantitative PCR (qPCR) using a Stratagene Mx3000P (Agilent Technologies) system. All reactions were performed using the Brilliant SYBR Green Master Mix (Stratagene) according to the manufacturer. For each sample, qPCRs were carried out in triplicate (technical repeats) using 10 μl Master Mix, 0.5 μl of 250 nM primers, 1 μl of diluted cDNA and nuclease-free water to a final volume of 20 μl. Fluorescence was measured at the end of each amplification cycle. Amplification was followed by a melting curve analysis with continuous fluorescence acquisition during the 55–95 °C melt. The raw data were manually analyzed and expression was normalized against VvGADPH (accession number CN938023), which presents constitutive expression in vegetative tissues and reproductive tissues during berry development (Almada et al. 2009; Gainza-Cortés et al. 2012). The primers used for qPCR analysis were MATE1-F, 5′-GGAGATTCAAATAGAAGATGCCCAAG-3′ and MATE1-R, 5′-AACTAACCAACTTCCACCAACTCG-3′; MATE2-F, 5′-TCTGCCTTTGAGTGAAGAATGTAA-3′ and MATE2-R, 5′-AAGCCACAACAATACTAGAAGAAC-3′; GAPDH-Fwd, 5′-TTCCGTGTTCCTACTGTTG-3′ and GAPDH-Rev, 5′-CCTCTGAC- TCCTCCTT GAT-3′.
In silico analysis of promoters
Analysis of cis-regulatory elements in the VvMATE1, VvMATE2 and AtTT12 promoter regions was performed using the PLACE (http://www.dna.affrc.go.jp/PLACE/signalscan.html; Higo et al. 1999), PlantCARE (http://www.bioinformatics.psb.ugent.be/webtools/plantcare/html; Rombauts et al. 1999) and PlantPAN (http://www.plantpan.mbc.nctu.edu.tw/seq_analysis.php; Chang et al. 2008) databases.
Abbreviations
- PA:
-
Proanthocyanidin
- PAs:
-
Proanthocyanidins
- ABA:
-
Abscisic acid
- MeJA:
-
Methyl jasmonate
- GFP :
-
Green fluorescent protein
- qRT-PCR:
-
Quantitative reverse-transcriptase polymerase chain reaction
- ER:
-
Endoplasmic reticulum
- MATE :
-
Multidrug And Toxic Compound Extrusion
References
Abrahams S, Tanner GJ, Larkin PJ, Ashton AR (2002) Identification and biochemical characterization of mutants in the proanthocyanidin pathway in Arabidopsis. Plant Physiol 130:561–576
Abrahams S, Lee E, Walker A, Tanner G, Larkin P, Ashton A (2003) The Arabidopsis TDS4 gene encodes leucoanthocyanidin dioxygenase (LDOX) and is essential for proanthocyanidin synthesis and vacuole development. Plant J 35:624–636
Akagi T, Ikegami A, Tsujimoto T, Kobayashi S, Sato A, Kono A, Yonemori K (2009) DkMyb4 is a Myb transcription factor involved in proanthocyanidin biosynthesis in persimmon fruit. Plant Physiol 151:2028–2045
Akagi T, Katayama-Ikegami A, Kobayashi S, Sato A, Kono A, Yonemori K (2012) Seasonal abscisic acid signal and a basic leucine zipper transcription factor, DkbZIP5, regulate proanthocyanidin biosynthesis in persimmon fruit. Plant Physiol 158:1089–1102
Almada R, Cabrera N, Casaretto JA, Ruiz-Lara S, González E (2009) VvCO and VvCOL1, two CONSTANS homologous genes, are regulated during flower induction and dormancy in grapevine buds. Plant Cell Rep 28:1193–1203
Baxter I, Young J, Armstrong G, Foster N, Bogenschutz N, Cordova T, Peer W, Hazen S, Murphy A, Harper JF (2005) A plasma membrane H1-ATPase is required for the formation of proanthocyanidins in the seed coat endothelium of Arabidopsis thaliana. Proc Natl Acad Sci 102:2649–2654
Bogs J, Downey M, Harvey J, Ashton A, Tanner G, Robinson S (2005) Proanthocyanidin synthesis and expression of genes encoding leucoanthocyanidin reductase and anthocyanidin reductase in developing grape berries and grapevine leaves. Plant Physiol 139:652–663
Bogs J, Jaffé F, Takos A, Walker A, Robinson S (2007) The grapevine transcription factor VvMYBPA1 regulates proanthocyanidin synthesis durint fruit development. Plant Physiol 243:1347–1361
Boss P, Davies C, Robinson S (1996) Analysis of the expression of anthocyanin pathway genes in developing Vitis vinifera L. cv shiraz grape berries and the implications for pathway regdation. Plant Physiol 111:1059–1066
