
Abstract
The present study aimed at developing temporary immersion bioreactor techniques for multiplication of cacao somatic embryos. Temporary Immersion System (TIS), i.e. flooding of plant tissue at regular time intervals provides an efficient way to propagate plants. Somatic embryos were regenerated in twin flask bioreactors. The TIS proved to be suitable for mass regeneration of somatic embryos and for their subsequent direct sowing. The number of embryos after 3 months of culture was significantly higher in TIS cultures than in the solid medium variant. TIS also improved embryo development regarding the conversion to torpedo shaped forms. Matured embryos derived from TIS and pre-treated with 6% sucrose were converted into plants after direct sowing. Additionally to the influence of culture conditions on the development of somatic embryogenesis the content and composition of free amino acids were analysed. The content of free amino acids in somatic embryos rose as immersion frequency increased. The endogenous free GABA content in embryogenic callus was significantly higher than in non-embryogenic callus.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Conventional breeding methods used in T. cacao have resulted in a very narrow genetic base for commercial genotypes worldwide. The use of this narrow genetic base, confounded by mislabelling of breeding stock and germplasm collections, and the disruption in the continuity of breeding programmes has led to only marginal success in traditional cacao breeding (Lockwood 2003; Motilal et al. 2002). Typically, it takes from 3 to 6 years to produce a sexually mature cacao tree suitable for use in a breeding programme. Due also to the long breeding cycle, relatively little successful improvement of cacao has been made since the 1930s (Saunders et al. 2004). The lack of a long-term recurrent selection programme for cacao has resulted in the selection of heterozygous productive seedlings as parents. These seedlings are cloned and placed in seed gardens to serve as parental stock for multiplication of the clone. As a result of these factors, up to 80% of multiplied genotypes are unproductive on farms (Hunter 1990) due to genetic segregation.
The propagation of cacao can be achieved by rooted cuttings of buddings from plagiotropic or orthotropic shoots. Plagiotropic material, when propagated, results in a sprawling, bush-like architecture lacking the normal dimorphic growth habit. The resulting tree requires pruning to achieve a more convenient shape for harvesting and other farm operations. Plagiotropic material is inconsistent in forming taproots. Thus plagiotropic derived plants are sensible to wind. Orthotropic materials develop the seedlings-architecture but they are limited in availability and are poor in rooting and/or establishment (for review see Miller and Guiltinan 2003).
The improvement of T. cacao using modern breeding techniques can be greatly facilitated by multiplying elite material using somatic embryogenesis. It is now possible to produce somatic embryos and plantlets from a large number of genotypes (Maximova et al. 2002, 2005). However, production is not sufficient for a scaling-up step, and many genotypes remain recalcitrant. Moreover its application is limited by the long period of time from culture initiation to embryo production (6–8 months), and the relatively high cost per plant produced (Traore et al. 2003).
Since the main factor in the production cost of in vitro propagated plants is the high input of manual labour, reduction of labour by automation is one of the prime aims of commercial tissue culture. Generally, liquid culture systems are better amenable to automation and moreover result in faster growth and propagation, but these systems are often of limited use due to ensuing physiological abnormalities of the plants, especially hyperhydricity (Preil 2005).
Temporary Immersion Systems (TIS), i.e. flooding of plant tissue at regular time intervals, open possibilities to automate some in vitro culture stages (Alvard et al. 1993) and to increase the biological efficiency and productivity of propagated material (Lorenzo et al. 1998; Escalona et al. 1999; Martre et al. 2001). Plant morphology and physiological performance of crops in TIS are very similar to plants in ex vitro conditions providing a higher surviving rate (Teisson et al. 1996; Etienne and Berthouly 2002; Ziv 2005). Advantages of TIS have been reported on several tropical plant species (for review see González 2005). Mass propagation of somatic embryos from elite planting material has been demonstrated for Citrus deliciosa (Cabasson et al. 1997), Coffea arabica and Coffea canephora (Berthouly et al. 1995), Hevea brasiliensis (Etienne et al. 1997), Psidium guajava (Kosky et al. 2005). In the present report, the applicability of TIS for cacao somatic embryogenesis regeneration was studied using a twin flask bioreactor system.
Cell competence is associated with a particular metabolic and bioenergetic cell-state, which enables the somatic cell under auxin treatment to switch from a differentiated and resting state to a way which triggers the embryogenic pathway (Pedroso and Pais 1994,1995; Verdeil et al. 2007). During carrot somatic embryogenesis, development is associated with a continuous increase of glutamate and glutamine concentration whereas arginine, ornithine and γ-aminobutyric acid (GABA) have a biphasic behaviour, increasing in globular and torpedo stage embryos and decreasing in germinating embryos (Joy et al. 1996). Certain amino acids have been used as in vitro additives. In a comparative study of androgenesis-recalcitrant and androgenesis-responsive barley genotypes, Ouedraogo et al. (1998) were able to induce embryogenesis in the recalcitrant cultivar through the application of 19 amino acids. In this study, in addition to TIS investigations, we monitored amino acid distribution in callus material and embryos for subsequent possible use for improving cacao somatic embryogenesis. A comparison was made between embryos derived from solid medium with those originated from TIS. Such information is relevant to control the morphogenesis of plants and the effectiveness of plant production in a temporary immersion system.
Materials and methods
Primary somatic embryogenesis from cacao flower explants
The cacao clone Scavina6 (Sca6) was used for all experiments. The somatic embryogenesis process involved three stages as described by Li et al. (1998). In stages 1 and 2 corresponding to production of callus in the dark, flower buds were surface-sterilized in 3% sodium hypochlorite and used as explants. Staminode and petal explants were cultured for 2 weeks in a Petri dish containing 15 ml primary callus growth medium (PCG), then transferred for another 2 weeks in a secondary callus growth medium (SCG). PCG medium consisted of DKW basal salt (Driver and Kuniyuki 1984) supplemented with 250 mg l−1 glutamine, 200 mg l−1 myo-inositol, 20 g l−1 glucose, 2 mg l−1 2,4-D and 5 μg l−1 TDZ. SCG medium is made of Woody Plant Medium (WPM) and vitamins, 20 g l−1 glucose, 2 mg l−1 2,4-D and 50 μg l−1 BAP. Stage 3 corresponds to induction and maintenance of somatic embryos. Callus from SCG medium were subcultured every 2 weeks in embryo development medium (ED) which contained DKW salts and vitamins, 30 g l−1 saccharose and 1 g l−1 glucose. During the ED culture period, large number of somatic embryos developed. Cultures were maintained on ED with a subculture interval of 14 days. DKW vitamins consisted of 0.1 g l−1 myo-inositol, 2 mg l−1 thiamine–HCl, 1 mg l−1 nicotinic acid and 2 mg l−1 glycine.
Secondary embryogenesis from cotyledon explants of primary somatic embryos
Secondary somatic embryogenesis was induced as described by Maximova et al. (2002) from the cotyledons of primary somatic embryos. Cotyledons from mature embryos were excised and sub-cultured for 2 weeks in SCG medium for callus induction. Thereafter they were transferred in ED medium (sub-cultures every 2 weeks) where secondary somatic embryos regenerated.
Comparison of different culture systems
In order to compare the multiplication and growth of somatic embryos on solid medium and in a temporary immersion system, two different experiments were carried out. For treatment in solid medium, five cotyledon explants from primary somatic embryos were cultured on 15 ml SCG (14 days) and subsequently transferred on 15 ml ED medium for 3 months with subculture every 14 days. Media were solidified with 0.5% agar (Duchefa).
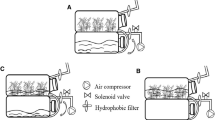
The temporary immersion system (TIS) used in our studies (Fig. 1a) was based on the “twin flasks” type from Escalona et al. (1999) and Hempfling and Preil (2005) with some modifications. The plant compartment consisted of glass jars (Weck) of 1,000 ml volume covered with glass lids and sealed with rubber rings. Inside this compartment cotyledon explants lay on top of a layer of 150 μm sieves (Laborbedarf-Vertriebs GmbH, Heidelberg, Germany) adapted between a 250 ml Kautex polypropylene bottle (Rotilabo®). To avoid floating, the weight of the Kautex was increased with stainless steel screws. The medium compartment was a Schott Duran® flask of 1000 ml volume. The lids of Weck flasks were 10 mm diameter holes while those of Duran flask were 8 mm diameter. The two compartments were connected with silicon tubing’s (8 mm inner, 12 mm outer diameter) coupled with a special polycarbonate 3/8″ system (Infiltec GmbH, No. MPC17006T03; Novodirect, No L39957, Germany). The medium was moved periodically by applying compressed air, which was sterilized through an autoclavable 0.2 μm air filter (Midisart 2000, Sartorius). Magnetic valves provide an on/off operation while digital timers controlled the frequency and length of the immersion period.