Braidot E, Zancani M, Petrussa E, Peresson C, Bertolini A, Patui S, Macri F, Vianello A (2008) Transport and accumulation of flavonoids in grapevine (Vitis vinifera L.). Plant Signal Behav 3:626–632
Brillouet JM, Romieu C, Schoefs B, Solymosi K, Cheynier V, Fulcrand H, Verdeil JL, Conéjéro G (2013) The tannosome is an organelle forming condensed tannins in the chlorophyllous organs of Tracheophyta. Ann Bot 112:1003–1014
Burko Y, Geva Y, Refael-Cohen A, Shleizer-Burko S, Shani E, Berger Y, Halon E, Chuck G, Moshelion M, Ori N (2011) From organelle to organ: ZRIZI MATE-Type transporter is an organelle transporter that enhances organ initiation. Plant Cell Physiol 52:518–527
Chang W-C, Lee T-Y, Huang H-D, Huang H-Y, Pan R-L (2008) PlantPAN: plant promoter analysis navigator, for identifying combinatorial cis-regulatory elements with distance constraint in plant gene groups. BMC Genom 9:561
Conn S, Curtin C, Bézier A, Franco C, Zhang W (2008) Purification, molecular cloning, and characterization of glutathione S-transferases (GSTs) from pigmented Vitis vinifera L. cell suspension cultures as putative anthocyanin transport proteins. J Exp Bot 59:3621–3634
Conn S, Franco C, Zhang W (2010) Characterization of anthocyanic vacuolar inclusions in Vitis vinifera L. cell suspension cultures. Planta 231:1343–1360
Coombe BG (1995) Growth stages of the grapevine: adoption of a system for identifying grapevine growth stages. Aust J Grape Wine Res 1:104–110
Cutanda-Perez MC, Ageorges A, Gomez C, Vialet S, Terrier N, Romieu C, Torregrosa L (2009) Ectopic expression of VlmybA1 in grapevine activates a narrow set of genes involved in anthocyanin synthesis and transport. Plant Mol Biol 69:633–648
Davies C, Robinson S (1996) Sugar accumulation in grape berries. Plant Physiol 111:275–283
Debeaujon I, Peeters A, León-Kloosterziel K, Koornneef M (2001) The TRANSPARENT TESTA12 gene of Arabidopsis encodes a multidrug secondary transporter-like protein required for flavonoid secuestration in vacuoles of the seed coat endothelium. Plant Cell 13:853–871
Debeaujon I, Nesi N, Pérez P, Devic M, Grandjean O, Caboche M, Lepiniec L (2003) Proanthocyanidin-accumulating cells in Arabidopsis testa: regulation of differentiation and role in seed development. Plant Cell 15:2514–2531
Dixon R, Xie DY, Sharma S (2005) Proanthocyanidins—a final frontier in flavonoid research? New Phytol 165:9–28
Downey M, Harvey J, Robinson S (2003) Analysis of tannins in seeds and skins of Shiraz grapes throughout berry development. Aust J Grape Wine Res 9:15–27
Durrett TP, Gassmann W, Rogers EE (2007) The FRD3-mediated efflux of citrate into the root vasculature is necessary for efficient iron translocation. Plant Physiol 144:197–205
Escribano-Bailon T, Alvarez-Garcia M, Rivas-Gonzalo JC, Herediam FJ, Santos-Buelga C (2001) Color and stability of pigments derived from the acetaldehyde-mediated condensation between malvidin 3-O-glucoside and (+)-catechin. J Agric Food Chem 49:1213–1217
Francisco RM, Regalado A, Ageorges A, Burla BJ, Bassin B, Eisenach C, Zarrouk O, Vialet S, Marlin T, Chaves MM, Martinoia E, Nagy R (2013) ABCC1, an ATP binding cassette protein from grape berry, transports anthocyanidin 3-O-glucosides. Plant Cell 25:1840–1854
Frank S, Keck M, Sagasser M, Niehaus K, Weisshaar B, Stracke R (2011) Two differentially expressed MATE factor genes from apple complement the Arabidopsis transparent testa12 mutant. Plant Biol 13:42–50
Furukawa J, Yamaji N, Wang H, Mitani N, Murata Y, Sato K, Katsuhara M, Takeda K, Ma JF (2007) An aluminum-activated citrate transporter in barley. Plant Cell Physiol 48:1081–1091
Gainza-Cortés F, Pérez-Díaz R, Pérez-Castro R, Tapia J, Casaretto JA, González S, Peña-Cortés H, Ruiz-Lara S, González E (2012) Characterization of a putative grapevine Zn transporter, VvZIP3, suggests its involvement in early reproductive development in Vitis vinifera L. BMC Plant Biol 12:111–124