Somatic embryos regeneration on TIS after 3 months of culture. a Twin-flask temporary immersion system. b Morphological aspects of somatic embryos after 3 months in polypropylene bottles. c and d Somatic embryos from TIS (4 × 1) and TIS (6 × 1) showing their heterogeneity. e A TIS showing the domination of non-embryogenic callus over embryogenic callus
For somatic embryo multiplication in TIS, 400–500 mg of embryogenic callus were placed in the bioreactor with 300 ml ED medium. Media were changed every 30 days and complete embryo development was achieved after 3 months. All cultures were kept at 28 ± 1°C in the dark. Experiments to determine the optimal number of daily immersion cycles were carried out. In the first experiment the explants were flooded once a day for 1 min, “TIS (1 × 1)”. In treatment “TIS (4 × 1)”, explants were immersed four times per day for 1 min and in “TIS (6 × 1)”, they were flooded six times per day for 1 min.
Effect of immersion cycles and sugar content on germination
Three months after somatic embryo development, germination was triggered by applying DKW (mineral and vitamins) liquid medium containing 300 mg l−1 KNO3, amino acids as described by Li et al. (1998) and two different ratios of glucose/sucrose (20/10 and 10/5 g l−1). Immersion time consisted of four (TIS 4 × 1) and eight (TIS 8 × 1) daily immersions 1 min each. Experiments were carried out under 15 h photoperiod at 25 ± 1°C.
Sowing of somatic embryos and plantlet conversion
Two-month old germinated embryos with either small or large size cotyledons and plantlets were selected from TIS (4 × 1) or TIS (8 × 1) rinsed in sterile distilled water and sowed directly in Weck glasses containing one of the following substrates: (1) sand, (2) commercial potting mixture (CPM) (FloradurTM), (3) CPM overlaid with sand, (4) vermiculite or (5) a mixture of vermiculite and sand (1/1). Substrates were autoclaved, under moist conditions, twice (120°C for 20 min) with an interval of 24 h. About 20–30 ml of sterile modified Primary Embryo Conversion medium (PEC containing 10 g l−1 glucose and 5 g l−1 sucrose) were added to each vessel. Basal PEC medium consisted of DKW salts and vitamins, 300 mg l−1 KNO3, 20 g l−1 glucose, 10 g l−1 saccharose, 43.55 mg l−1 arginine, 18.76 mg l−1 glycine, 32.80 mg l−1 leucine, 45.65 mg l−1 lysine and 52.05 mg l−1 tryptophane.
Somatic embryos were carefully inserted into the medium using sterile forceps in the laminar flow hood. Ten embryos were placed in each vessel. Vessels were sealed with parafilm and incubated with a 15 h photoperiod (Philips TLD 58W/84o fluorescent tubes at 150–200 μmol s−1 m−2) at 25°C. There were 10 Weck vessels containing ten embryos for each treatment. Observation of the number of plants regenerated (regeneration defined as an embryo developing into a plant with a well-defined shoot with one or more true leaves) were recorded after 3 months.
To prepare somatic embryos for the best direct sowing, a preliminary test was done by exposing embryos with small size cotyledons to a high concentration (60 g l−1) of sucrose in ED medium for 2 weeks. For this test, embryos were inoculated in sand, vermiculite or a mixture of sand and vermiculite. Three Weck glasses containing ten pre-treated embryos each were prepared for each substrate.
HPLC analysis of free amino acids
Free amino acid contents were analysed according to the method described by Rohsius et al. (2006): 50–100 mg lyophilised callus or somatic embryos were powdered and stirred at 4°C for 1 h with 100–200 mg polyvinyl-polypyrrolidon (PVPP) and 5–10 ml distilled water. Immediately after adding water, the pH was adjusted at 2.5 with 10% aqueous trifluoroacetic acid. The homogenate was centrifuged for 10 min at 5,000 rpm. The clear supernatant solution was filtered through a 0.45 μm filter (Multoclear, CS-Chromatography). About 30 μl of each sample were lyophilised (1 h at −20°C, 0.05 mbar) directly into the vial and kept at −20°C until analysis.
Free amino acids were derivatized with O-phthalaldehyde (OPA) prior to HPLC analysis. Chromatographic separation was performed with a LiChroCART 250-4 (Merck) provided with precolumn LiChrospher 100 RP-18 (5 μm) (Merck, VWR international). Chromatographic analyses were made with a reverse phase binary gradient [A: 1.6 L sodium acetate solution/glacial acetic acid (50 mmol l−1; pH 6.2), 50 ml MeOH (Lichrosolv®, gradient grade), 20 ml tetrahydrofuran (Lichrosolv®; gradient grade); B: 200 ml sodium acetate solution/glacial acetic acid (50 mmol l−1; pH 6.2), 800 ml MeOH (Lichrosolv®, gradient grade)] at a flow rate of 1.3 ml min−1.