Gawel R, Iland P, Francis I (2001) Characterizing the astringency of red wine: a case of study. Food Qual Prefer 12:83–94
Gény L, Saucier C, Bracco S, Daviaud F, Glories Y (2003) Composition and cellular localization of tannins in grape seeds during maturation. J Agric Food Chem 51:8051–8054
Gomez C, Terrier N, Torregrosa L, Vialet S, Fournier-Level A, Verrie C, Souquet JM, Mazauric JP, Klein M, Cheynier V, Ageorges A (2009) Grapevine MATE-Type proteins act as vacuolar H+-dependent acylated anthocyanin transporters. Plant Physiol 150:402–415
Gomez C, Conejero G, Torregrosa L, Cheynier V, Terrier N, Ageorges A (2011) In vivo grapevine anthocyanin transport involves vesicle-mediated trafficking and the contribution of anthoMATE transporters and GST. Plant J 67:960–970
Goodman CD, Casati P, Walbot V (2004) A multidrug resistanceassociated protein involved in anthocyanin transport in Zea mays. Plant Cell 16:1812–1826
Grotewold E, Davies K (2008) Trafficking and sequestration of anthocyanins. Nat Prod Comm 3:1251–1258
Hall TA (1999) BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser 1999(41):95–98
Harborne JB, Grayer RJ (1993) Flavonoids and insects. In: Harborne JB (ed) The flavonoids: advances in research since 1986. Chapman & Hall, London, pp 589–618
Higo K, Ugawa Y, Iwamoto M, Korenaga T (1999) Plant cis-acting regulatory DNA elements (PLACE) database. Nucleic Acids Res 27:297–300
Horton P, Park KJ, Obayashi T, Fujita N, Harada H, Adams-Collier CJ, Nakai K (2007) WoLF PSORT: protein localization predictor. Nucleic Acids Res 35:585–587
Kennedy J, Jones G (2001) Analysis of proanthocyanidin cleavage products following acid-catalysis in the presence of excess phloroglucinol. J Agric Food Chem 40:1740–1746
Kennedy J, Hayasaka Y, Vidal S, Waters E, Jones G (2001) Composition of grape skin proanthocyanidins at differents stages of berry development. J Agric Food Chem 49:5348–5355
Kim SY, Chung HJ, Thomas TL (1997) Isolation of a novel class of bZIP transcription factors that interact with ABA-responsive and embryo-specification elements in the Dc3 promoter using a modified yeast one-hybrid system. Plant J 11:1237–1251
Kitamura S (2006) Transport of flavonoids: from cytosolic synthesis to vacuolar accumulation. In: Grotewold E (ed) Science of flavonoids. Springer, Berlin, pp 123–146
Kitamura S, Shikazono N, Tanaka A (2004) TRANSPARENT TESTA 19 is involved in the accumulation of both anthocyanins and proanthocyanidins in Arabidopsis. Plant J 37:104–114
Kitamura S, Matsuda F, Tohge T, Yonekura-Sakakibara K, Yamazaki M, Saito K, Narumi I (2010) Metabolic profiling and cytological analysis of proanthocyanidins in immature seeds of Arabidopsis thaliana flavonoid accumulation mutants. Plant J 62:549–559
Krogh A, Larsson B, von Heijne G, Sonnhammer ELL (2001) Predicting transmembrane protein topology with a hidden markov model: application to complete genomes. J Mol Biol 305:567–580
Lacampagne S, Gagné S, Gény L (2010) Involvement of abscisic acid in controlling the proanthocyanidin biosynthesis pathway in grape skin: new elements regarding the regulation of tannin composition and leucoanthocyanidin reductase (LAR) and anthocyanidin reductase (ANR) activities and expression. J Plant Growth Regul 29:81–90
Lepiniec L, Debeaujon I, Routaboul JM, Baudry A, Pourcel L, Nesi N, Caboche M (2006) Genetics and biochemistry of seed flavonoids. Annu Rev Plant Biol 57:405–430
Liu J, Magalhaes JV, Shaff J, Kochian LV (2009) Aluminum-activated citrate and malate transporters from the MATE and ALMT families function independently to confer Arabidopsis aluminum tolerance. Plant J 57:389–399
Magalhaes JV, Liu J, Guimarães CT, Lana UG, Alves VM, Yi-H Wang, Schaffert RE, Hoekenga OA, Piñeros MA, Shaff JE, Klein PE, Carneiro NP, Coelho CM, Trick HN, Kochian LV (2007) A gene in the multidrug and toxic compound extrusion (MATE) family confers aluminum tolerance in sorghum. Nat Genet 39:1156–1161
Mané C, Souquet JM, Ollé D, Verries C, Verán F, Mazerolles G, Cheynier V, Fulcrand H (2007) Optimization of simultaneous flavanol, phenolic acid, and anthocyanin extraction from grapes using an experimental design: application to the characterization of champagne grape varieties. J Agric Food Chem 55:7224–7233
Marinova K, Pourcel L, Weder B, Schwarz M, Barron D, Routaboul JM, Debeaujon I, Klein M (2007) The Arabidopsis MATE transporter TT12 acts as a vacuolar flavonoid/H+-antiporter active in proanthocyanidin-accumulating cells of the seed coat. Plant Cell 19:2023–2038
Maron LG, Piñeros MA, Guimarães CT, Magalhaes JV, Pleiman JK, Mao C, Shaff J, Belicuas SNJ, Kochian LV (2010) Two functionally distinct members of the MATE (multidrug and toxic compound extrusion) family of transporters potentially underlie two major aluminum tolerance QTLs in maize. Plant J 61:728–740
Mathews H, Clendennen SK, Caldwell CG, Liu XL, Connors K, Matheis N, Schuster DK, Menasco DJ, Wagoner W, Lightener J, Wagner DR (2003) Activation tagging in tomato identifies a transcriptional regulator of anthocyanin biosynthesis, modification, and transport. Plant Cell 15:1689–1703
Müller M, Knudsen S (1993) The nitrogen reponse of a barley C-hordein promoter is controlled by positive and negative regulation of thr GCN4 and endosperm box. Plant J 4:343–355
Nawrath C, Heck S, Parinthawong N, Métraux J (2002) EDS5, an essential component of salicylic acid-dependent signaling for disease resistance in Arabidopsis, is a member of the MATE transporter family. Plant Cell 14:275–286
Nelson B, Cai X, Nebenfu A (2007) A multicolored set of in vivo organelle markers for co-localization studies in Arabidopsis and other plants. Plant J 51:1126–1136
Peleg H, Gacon K, Schlich P, Noble A (1999) Bitterness and astringency of flavan-3-ol monomers, dimers and trimers. J Sci Food Agric 79:1123–1129
Peters DJ, Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis: cloning and expression of dihydroflavonol reductase from trembling aspen (Populus tremuloides). Plant J 32:701–712
Pourcel L, Routaboul JM, Kerhoas L, Caboche M, Lepiniec L, Debeaujon I (2005) TRANSPARENT TESTA10 encodes a laccase-like enzyme involved in oxidative polymerization of flavonoids in Arabidopsis seed coat. Plant Cell 17:2966–2980
Poustka F, Irani NG, Feller A, Lu Y, Pourcel L, Frame K, Grotewold E (2007) A trafficking pathway for anthocyanins overlaps with the endoplasmic reticulum-to-vacuole protein-sorting route in Arabidopsis and contributes to the formation of vacuolar inclusions. Plant Physiol 145:1323–1335
Remy S, Fulcrand H, Labarbe B, Cheynier V, Moutounet M (2000) First confirmation in red wine of products resulting from direct anthocyanin-tannin reactions. J Sci Food Agric 80:745–751
Rombauts S, Déhais P, Van Montagu M, Rouzé P (1999) PlantCARE, a plant cis-acting regulatory element database. Nucleic Acids Res 27:295–296
Ryngajllo M, Childs L, Lohse M, Giorgi F, Lude A, Selbig J, Usadel B (2011) SLocX: predicting subcellular localization of Arabidopsis proteins leveraging gene expression data. Front Plant Sci 2:1–19
Saslowsky DE, Warek U, Winkel BSJ (2005) Nuclear localization of flavonoid enzymes in Arabidopsis. J Biol Chem 280:23735–23740
Seo PJ, Park J, Park MJ, Kim YS, Kim SG, Jung JH, Park CM (2012) A Golgi-localized MATE transporter mediates iron homoeostasis under osmotic stress in Arabidopsis. Biochem J 442:551–561
Shoji T, Inai K, Yazaki Y, Sato Y, Takase H, Shitan N, Yazaki K, Goto Y, Toyooka K, Matsuoka K, Hashimoto T (2009) Multidrug and toxic compound extrusion-type transporters implicated in vacuolar sequestration of nicotine in tobacco roots. Plant Physiol 149:708–718