The OPA derivatization procedure was performed according to Rohsius et al. (2006). 20 μl of the derivative samples were injected into the column for separation and were subsequently detected with Hitachi F-1050 Fluorescence Spectrophotometer (λex = 334 nm, λem = 425 nm). The column temperature was 30°C. The autosampler was from Merck-Hitachi AS-4000. The solution degasser consisted of the Degassex DG-4400 from Phenomenex. Quantification was calculated via peak area of chromatograms from standard mixtures containing 1–10 pmol μl−1 of each amino acid. Total free amino acid concentrations were obtained by summation of the individual amino acid concentration.
Biological parameters and statistical analyses
Investigations were conducted after 3 months of culture: biomass fresh weight, proportion of embryos in the biomass, number of embryos, and proportion of torpedo-shaped embryos. Statistical analyses were carried out using SPSS Version 7.5. The results were analyzed using one-way ANOVA, followed by Duncan’s multiple range test. A probability of P ≤ 0.05 was considered significant.
Results
Effect of immersion cycles on somatic embryo production
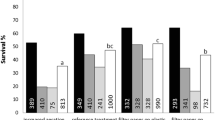
Regeneration of T. cacao secondary somatic embryos in an ED solid culture medium was compared to that produced in bioreactors. Over a period of 3 months, the biomass fresh weight, the number and the morphology of embryos differed significantly between the solid medium and TIS (Table 1). The tendency is that, increasing frequency immersion leads to an increase in the biomass fresh weight. System with four daily immersion of 1 min [TIS (4 × 1)] produced the largest amount of biomass mainly consisting of somatic embryos (159.0 ± 77.5 embryos per 250 ml bioreactor). The highest proportion (62.2 ± 15.1 and 48.4 ± 22.1 embryos) of torpedo-shapes embryos was obtained with TIS (4 × 1) and TIS (6 × 1).
Somatic embryos regenerated in TIS bioreactors were asynchronous displaying globular, torpedo and germinated embryos (Fig. 1b). In TIS (4 × 1) germinated embryos developed a pronounced hypocotyl (Fig. 1c). Embryos from TIS (6 × 1) germinated rarely and their cotyledon accumulated anthocyanins (Fig. 1d). In bioreactors, when a non-embryogenic callus occurred early, embryogenesis was strongly inhibited and callus growth dominated (Fig. 1e).
Influence of sugar content on somatic embryo production in bioreactors
The aim of this experiment was to test the effect of different sucrose/glucose concentrations in the culture medium on the multiplication rate of embryos with four daily immersions of 1 min each (TIS 4 × 1). After 3 months of culture, among the four sucrose/glucose treatments tested (Table 2), the concentration of 30/1 g l−1 resulted in the best production of fresh biomass and torpedo- shaped embryos. Reducing the sucrose/glucose to half strength (15/0.5 g l−1) yielded more embryos (185.0 ± 10.8) per 250 ml bioreactor. However most of these embryos remained at the globular stage. With 40/1 g l−1 sucrose/glucose, embryos developed large cotyledons which were tough and displaying stipule-like structures on their surface (pictures not shown).
Effect of immersion cycles and sugar content on germination of somatic embryos
Successful germination of cacao somatic embryos was obtained in the bioreactor where different morphological stages could be distinguished: torpedo-shaped, germinated stages of somatic embryos with small cotyledon size, germinated stages with large cotyledon size (rosette-like structure) and small plantlets with taproot and light-green leaves. Germinated embryos occurred in medium supplemented with 20/10 g l−1 glucose/sucrose during immersion cycle of 4 × 1 or 8 × 1. Plantlets were observed only in medium containing 10/5 g l−1 glucose/sucrose with an immersion cycle of 8 × 1. After 2 months of culture 26–28% of germinated embryos with small cotyledon size were recorded. Using the TIS (8 × 1) with low sugar concentration (glucose/sucrose, 10/5 g l−1), up to 70% of embryos were able to produce plantlets within one month of culture (data not shown).