Souquet JM, Cheynier V, Brossaud F, Moutounet M (1996) Polymeric proanthocyanidins from grape skins. Phytochemistry 43:509–512
Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol Biol Evol 24:1596–1599
Terrier N, Torregrosa L, Ageorges A, Vialet S, Verries C, Cheynier V, Romieu C (2009) Ectopic expression of VvMybPA2 promotes proanthocyanidin biosynthesis in grapevine and suggests additional targets in the pathway. Plant Physiol 149:1028–1041
Uhde-Stone C, Liu J, Zinn KE, Allan DL, Vance CP (2005) Transgenic proteoid roots of white lupin: a vehicle for characterizing and silencing root genes involved in adaptation to P stress. Plant J 44:840–853
Verries C, Guiraud JL, Souquet JM, Vialet S, Terrier N, Ollé D (2008) Validation of an extraction method on whole pericarp of grape berry (Vitis vinifera L. cv. Shiraz) to study biochemical and molecular aspects of flavan-3-ol synthesis during berry development. J Agric Food Chem 56:5896–5904
Wang H, Race EJ, Shrikhande AJ (2003) Anthocyanin transformation in Cabernet Sauvignon wine during aging. J Agric Food Chem 51:7989–7994
Wang H, Wang W, Li H, Zhang P, Zhan J, Huang W (2010) Gene transcript accumulation, tissue and subcellular localization of anthocyanidin synthase (ANS) in developing grape berries. Plant Sci 179:103–113
Winkel BSJ (2004) Metabolic channeling in plants. Annu Rev Plant Biol 55:85–107
Wu CY, Washida H, Onodera Y, Harada K, Takaiwa F (2000) Quantitative nature of the prolamin-box, ACGT and AACA motifs in a rice glutelin gene promoter: minimal cis-element requirements for endosperm-specific gene expression. Plant J 23:415–421
Wu FH, Shen SC, Lee LY, Lee SH, Chan MT, Lin CS (2009) Tape-Arabidopsis Sandwich—a simpler Arabidopsis protoplast isolation method. Plant Methods 5:16
Xie DY, Sharma S, Paiva N, Ferreira D, Dixon R (2003) Role of anthocyanidin reductase, encoded by BANYULS in plant flavonoid biosynthesis. Science 299:396–399
Yokosho K, Yamaji N, Ueno D, Mitani N, Ma JF (2009) OsFRDL1 is a citrate transporter required for efficient translocation of iron in rice. Plant Physiol 149:297–305
Zhao J, Dixon RA (2009) MATE transporters facilitate vacuolar uptake of epicatechin 3′-O-glucoside for proanthocyanidin biosynthesis in Medicago truncatula and Arabidopsis. Plant Cell 21:2323–2340
Zhao J, Pang Y, Dixon RA (2010) The mysteries of proanthocyanidin transport and polymerization. Plant Physiol 153:437–443
Zhao J, Huhman D, ShadleG He X-Z, Sumner LW, Tang Y, Dixon RA (2011) MATE2 mediates vacuolar sequestration of flavonoid glycosides and glycoside malonates in Medicago truncatula. Plant Cell 23:1536–1555
Acknowledgments
This work was supported by Fondo de Fomento al Desarrollo Científico y Tecnológico (FONDEF) grant No. G07I-1003. RPD was recipient of CONICYT doctoral fellowships and for a research stay. JPD was recipient of a Universidad de Talca doctoral fellowship and a DAAD fellowship for a research stay.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by P. Puigdomenech.
Electronic supplementary material
Below is the link to the electronic supplementary material.
299_2014_1604_MOESM1_ESM.ppt
Supplementary Fig. 1. Subcellular localization of fusion proteins and organellar markers. (A-D) VvMATE1-GFP without orgarnellar marker. (E–H) VvMATE2-GFP without organellar marker. (I-L) VvMATE2-GFP and vacuole marker (mCherry). (M-P) vacuolar marker (mCherry). (Q-T) Golgi marker (mCherry). (U-W) pAM1 empty vector (GFP). Bars in all images are 15 μm (PPT 1939 kb)
Rights and permissions
About this article
Cite this article
Pérez-Díaz, R., Ryngajllo, M., Pérez-Díaz, J. et al. VvMATE1 and VvMATE2 encode putative proanthocyanidin transporters expressed during berry development in Vitis vinifera L.. Plant Cell Rep 33, 1147–1159 (2014). https://doi.org/10.1007/s00299-014-1604-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00299-014-1604-9