Direct seeding of somatic embryos
All embryos and plantlets derived from germination medium started to dry out and shriveled. Those on CPM showed signs of contamination very early and died one month after subculture. In comparison the live period of embryos seeded on sand, CPM overlaid with sand, mixture of sand and vermiculite (Fig. 2) and on vermiculite (figure not shown), was longer (more than 2 months).

Direct sowing of cacao somatic embryos. a Mature somatic embryos directly sown on sand (a), on commercial potting mixture (CPM) + sand (b) and CPM (c). b 2 weeks old leave primordia developed from direct sown mature somatic embryos on sand. c One month old (a) and 2 months old (b and c) regenerated cacao plant from direct sowing
High sucrose concentration (60 g l−1) enhanced elongation of embryo axes. It also exerted strong positive effects on the conversion of embryos into plants and their subsequent growth. 10% of the embryos were able to develop true leaves after 2 weeks (Fig. 2b) and converted into plants after one month. Similar results were recorded on sand and on the mixture of sand and vermiculite (Fig. 2c). Plants regenerated from direct seeding of somatic embryos exhibited normal orthotropic shoot architecture (with flush emission) similar to that of the seminal seedlings.
Content and composition of free amino acids in different developmental stages
The concentration of free amino acids in cacao tissue depends on the ontogenic stage of the somatic embryos and the culture conditions. The main free amino acids detected were asparagine, glutamine, arginine and γ-aminobutyric acid (GABA) (Fig. 4).
Non embryogenic callus contained the highest concentration of total free amino acids in both solid medium and TIS (Fig. 3). The lowest concentrations of total free amino acids were found in globular embryos developed on solid medium. In embryogenic callus and somatic embryos, concentration of total free amino acids varied considerably depending on the culture support. Globular and cotyledonnary embryos displayed high amounts of total free amino acids in TIS (30,000 μg mg−1 DW on average) than in solid medium (20,000 μg mg−1 DW on average). In torpedo stage embryos the average content of free amino acid was not influenced by the culture conditions (Fig. 3).

Total free amino acid concentrations in callus and somatic embryos of T. cacao. Callus was cultured for 90 days either on ED solid medium, TIS (4 × 1), or TIS (6 × 1). NEC non-embryogenic callus, EC embryogenic callus. Means values (±SE) are indicated (n = 3). Embryogenic callus from TIS (6 × 1) was not analysed
Globular embryos from TIS (4 × 1) displayed high amounts of Glutamine, Arginine and GABA compared to torpedo and cotyledonnary embryos. In embryogenic callus, GABA was present in higher concentrations than in non embryogenic cells irrespective of the culture conditions (Fig. 4).

Amino acid concentrations in callus and somatic embryos of T. cacao. Callus was cultured for 90 days either on ED solid medium (a), TIS (4 × 1) (b), or TIS (6 × 1) (c). NEC non-embryogenic callus, EC embryogenic callus. Means values (±SE) are indicated (n = 3). Data ≤ 2,000 μg/g DW are less representative
Discussion and conclusion
The multiplication rate of cacao somatic embryos (SE) and conversion of SE to plantlets by a conventional in vitro technique (agar-gelled cultures) are relatively low. Under the described conditions only small numbers of embryos were produced. These results are similar to those of Maximova et al. (2002) using the same clone (Sca6). These authors produced 10.2 ± 1.9 embryos per explant after 12 months of culture on solid medium. This low production of secondary embryos hampers the commercial propagation of elite clones.
The system used in this study improved the efficiency of in vitro multiplication of cacao somatic embryos. One minute immersion every 6 h per day (TIS 4 × 1) increased the production of somatic embryos approximately 13-fold after 3 months of culture. This culture system also increased the conversion of somatic embryos into torpedo shaped forms. Teisson and Alvard (1995) also discussed that a higher frequency of short immersions stimulate embryo production.
Under the described TIS conditions, cacao somatic embryos promote multiplication without causing negative morphological characteristics, e.g. no hyperhydric embryos occurred. It is well established that prolonged contact between plant tissues and the liquid medium is a source of hyperhydricity (Ziv et al. 1983; Hussey 1986; Albarrán et al. 2005).Thus the good embryo morphology may be achieved by the sieve used (150 μm) which allowed a better drainage of explants after immersion.
Pre-treated mature embryos derived from TIS with 60 g l−1 sucrose were able to develop directly into plantlets after sowing on sand or on a mixture of sand and vermiculite. For Rosa multiflora L. cv. Montse it was also reported that the survival rate during acclimatisation was positively correlated with higher sucrose concentrations in the nutrient media (Capellades et al. 1991). Capellades et al. formulated the hypothesis that this is caused by the translocation of stored assimilates. Moreover, Winkelmann et al. (2006) found that storage proteins were accumulated in cyclamen somatic embryos grown in high sucrose concentrations. These authors concluded that the treatment with 60 g l−1 sucrose promoted maturation of cyclamen somatic embryos and improved the desiccation tolerance of the embryos (Winkelmann et al. 2004). Klimaszewska et al. (2004) were able to show that maturation in media with 6% sucrose as compared to 3% resulted in higher levels of the most important storage proteins, 11S globulins and 7S vicillin like proteins, in Pinus strobus.
Thus, pre-treatment according to our preliminary study could help avoid acclimatization of cacao somatic embryos since this stage is one of the critical in plant production via somatic embryogenesis. However, further improvement of the technique is necessary.
Mobilisation of nutrients in the culture medium is used by different biosynthesis pathways for embryo growth and development, e.g. amino acids, proteins, lipids. Differences in the concentrations of total amino acids were especially evident for globular somatic embryos grown on solid medium and in TIS. Temporary immersion culture increased the accumulation of free amino acids in early stages of somatic embryos while further developed stages revealed no differences compared to somatic embryos grown on solid medium. For zygotic embryos the increase of free amino acid content in the early developmental stages was also reported for Ocotea catharinensis (Santa-Catarina et al. 2006).
Especially for non embrygonic callus material the different culture conditions and systems influence the Arginine content. Our results are in contrast to Komamine et al. (1992); Joy et al. (1996); Thorpe and Stasolla (2001), who report higher concentrations of Arginine in embrygenic cultures of carrots.
The contribution of GABA to the total free amino acids in embryogenic callus is substantial compared to non embryogenic callus and further developmental stages of cacao somatic embryos. This finding fits the evidence that acquisition of embryogenic competence is a stress process. GABA is commonly found associated with stress conditions in plants (Bown and Shelp 1997; Mesnard et al. 2000). It is produced in plants as a result of the activity of glutamate decarboxylase and rapidly accumulates under various stress conditions (Shelp et al. 1999; Bouché et al. 2003; Bouché and Fromm 2004).
Further studies dealing with the content of free amino acids are needed to illuminate their role in the developmental processes of somatic embryogenesis.
The temporary immersion system used here makes cacao somatic embryo regeneration more efficient than the conventional method. We are presently using the system in the development of elite cacao clones. An investigation into different sucrose, PEG and abscisic acid concentrations and the duration of their application on the maturation of TIS-derived somatic embryos for direct seeding purposes is also underway.
References
Albarrán J, Bertrand B, Lartaud M, Etienne H (2005) Cycle characteristics in a temporary immersion bioreactor affect regeneration, morphology, water and mineral status of coffee (Coffea arabica) somatic embryos. Plant Cell Tissue Organ Cult 81:27–36
Alvard D, Côte F, Teisson C (1993) Comparison of methods of liquid medium culture for banana micropropagation. Effects of temporary immersion of explants. Plant Cell Tissue Organ Cult 32:55–60
Berthouly M, Dufour M, Alvard D, Carasco C, Alemanno L, Teisson C (1995) Coffee micropropagation in a liquid medium using the temporary immersion technique. In: ASIC Publishers (eds) Proceedings of the 16th international scientific Colloquim on Coffee, Kyoto, Japan. Vevey, Switzerland, pp 514–519
Bouché N, Fait A, Bouchez D, Moller SG, Fromm H (2003) Mitochondrial succinic-semialdehyde dehydrogenase of the γ-aminobutyrate shunt is required to restrict level of reactive oxygen intermediate in plants. Proc Natl Acad Sci USA 100:6843–6848
Bouché N, Fromm H (2004) GABA in plants: just a metabolite? Trends Plant Sci 9:110–115
Bown AW, Shelp BJ (1997) The metabolism and functions of γ-aminobutyric acid. Plant Physiol 115:1–5
Cabasson C, Alvard D, Dambier D, Ollitrault P, Teisson C (1997) Improvement of Citrus somatic embryos development by temporary immersion. Plant Cell Tissue Organ Cult 50:33–37
Capellades M, Lemeur R, Debergh P (1991) Effects of sucrose on starch accumulation and rate of photosynthesis in Rosa cultured in vitro. Plant Cell Tissue Organ Cult 25:21–26
Driver JA, Kuniyuki AH (1984) In vitro propagation of Paradox walnut root stock. Hortic Sci 19:507–509
Escalona M, Lorenzo JC, González B, Daquinta M, González JL, Desjardins Y, Borroto CG (1999) Pineapple (Ananas comosus L. Merr) micropropagation in temporary immersion systems. Plant Cell Rep 18:743–748
Etienne H, Berthouly M (2002) Temporary immersion systems in plant micropropagation. Plant Cell Tissue Organ Cult 69:215–231
Etienne H, Lartaud M, Michaux-Ferriere N, Carron MP, Berthouly M, Teisson C (1997) Improvement of somatic embryogenesis in Hevea brasiliensis (Müll. Arg) using the temporary immersion technique. In Vitro Cell Dev Biol Plant 33:81–87
González EJ (2005) Mass propagation of tropical crops in temporary immersion systems. In: Hvoslef-Eide AK, Preil W (eds) Liquid culture systems for in vitro plant propagation. Springer, Berlin, pp 197–211
Hempfling T, Preil W (2005) Application of a temporary immersion system in mass propagation of Phalaenopsis. In: Hvoslef-Eide AK, Preil W (eds) Liquid culture systems for in vitro plant propagation. Springer, Berlin, pp 231–242
Hunter RJ (1990) The status of cocoa (Theobroma cacao, Sterculiaceae) in the western hemisphere. Econ Bot 44:425–439
Hussey G (1986) Problems and prospects in the in vitro propagation of herbaceous plants. In: Withers LA, Alderson PG (eds) Plant tissue culture and its agricultural applications. Butterworths, Boston, pp 69–84
Joy RW IV, McIntyre DD, Vogel HJ, Thorpe TA (1996) Stage-specific nitrogen metabolism in developing carrot somatic embryos. Physiol Plant 97:149–159
Klimaszewska K, Morency F, Jones-Overton C, Cooke J (2004) Accumulation pattern and identification of seed storage proteins in zygotic embryos of Pinus strobus and in somatic embryos from different maturation treatments. Physiol Plant 121:682–690
Komamine A, Kawahara R, Matsumoto M, Sunabori S, Toya T, Fujiwara A, Tsukahara M, Smith J, Ito M, Fukuda H, Nomura K, Fujimura T (1992) Mechanisms of somatic embryogenesis in cell cultures: physiology, biochemistry, and molecular biology. In Vitro Cell Dev Biol Plant 28:11–14
Kosky RG, Perozo JV, Valero NA, Peñalver DA (2005) Somatic embryo germination of Psidium guajava L. in the RITA® temporary immersion system and on solid medium. In: Hvoslef-Eide AK, Preil W (eds) Liquid culture systems for in vitro plant propagation. Springer, The Netherlands, pp 225–229
Li Z, Traore A, Maximova S, Guiltinan M (1998) Somatic embryogenesis and plant regeneration from floral explants of cacao (Theobroma cacao L.) using thidiazuron. In Vitro Cell Dev Biol Plant 34:293–299
Lockwood R (2003) Who needs clothing? INGENIC Newsl 8:2–5
Lorenzo JC, Gonzalez BL, Escalona M, Teisson C, Espinosa P, Borroto C (1998) Sugarcane shoot formation in an improved temporary immersion system. Plant Cell Tissue Organ Cult 54:197–200
Martre P, Lacan D, Just D, Teisson C (2001) Physiological effects of temporary immersion on Hevea brasiliensis (Müll. Arg.) callus. Plant Cell Tissue Organ Cult 67:25–35
Maximova SN, Alemanno L, Young A, Ferriere N, Traore A, Guiltinan MJ (2002) Efficiency, genotypic variability, and cellular origin of primary and secondary somatic embryogenesis of Theobroma cacao L. In Vitro Cell Dev Biol Plant 38:252–259
Maximova SN, Young A, Pishak S, Miller C, Traore A, Guiltinan MJ (2005) Integrated system for propagation of Theobroma cacao L. In: Jain SM, Gupta PK (eds) Protocol for somatic embryogenesis in woody plants. Springer, The Netherlands, pp 209–227
Mesnard F, Azaroual N, Marty D, Fliniaux M-A, Robins RJ, Vermeersch G, Monti J-P (2000) Use of 15N reverse gradient two-dimensional nuclear magnetic resonance spectroscopy to follow metabolic activity in Nicotiana plumbaginifolia cell-suspension cultures. Planta 210:446–453
Miller CR, Guiltinan MJ (2003) Perspective on rapid vegetative multiplication for orthotropic scion and rootstock varieties of cocoa. International workshop on cocoa breeding for improved production systems (INGENIC), 19–21 October 2003, Accra, Ghana, pp 189–194
Motilal LA, Butler DR, Mooleedhar V (2002) Verification in global cocoa germplasm collections. INGENIC Newsl 7:4–8
Ouedraogo JT, St-Pierre CA, Collin J, Rioux S, Comeau A (1998) Effect of amino acids, growth regulators and genotype on androgenesis in barley. Plant Cell Tissue Organ Cult 53:59–66
Pedroso MC, Pais MS (1994) Early detection of embryogenic competence and of polarity in Cammelia japonica L. by electron probe X-ray microanalysis. Plant Sci 96:189–201
Pedroso MC, Pais MS (1995) Factors controlling somatic embryogenesis—Cell wall changes as an in vitro marker of embryogenic competence. Plant Cell Tissue Organ Cult 43:147–154
Preil W (2005) General introduction: a personal reflection on the use of liquid media for in vitro culture. In: Hvoslef-Eide AK, Preil W (eds) Liquid culture systems for in vitro plant propagation. Springer, Berlin, pp 1–18
Rohsius C, Matissek R, Lieberei R (2006) Free amino acid amounts in raw cocoas from different origins. Eur Food Res Technol 222:432–438
Santa-Catarina C, Silveira V, Balbuena TS, Viana AM, Estelita MEM, Handro W, Floh EIS (2006) IAA, ABA, polyamines and free amino acids associated with zygotic embryo development of Ocotea catharinensis. Plant Growth Regul 49:237–247
Saunders JA, Mischke S, Leamy EA Hameida AA (2004) Selection of international molecular standards for DNA fingerprinting of Theobroma cacao. Theor Appl Genet 110:41–47
Shelp BJ, Bown AW, MacLean MD (1999) Metabolism and function of γ-aminobutyric acid. Trends Plant Sci 4:446–452
Teisson C, Alvard D (1995) A new concept of plant in vitro cultivation liquid medium: temporary immersion. In: Terzi M et al (eds) Current issues in plant molecular and cellular biology. Kluwer Academic Publishers, Dordrecht, The Netherlands, pp 105–110
Teisson C, Alvard D, Berthouly B, Cote F, Escalant JV, Etienne H, Lartaud M (1996) Simple apparatus to perform plant tissue culture by temporary immersion. Acta Hortic 440:521–526
Thorpe TA, Stasolla C (2001) Somatic embryogenesis. In: Bhojwani SS, Soh WY (eds) Current trends in the embryology of angiosperms. Kluwer Academic Publishers, Dordrecht, The Netherlands, pp 279–236
Traore A, Maximova S, Guiltinan M (2003) Micropropagation of Theobroma cacao using somatic embryo-derived plants. In Vitro Cell Dev Biol Plant 39:332–337
Verdeil J-L, Alemanno L, Niemenak N, Trangarder TJ (2007) Pluripotent versus totipotent plant stem cells: dependence versus autonomy. Trends Plant Sci 12:245–252
Winkelmann T, Heintz D, Van Dorsselaer A, Serek M, Braun H-P (2006) Proteomic analyses of somatic and zygotic embryos of Cyclamen persicum Mill. reveal new insights into seed and germination physiology. Planta 224:508–519
Winkelmann T, Meyer L, Serek M (2004) Germination of encapsulated somatic embryos of Cyclamen persicum. Hortic Sci 39:1093–1097
Ziv M (2005) Simple bioreactors for mass propagation of plants. In: Hvoslef-Eide AK, Preil W (eds) Liquid culture systems for in vitro plant propagation. Springer, Berlin, pp 79–94
Ziv M, Meir G, Halevy A (1983) Factors influencing the production of hardened carnation plantlets in vitro. Plant Cell Tissue Organ Cult 2:55–60
Acknowledgment
The study was supported by the Alexander von Humboldt Stiftung (http://www.humboldt-stiftung.de) via grant to Nicolas Niemenak (Grant No. KAM/1115305). The authors express gratitude to Thomas Tumforde and Detlef Böhm for their assistance during experiments.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by R. Reski.
Rights and permissions
About this article
Cite this article
Niemenak, N., Saare-Surminski, K., Rohsius, C. et al. Regeneration of somatic embryos in Theobroma cacao L. in temporary immersion bioreactor and analyses of free amino acids in different tissues. Plant Cell Rep 27, 667–676 (2008). https://doi.org/10.1007/s00299-007-0497-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00299-007-0497-2